
Ферменты казеин связки структурные белки дифтерийный токсин
ВНИМАНИЕ! САЙТ ЛЕКЦИИ.ОРГ проводит недельный опрос. ПРИМИТЕ УЧАСТИЕ. ВСЕГО 1 МИНУТА.
По механизму действия белковые бактериальные токсины делят на следующие группы:
Токсины, повреждающие клеточные мембраны (мембранотоксины).
Токсины ингибиторы синтеза белка (цитотоксины).
Токсины, активирующие пути метаболизма, вторичных посредников (мессенджеров) - функциональные блокаторы.
Активаторы иммунного ответа.
Протеазы.
1.Токсины, повреждающие клеточные мембраны – мембранотоксины. Эти токсины способны повреждать плазматическую мембрану эукариотических клеток двумя путями:
-с помощью ферментативного гидролиза,
-в результате формирования пор в мембране.
Такие повреждения вызывают не только лизис клетки, но и способствуют распространению бактерий в макроорганизме.
А. С помощью ферментативного гидролиза. Ферментативный гидролиз – действие альфа-токсина Clostridiumperfringens, обладающего активностью фосфолипазы С. Б. В результате формирования пор в мембране. Вызванные ими повреждения – они формируют трансмембранные поры и нарушают селективный вход и выход ионов через плазматическую мембрану, что ведет к лизису клетки. В эту группу входят: гемолизин E . coli, лейкотоксин, О-листериолизин Lysteriamonocytogenes; пневмолизин S . pneumoniae.
2. Токсины ингибиторы синтеза белка. Они блокируют синтез белка на субклеточном уровне. Субстратом для них служат факторы элонгации и 28 S рибосомальная РНК. Они индуцируют развитие гемолитического уремического синдрома. Эти токсины проникая из просвета кишечника связываются с рецепторами эндотелиальных клеток капилляров почечных гломерул. Это приводит к набуханию клеток, сужение просвета сосудов, способствует агрегации тромбоцитов, развитию гемолиза эритроцитов и уремии.
В эту группу входят антиэлонгаторы - например, дифтерийный гистотоксин, токсин синегнойной палочки, они выводят из строя фермент трансферазу 2, ответственный за элонгацию (наращивание) полипептидной цепи на рибосомах, что приводит к блокаде синтеза белка и вызывает гибель клетки. К данному типу токсинов принадлежат токсины с энтеропатогенной активностью и дермонекротоксины.
3.Токсины, активирующие пути метаболизма, вторичных посредников (мессенджера). В молекулу токсина включен фермент аденилатциклаза, который генерирует вторичный мессенджер.
Вторичные посредники – это малые сигнальные молекулы, которые являются компонентами передачи сигнала в клетке. К ним относятся циклические нуклеотиды, ионы Са, оксид азот. Они быстро образуются, активируют эффекторные белки, которые опосредуют клеточные реакции на внеклеточные сигналы. То есть активируют пути метаболизма.
Они влияют на функцию отдельных белков эукариотической клетки, не вызывая ее гибели. Механизм их действия заключается в том, что они способны активировать вторичных посредников, которые усиливают клеточные реакции на внеклеточные сигналы.
К данной группе относятся: цитотоксический некротизирующий фактор, термолабильный LT и термостабильный ST токсины E . coli ;коклюшный и дерматонекротический токсиныBordetellapertussis; токсины А и ВClostridiumdifficile, холерный энтеротоксин. В эту группу входят нейротоксины (тетаноспазмин, ботулинический токсин).
4. Активаторы иммунного ответа. К токсинам, входящим в эту группу относятся: токсин синдрома токсического шока ( TSST -1), энтеротоксины и эксфолиативные токсиныStreptococcuspyogenes. Они относятся к суперантигенам и могут действовать непосредственно на презентирующие клетки иммунной системы.
5.Протеазы – ботулинический и столбнячный нейротоксины.
Мишенями действия токсинов в клетках является группа белков, которые участвуют в процессах высвобождения нейромедиаторов и ингибирования секреции ацетилхолина. Возникают нарушения мышечных сокращений, что приводит к развитию вялых параличей перифирических нервов. Столбнячный и ботулинический токсины – супертоксины – это самые сильные биологические яды.
Эндотоксины.
Эндотоксины относятся к бактериальным модулинам, обладающим большим спектром биологической активности, индуцирующим синтез цитокинов и других медиаторов. За проявление биологической активности эндотоксинов ответственна вся молекула ЛПС. В основе действия ЛПС лежит неспецифическое лиганд- липидное взаимодействие.
§ 10. КЛАССИФИКАЦИЯ БЕЛКОВ
Существуют несколько подходов к классификации белков: по форме белковой молекулы, по составу белка, по функциям. Рассмотрим их.
Классификация по форме белковых молекул
По форме белковых молекул различают фибриллярные белки и глобулярные белки.
Фибриллярные белки представляют собой длинные нитевидные молекулы, полипептидные цепи которых вытянуты вдоль одной оси и скреплены друг с другом поперечными сшивками (рис. 18,б). Эти белки отличаются высокой механической прочностью, нерастворимы в воде. Они выполняют главным образом структурные функции: входят в состав сухожилий и связок (коллаген, эластин), образуют волокна шелка и паутины (фиброин), волосы, ногти, перья (кератин).
В глобулярных белках одна или несколько полипептидных цепей свернуты в плотную компактную структуру – клубок (рис. 18,а). Эти белки, как правило, хорошо растворимы в воде. Их функции многообразны. Благодаря им осуществляются многие биологические процессы, о чем подробнее будет изложено ниже.

Рис. 18. Форма белковых молекул:
а – глобулярный белок, б – фибриллярный белок
Классификация по составу белковой молекулы
Белки по составу можно разделить на две группы: простые и сложные белки. Простые белки состоят только из аминокислотных остатков и не содержат других химических составляющих. Сложные белки, помимо полипептидных цепей, содержат другие химические компоненты.
К простым белкам относятся РНКаза и многие другие ферменты. Фибриллярные белки коллаген, кератин, эластин по своему составу являются простыми. Запасные белки растений, содержащиеся в семенах злаков, – глютелины, и гистоны – белки, формирующие структуру хроматина, принадлежат также к простым белкам.
Среди сложных белков различают металлопротеины, хромопротеины, фосфопротеины, гликопротеины, липопротеины и др. Рассмотрим эти группы белков подробнее.
Металлопротеины
К металлопротеинам относят белки, в составе которых имеются ионы металлов. В их молекулах встречаются такие металлы, как медь, железо, цинк, молибден, марганец и др. Некоторые ферменты по своей природе являются металлопротеинами.
Хромопротеины
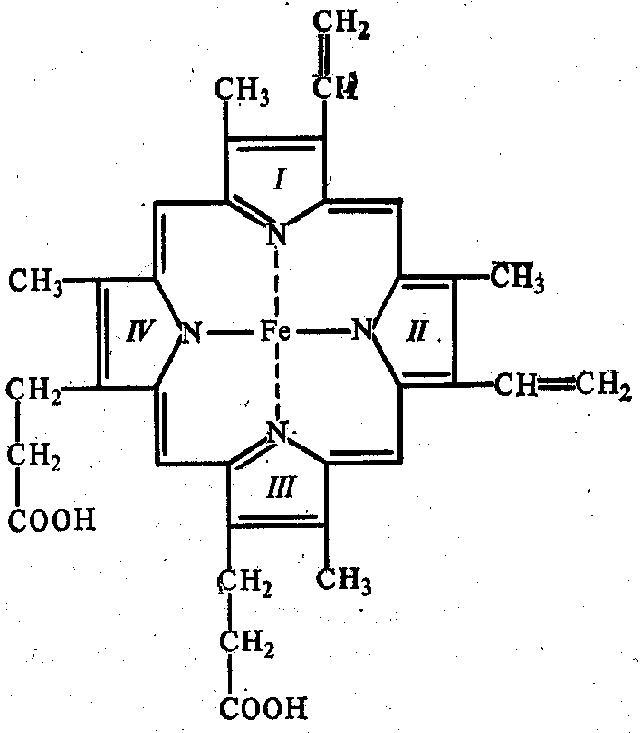
В составе хромопротеинов в качестве простетической группы присутствуют окрашенные соединения. Типичными хромопротеинами являются зрительный белок родопсин, принимающий участие в процессе восприятие света, и белок крови гемоглобин (Hb), четвертичная структура которого рассмотрена в предыдущем параграфе. В состав гемоглобина входит гем, представляющий собой плоскую молекулу, в центре которой расположен ион Fe 2+ (рис. 19). При взаимодействии гемоглобина с кислородом образуется оксигемоглобин. В альвеолах легких гемоглобин насыщается кислородом. В тканях, где содержание кислорода незначительно, оксигемоглобин распадается с выделением кислорода, который используется клетками:
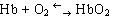
.
Гемоглобин может образовывать соединение с оксидом углерода (II), которое называется карбоксигемоглобином:
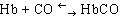
.
Карбоксигемоглобин не способен присоединять кислород. Вот почему происходит отравление угарным газом.
Гемоглобин и другие гем-содержащие белки (миоглобин, цитохромы) называют еще гемопротеинами из-за наличия в их составе гема (рис. 19).
Фосфопротеины
Фосфопротеины в своем составе содержат остатки фосфорной кислоты, связанные с гидроксильной группой аминокислотных остатков сложноэфирной связью (рис. 20).
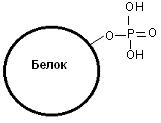
Рис. 20. Фосфопротеин
К фосфопротеинам относится белок молока казеин. В его состав входят не только остатки фосфорной кислоты, но и ионы кальция. Фосфор и кальций необходимы растущему организму в больших количествах, в частности, для формирования скелета. Кроме казеина, в клетках много и других фосфопротеинов. Фосфопротеины могут подвергаться дефосфорилированию, т.е. терять фосфатную группу:
фосфопротеин + Н2 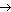
протеин + Н3РО4
Дефосфорилированные белки могут при определенных условиях быть снова фосфорилированы. От наличия фосфатной группы в их молекуле зависит их биологическая активность. Одни белки проявляют свою биологическую функцию в фосфорилированном виде, другие – в дефосфорилированном. Посредством фосфорилирования – дефосфорилирования регулируются многие биологические процессы.
Липопротеины
К липопротеинам относятся белки, содержащие ковалентно связанные липиды. Эти белки встречаются в составе клеточных мембран. Липидный (гидрофобный) компонент удерживает белок в мембране (рис. 21).
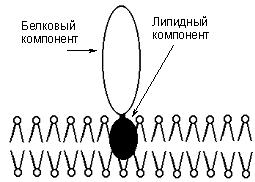
Рис. 21. Липопротеины в клеточной мембране
К липопротеинам относят также белки крови, участвующие в транспорте липидов и не образующие с ними ковалентную связь.
Гликопротеины
Гликопротеины содержат в качестве простетической группы ковалентно связанный углеводный компонент. Гликопротеины разделяют на истинные гликопротеины и протеогликаны. Углеводные группировки истинных гликопротеинов содержат обычно до 15 – 20 моносахаридных компонентов, у протеогликанов они построены из очень большого числа моносахаридных остатков (рис. 22).
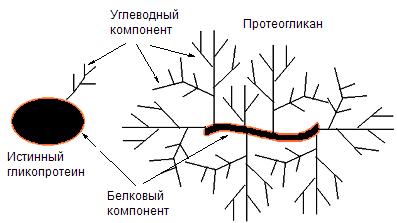
Рис. 22. Гликопротеины
Гликопротеины широко распространены в природе. Они встречаются в секретах (слюне и т.д.), в составе клеточных мембран, клеточных стенок, межклеточного вещества, соединительной ткани и т.д. Многие ферменты и транспортные белки являются гликопротеинами.
Классификация по функциям
По выполняемым функциям белки можно разделить на структурные, питательные и запасные белки, сократительные, транспортные, каталитические, защитные, рецепторные, регуляторные и др.
Структурные белки
К структурным белкам относятся коллаген, эластин, кератин, фиброин. Белки принимают участие в формировании клеточных мембран, в частности, могут образовывать в них каналы или выполнять другие функции ( рис. 23).
Рис. 23. Клеточная мембрана.
Питательные и запасные белки
Питательным белком является казеин, основная функция которого заключается в обеспечении растущего организма аминокислотами, фосфором и кальцием. К запасным белкам относятся яичный белок, белки семян растений. Эти белки потребляются во время развития зародышей. В организме человека и животных белки в запас не откладываются, они должны систематически поступать с пищей, в противном случае может развиться дистрофия.
Сократительные белки
Сократительные белки обеспечивают работу мышц, движение жгутиков и ресничек у простейших, изменение формы клеток, перемещение органелл внутри клетки. Такими белками являются миозин и актин. Эти белки присутствуют не только в мышечных клетках, их можно обнаружить в клетках практически любой ткани животных.
Транспортные белки
Гемоглобин, рассмотренный в начале параграфа, является классическим примером транспортного белка. В крови присутствуют и другие белки, обеспечивающие транспорт липидов, гормонов и иных веществ. В клеточных мембранах находятся белки, способные переносить через мембрану глюкозу, аминокислоты, ионы и некоторые другие вещества. На рис. 24 схематически показана работа переносчика глюкозы.
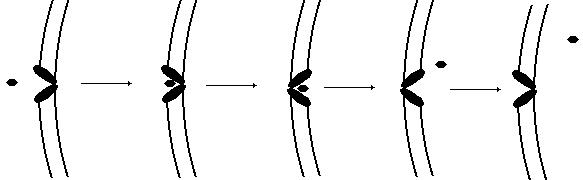
Рис. 24. Транспорт глюкозы через клеточную мембрану
Белки-ферменты
Каталитические белки, или ферменты, представляют собой самую многообразную группу белков. Почти все химические реакции, протекающие в организме, протекают при участии ферментов. К настоящему времени открыто несколько тысяч ферментов. Более подробно они будут рассмотрены в следующих параграфах.
Защитные белки
К этой группе относятся белки, защищающие организм от вторжения других организмов или предохраняющие его от повреждений. Иммуноглобулины, или антитела, способны распознавать проникшие в организм бактерии, вирусы или чужеродные белки, связываться с ними и способствовать их обезвреживанию.
Другие компоненты крови, тромбин и фибриноген, играют важную роль в процессе свертывания крови. Они предохраняют организм от потери крови при повреждении сосудов. Под действием тромбина от молекул фибриногена отщепляются фрагменты полипептидной цепи, в результате этого образуется фибрин:
фибриноген
фибрин.
Образовавшиеся молекулы фибрина агрегируют, формируя длинные нерастворимые цепи. Сгусток крови вначале является рыхлым, затем он стабилизируется за счет межцепочечных сшивок. Всего в процессе свертывания крови участвует около 20 белков. Нарушения в структуре их генов является причиной такого заболевания, как гемофилия – сниженная свертываемость крови.
Рецепторные белки
Клеточная мембрана является препятствием для многих молекул, в том числе и для молекул, предназначенных для передачи сигнала внутрь клеток. Тем не менее клетка способна получать сигналы извне благодаря наличию на ее поверхности специальных рецепторов, многие из которых являются белками. Сигнальная молекула, например, гормон, взаимодействуя с рецептором, образует гормон-рецепторный комплекс, сигнал от которого передается далее, как правило, на белковый посредник. Последний запускает серию химических реакций, результатом которых является биологический ответ клетки на воздействие внешнего сигнала (рис. 25).
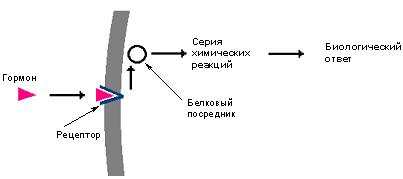
Рис.25. Передача внешних сигналов в клетку
Регуляторные белки
Белки, участвующие в управлении биологическими процессами, относят к регуляторным белкам. К ним принадлежат некоторые гормоны. Инсулин и глюкагон регулируют уровень глюкозы в крови. Гормон роста, определяющий размеры тела, и паратиреоидный гормон, регулирующий обмен фосфатов и ионов кальция, являются регуляторными белками. К этому классу белков принадлежат и другие протеины, участвующие в регуляции обмена веществ.
Интересно знать! В плазме некоторых антарктических рыб содержатся белки со свойствами антифриза, предохраняющие рыб от замерзания, а у ряда насекомых в местах прикрепления крыльев находится белок резилин, обладающий почти идеальной эластичностью. В одном из африканских растений синтезируется белок монеллин с очень сладким вкусом.
1. Фибриллярным белкам присущи следующие характеристики:
а) для них важна вторичная структура;
б) их длинные цепи скрепляются сшивками;
в) их цепи свернуты в глобулы;
г) для них важна третичная структура;
д) образуют длинные волокна;
е) являются регуляторными белками.
2. Для третичной структуры белков характерно:
а) в ее образовании принимают участие только водородные связи;
б) образуется при укладке полипептидных цепей в глобулы;
в) на ее уровне существуют белки, выполняющие защитную и механическую функции;
г) образуется при упорядоченном свертывании полипептидной цепи в спираль;
д) в ее образовании принимают участие водородные, ионные, дисульфидные связи;
е) на ее уровне существуют ферменты, антитела, некоторые гормоны.
Заполните пропуски
(заполните пропуски в предложениях, используя слова, выделенные курсивом)
В результате взаимодействия различных (1…) спирализованная молекула белка образует (2…) структуру, которая в свою очередь зависит от (3…) структуры, т.е. от (4…) аминокислот в молекуле белка. Субъединицы (отдельные цепи) некоторых белков образуют (5…) структуру. Примером такого белка является (6…).
(Гемоглобин, первичной, аминокислот, четвертичную, последовательности, третичную.)
Выберите верные утверждения
1. Белки по составу делятся на глобулярные, фибриллярные и промежуточные.
2. Мономером белка является радикал.
3. Аминокислота имеет радикал, аминогруппу, карбоксильную группу.
4. Вторичная структура белка представлена спиралью или складчатым слоем.
5. Все белки – ферменты.
6. Дисульфидные связи – связи между серосодержащими аминокислотами в молекуле белка.
7. Активный центр фермента – это липид.
8. Гистоны входят в состав хромосом.
9. В 1890 г. Э. Фишером был открыт активный центр фермента.
11. Фибриноген – белок промежуточной структуры.
12. Денатурация – это восстановление трехмерной структуры конформации белка.
13. Гемоглобин – белок четвертичной структуры.
14. Альбумины – это простые белки.
15. Простетическая группа гликопротеида – углевод.
16. Казеин молока – глобулин.
17. Аминокислота – амфотерное соединение.
18. Ферменты и антитела – глобулярные белки.
19. Связь между Н и О называют пептидной.
20. Реакция конденсации идет с выделением СО2.
21. Первичную конформацию белка поддерживают водородные связи.
22. Трипсин катализирует гидролиз белков.
23. Ферменты – это биологические катализаторы.
24. Водородные связи между витками спирали образуются через 2 аминокислоты.
25. Ферменты обладают специфичностью.
26. Активность фермента не зависит от рН.
27. Анаболические реакции – реакции распада.
28. Глюкагон инициирует распад гликогена в печени.
29. Миоглобин переносит О2 в мышцах.
30. Ферменты делят на 6 классов.
Установите соответствие
(установите соответствие между содержанием первого, второго и третьего столбца таблицы)
| Класс белков (по функциям) | Примеры | Локализация/функция |
| А. Ферменты | 1. Казеин | 9. Связки |
| Б. Структурные белки | 2. Дифтерийный токсин | 10. Свертываемость крови |
| В. Гормоны | 3. Тромбин | 11. Дифтерийная палочка |
| Г. Транспортные белки | 4. Гемоцианин | 12. Кровь беспозвоночных |
| Д. Защитные белки | 5. Инсулин | 13. Островки Лангерганса в поджелудочной железе |
| Е. Сократительные белки | 6. Трипсин | 14. Катализирует гидролиз белков |
| Ж. Запасные белки | 7. Эластин | 15. Неподвижные нити миофибрилл |
| З. Токсины | 8. Актин | 16. Белок молока |
Найдите ошибки в приведенном тексте, исправьте их
(укажите номера предложений, в которых сделаны ошибки, объясните их)
1.
1. Большое значение в строении и жизнедеятельности организмов имеют белки. 2. Это биополимеры, мономерами которых являются азотистые основания. 3. Белки входят в состав плазматической мембраны. 4. Многие белки выполняют в клетке ферментативную функцию. 5. В молекулах белка зашифрована наследственная информация о признаках организма. 6. Молекулы белка и тРНК входят в состав рибосом.
2.
1. Все присутствующие в организме белки – ферменты. 2. Каждый фермент ускоряет течение нескольких химических реакций. 3. Активный центр фермента соответствует конфигурации субстрата, с которым он взаимодействует. 4. Активность ферментов не зависит от таких факторов, как температура, рН среды и др.
Токсин дифтерийной палочки (Corynebacterium diphtheriae) является простым. Он синтезируется в виде одной полипептидной цепи с м.в. 62 кД. В результате активации происходит гидролиз пептидной связи в области дисульфидной петли белковой молекулы, в присутствии тиоловосстанавливающих агентов и белок протоксина фрагментируется на 2 полипептида: фрагмент А с м.в. 22-24 тыс. и фрагмент В - 38-39 тыс.
Фрагмент А - обладает ферментативными свойствами, является активатором (218 а.о.) и фрагмент В (371 а.о.) является рецептором.
Субьединица А обладает НАД - гликогидролазной активностью и АДФ - рибозилтрансферазной активностью.
Токсическое действие дифтерийного токсина осуществляется в результате последовательных процессов:
1) адсорбция на специфических клеточных рецепторах;
2) транспорт через плазматическую мембрану;
3) инактивация внутриклеточной мишени, участвующей в биосинтезе белка.
Клеточным рецептором, с которым взаимодействует дифтерийный токсин, является гликопротеин плазматической мембраны, м.в. 160 000. Этот гликопротеин обнаружен лишь у чувствительных к токсину клеток. Предполагается, что он обладает свойством протеазы.
Транспорт токсина через плазматическую мембрану может быть осуществлен 2 путями: активаторная субьединица проникает при помощи везикул, включающихся в эндоцитоз или проникновение активаторной субьединицы А идет с помощью специальных каналов в липидных двухслойных мембранах. Эти каналы образуются с помощью гидрофобного фрагмента В.
В основе токсического действия дифтерийного токсина лежит его способность подавлять биосинтез белка в чувствительных клетках.
Дифтерийный токсин специфически инактивирует фактор элонгации - 2, который соединяется с рибосомами и участвует в элонгации полипептидных цепей.
На первом этапе токсин гидролизует НАД до АДФ-рибозы и никотинамида, на втором - переносит АДФ - рибозу на EF-2. При этом образовавшийся комплекс полностью неактивен.
Под действием формалина в молекуле токсина образуется метиленовый мостик, который препятствует диссоциации токсина на фрагменты А и В. Особое значение имеют детерминанты, локализованные на С - терминальном участке фрагмента В. От них зависит присоединение токсина к клеточной мембране. Поэтому антитела только против этого участка полностью нейтрализуют токсин.
Другие токсины бактерий вызывающие блок синтеза белков.
Также как и у коринебактерий экзотоксин А Pseudomonas aeruginosa вызывает АДФ-рибозилирование фактора элонгации 2 (EF-2), нарушая синтез белка.
Shigella dysenteriae серотипа 1 и некоторые другие клинически значимые бактерии, например Stx-продуцирующие (Stx - Токсин Шига) E.coli (ЕНЕС [энтерогеморрагические E.coli]), вырабатывают Stx-токсины (или веротоксины). Stx-токсины инактивируют рибосомальную РНК, нарушая ее взаимодействие с факторами эллонгации. Подавление белкового синтеза данной группой токсинов приводит в конечном итоге к гибели клетки-мишени.
Stx-токсины являются мощными цитотоксинами и могут быть разделены на две группы, отличающиеся по антигенным свойствам и имеющие 50-60% гомологии: Stx=Stx-1 и Stx2-токсины. Stx-токсин S.dysenteriae серотипа 1 и Stx1-токсин E.coli отличаются только одной аминокислотой. Различные модификации Stx2-токсинов обнаружены у штаммов E.coli. Хотя Stx2-токсин рассматривается в качестве прототипа этой группы, были найдены различные модификации, отличающиеся по антигенным свойствам, степени сродства к рецепторам и способности активироваться под действием интестинальной слизи. Некоторые из перечисленных свойств обусловлены различиями в одном-двух нуклеотидах кодирующих генов.
Ген stx у S.dysenteriae всегда расположен на хромосоме, тогда как гены, кодирующие Stx1 и Stx2, могут входить в состав бактериальной хромосомы или генетического материала лизогенных бактериофагов. Гены, кодирующие А и В субъединицы Stx-токсинов (stxA и stxB соответственно), объединены в оперон. Оперон Stx/Stx1-токсинов (но не Stx2) содержит область, ответственную за регуляцию экспрессии Stx- и Stx1-токсинов, которая в свою очередь зависит от наличия ионов железа. На экспрессию Stx2-токсина не влияют ни наличие железа, ни другие внешние факторы. Однако кишечная слизь усиливает активность некоторых разновидностей Stx2-токсина. Stx-токсины, не способны активно выделяться через стенку бактериальной клетки. Предполагается, что в данном случае токсин попадает во внешнюю среду при лизисе клетки.
Stx-токсины имеют типичную АВ-структуру, то есть ферментативно активная субъединица А нековалентно связана с субъединицей В, непосредственно взаимодействующей с мишенью.
Субъединица А, обладающая ферментативной активностью, действует как N-гликозидаза, отщепляя единичный адениновый остаток от 28S рибосомальной РНК. Подобная депуринизация в конечном итоге подавляет синтез белка в пораженной клетке. При этом к действию N-гликозидазы одинаково чувствительны рибосомы про- и эукариот.
EHEC лишь недавно приобрела статус "проблемного" микроорганизма после вспышки в 1983 г. геморрагического колита, вызванного употреблением гамбургеров, подвергнутых недостаточной термической обработке. EHEC О157:Н7 ежегодно вызывает в США около 20 тыс. случаев геморрагического колита, тысячу случаев гемолитико-уремического синдрома и 100 летальных исходов.
Сайт СТУДОПЕДИЯ проводит ОПРОС! Прими участие :) - нам важно ваше мнение.
Бактериальные токсины
Токсические вещества, синтезируемые бактериями, по своей природе относятся к белкам и липополисахаридам. Липополисахариды (ЛПС) – эндотоксины, локализуются в клеточной стенке бактерий и освобождаются только после их разрушения. Токсические вещества белковой природы подразделяются на полностью секретируемые (экзотоксины), частично секретируемые и несекретируемые.
Способность бактерий образовывать белковые токсины называется токсигенностью.Установлено, что у одних видов бактерий – (Сorynebacterium diphtheriae, Staph. aureus) гены токсигенности (tox+ -ген) локализованы в ДНК умеренного фага, у других (E. соli, B. anthracis) – в плазмидах, у холерного вибриона – в хромосоме.
Одни из них являются термолабильными, другие относительно термостабильными. Так, например, термолабильный дифтерийный токсин, гистотоксин, разрушается при 60 0 С в течение часа, а столбнячный – в течение 20 мин. Термостабильные токсины клостридий ботулизма Cl. botulinum, кишечной палочки, стафилококков могут переносить кратковременное кипячение.
Итак, патогенные бактерии продуцируют загадочные субстанции, которые прямо или косвенно оказывают токсическое действие на клетки и организм хозяина. Экзотоксины – это секретируемые микробные протеины, обычно ферменты, которые убивают клетки хозяина в исключительно маленьких концентрациях.
Организация и механизм действия токсической молекулы. Большинство белковых токсинов представляют собой А-В структуру. Эта структура предполагает наличие двух компонентов – В-субъединицы, которая участвует в связывании токсина с рецептором на поверхности клетки хозяина и способствует транспортировке токсина в клетку хозяина; и А-субъединицы – проявляющей энзиматическую (токсическую) активность в клетке хозяина. Структура В-доменов зависит от структуры рецепторов-мишеней, с которыми взаимодействует токсин.
1. Порообразующие токсины. К ним относят бактериальные токсины, функционирующие посредством вставки в плазматическую мембрану хозяина и формирующие в ней трансмембранные поры, приводящие клетку к лизису. Механизм их действия хорошо изучен на примере альфа-токсина S.aureus, рассматриваемого как прототип пороформирующего цитотоксина. Стафилококковый альфа-токсин является цитолитическим в отношении различных типов клеток. К другим членам RTX-семейства относят гемолизин E.coli (HlyA), аденилатциклазу Bordetella pertussis (коклюшный), лейкотоксин Pasterella haemolitica.
Порообразующие токсины повышают проницаемость поверхностной мембраны эритроцитов (гемолизины) и лейкоцитов (лейкоцидины), вызывая гемолиз эритроцитов и разрушение лейкоцитов. Образование поры включает целый каскад вторичных реакций, приводящих к другим патологическим последствиям (активация эндонуклеаз, высвобождение цитокинов и медиаторов воспаления и др.).
2. Ингибирующие синтез белка. Субстратами для этих токсинов служат факторы элонгации и рибосомальная РНК. Дифтерийный токсин и экзотоксин A псевдомонад являются специфическими АДФ-рибозилтрансферазами, которые рибозилируют фактор элонгации 2 и, таким образом инактивируя его, подавляют синтез белка в клетках. Шигатоксин (Stx-токсин), так же называемый веротоксином, продуцируется Shigella dysenteriae первого серотипа и сравнительно недавно появившимися Stx-продуцирующими штаммами E. coli (STEC). Stx-токсины имеют типичную А-В структуру: энзиматически активную А-субъединицу, нековалентно связанную с 1-5 В-субъединицами. В результате действия этих токсинов происходят структурные изменения в рибосомальной РНК, синтез белка прекращается и клетка гибнет.
3. Генерирующие образование вторичных мессенджеров (посредников) – цитонекротические факторы(CNF1). Бактериальные токсины могут влиять на функцию отдельных белков эукариотической клетки, непосредственно не приводя ее к гибели. Для этого они активируют так называемых вторичных посредников, которые способны в большой степени усиливать и искажать клеточную реакцию на внеклеточные сигналы, нарушая клеточное деление. Цитотоксины – блокируют синтез белка на субклеточном уровне, нарушают клеточное деление. Например, дифтерийный гистотоксин, токсин синегнойной палочки выводят из строя фермент трансферазу 2, ответственную за наращивание полипептидной цепи на рибосоме. К этому типу принадлежат токсины с энтеропатогенной активностью и дермонекротоксины, поражающие соответствующие ткани и органы.
4. Протеолитические токсины (протеазы). Ботулинический и столбнячный токсины (оба цинк-металлоэндопротеазы), в опытах на животных обнаруживают наименьшую из известных LD50. Удивительно, насколько различную клиническую картину дают поражения этими токсинами, имеющими столь значительное сходство в структуре, энзиматической активности и мишенях среди клеток нервной системы, но при этом различающиеся путями проникновения в макроорганизм. Например, ботулинический токсин проникает в организм энтерально и вызывает вялые параличи периферических нервов. Столбнячный же токсин, образуясь на поверхностях ран, колонизированных Clostridium tetani, приводит к спастическим параличам через поражение ЦНС.
Летальный фактор B.anthracis также относится к разряду протеаз.
5. Активаторы иммунного ответа. Самое большое семейство токсинов данного типа называют токсинами-суперантигенами (PTSAg). Они могут действовать непосредственно на Т-клетки и антигенпрезентирующие клетки иммунной системы. Это приводит к массивной пролиферации более 20% периферических Т-клеток.
Следствием Т-клеточной экспансии является массивное высвобождение интерлейкинов (1, 2 и 6 типов), гамма-интерферона, факторов некроза опухолей (альфа и бета) и др. Совместно эти цитокины вызывают гипотензию, высокую температуру и диффузные эритематозные высыпания. Токсины данного типа характерны для случайных и факультативных паразитов.
Иммуногенные свойства белковых токсинов проявляются в способности вызывать иммунный ответ со стороны макроорганизма, в частности, индуцировать синтез специфических антител – антитоксинов, нейтрализующих гомологичный токсин.
Отличительной особенностью ряда белковых токсинов, например, столбнячного, дифтерийного, ботулинического, является их способность под действием 0,5 – 0,4% формалина и при температуре – 38–50 0 С утрачивать свою ядовитость, сохраняя при этом иммуногенные свойства. Такие токсины получили название анатоксинов. Они применяются в качестве вакцин для профилактики и лечения одноименных заболеваний.
Таким образом, свойства экзотоксинов сводятся к следующему:
Эндотоксины(только у Гр-)
Эндотоксины прочно связаны с телом бактериальной клетки. Свойство бактерий образовывать токсические вещества, вызывающие симптомы интоксикации, в том числе выделять в окружающую среду при их разрушении эндотоксины, называется токсичностью.В отличие от белковых токсинов, эндотоксины термостабильны и образуются Гр- бактериями, выделяясь в окружающую среду только после гибели бактериальной клетки. Это возбудители брюшного тифа, паратифов, гонореи, бруцеллеза, туляремии, менингита и многие др. патогенные Гр- бактерии. Это сложные белковолипополисахаридные комплексы, которые в лабораторных условиях можно получить путем экстракции трихлоруксусной кислотой.
В отличие от белковых токсинов, они
Симптомы интоксикации при заболеваниях, вызванных Гр- микробами, однотипны и связаны с действием образующихся медиаторов воспаления. ЛПС запускает синтез более 20 различных биологически активных веществ, которые обусловливают патогенез эндотоксикоза и обладают пирогенным действием. Основной точкой приложения являются макрофаги.
Эндотоксины менее токсичны, поражают организм в больших дозах, скрытый период у них исчисляется часами, избирательное действие выражено слабо. Они термоустойчивы, некоторые эндотоксины выдерживают кипячение и автоклавирование.
При введении больших доз вызывают стандартную реакцию: наблюдается угнетение фагоцитоза, слабость, одышка, расстройство кишечника, понижение температуры, падение сердечной деятельности. У людей поступление эндотоксина в кровяное русло приводит к лихорадке, лейкопении, гипогликемии, гипотонии. Большие дозы эндотоксина могут привести к токсико-септическому шоку.
Небольшие дозы эндотоксина, образующиеся постоянно представителями нормальной микрофлоры тела человека в кишечнике, оказывают благоприятное стимулирующее воздействие на клетки иммунной системы макроорганизма, что ведет к повышению неспецифической защиты макроорганизма, усилению его устойчивости к инфекционным заболеваниям и увеличению противоопухолевой активности клеток. В отличие от белковых токсинов, из эндотоксинов нельзя получить анатоксины.
Резюме о патогенности
Таким образом, патогенность носит сложный полидетерминантный характер. Основными материальными носителями патогенности микробов являются морфологические структуры клетки, ферменты и токсины. В макроорганизме они оказывают не изолированное, а комплексное воздействие. Например, нейраминидаза холерного вибриона способствует адгезии возбудителя к эпителиальным клеткам слизистой оболочки тонкого кишечника и взаимодействию его энтеротоксина с ганглиозидными рецепторами клеток, а гемоцитолизин, образуя каналы в мембранах клеток, ведет их к осмотическому повреждению и делает аденилатциклазу клеточных мембран более доступной.
Один и тот же фактор патогенности может участвовать в различных фазах инфекционного процесса, а в одной и той же фазе могут участвовать различные факторы патогенности. Например, капсулы бактерий способствуют их адгезии, препятствуют фагоцитозу и экранируют компоненты клетки, активирующие комплемент по альтернативному пути. Эндотоксин и инвазивные белки Гр- кишечных палочек не только способствуют их инвазии и развитию симптомов интоксикации, но и защищают бактерии от действия соляной кислоты и ферментов в желудке.
| | | следующая лекция ==> | |
| Основные особенности патогенных микроорганизмов | | | Инфекционного процесса |
Нам важно ваше мнение! Был ли полезен опубликованный материал? Да | Нет
Читайте также: