Анатомия и физиология вентрального кохлеарного ядра (ВКЯ)
Добавил пользователь Владимир З. Обновлено: 27.01.2026
слухова система є сенсорна система для сенсу слуху. Він включає як органи чуття (вуха) і слухові частини сенсорна система.
Зміст
Огляд системи
Лійки зовнішнього вуха звук вібрації до барабанної перетинки, збільшуючи звуковий тиск у середньому діапазоні частот. Кісточки середнього вуха ще більше посилюють вібраційний тиск приблизно в 20 разів. Основа стрічки поєднує вібрації в равлику через овальне вікно, яке вібрує перилімфну рідину (присутню у всьому внутрішньому вусі) і змушує кругле вікно розширюватися, коли овальне вікно випинається.
Вестибулярний і барабанний протоки заповнені перилімфою, а менший кохлеарний проток між ними заповнений ендолімфою, рідиною з дуже різною концентрацією іонів і напругою. [1] [2] [3] Вібрації перилімфи вестибулярного протоку згинають орган зовнішніх клітин Корті (4 лінії), викликаючи вивільнення престину в кінчиках клітин. Це призводить до того, що клітини хімічно видовжуються і стискаються (соматичний двигун), а пучки волосся зміщуються, що, в свою чергу, електрично впливає на рух базилярної мембрани (двигун пучків волосся). Ці двигуни (зовнішні клітини волосся) підсилюють амплітуди біжучої хвилі в 40 разів. [4] Зовнішні клітини волосся (OHC) мінімально інервуються спіральним ганглієм у повільних (немієлінізованих) взаємних комунікативних пучках (30+ волосків на нервове волокно); це протиставляє внутрішні клітини волосся (IHC), які мають лише аферентну іннервацію (30+ нервових волокон на одне волосся), але тісно пов’язані. Існує в три-чотири рази більше OHC, ніж IHC. Базилярна мембрана (ВМ) є бар'єром між лусочками, уздовж краю яких сидять IHC і OHC. Ширина базисної мембрани і жорсткість змінюються для контролю частот, які найкраще сприймаються IHC. На кохлеарній основі BM найвужчий і найжорсткіший (високочастотні), тоді як на верхівці кохлеарної області він найширший і найменш жорсткий (низькочастотні). Текторіальна мембрана (TM) сприяє посиленню кохлеарної ампліфікації, стимулюючи OHC (прямий) та IHC (за допомогою вібрацій ендолімфи). Ширина та жорсткість TM паралельні BM та подібним чином допомагають у диференціації частоти. [5] [6] [7] [8] [9] [10] [11] [12] [13]
вищий оливковий комплекс (SOC), в понах, є першим зближенням лівого та правого кохлеарних імпульсів. SOC має 14 описаних ядер; тут використано їх скорочення (див Покращений оливковий комплекс для їх повних назв). MSO визначає кут, з якого вийшов звук, вимірюючи різницю в часі в лівій та правій інформації. LSO нормалізує рівень звуку між вухами; він використовує інтенсивність звуку, щоб допомогти визначити кут звуку. LSO іннервує IHC. ВНТБ іннервує OHC. MNTB інгібує LSO через гліцин. LNTB є імунними до гліцину, використовуються для швидкої передачі сигналів. DPO є високочастотними та тонотопними. DLPO є низькочастотними та тонотопними. VLPO мають ту саму функцію, що і DPO, але діють в іншій галузі. PVO, CPO, RPO, VMPO, ALPO та SPON (інгібуються гліцином) є різними сигнальними та інгібуючими ядрами. [14] [15] [16] [17]
Трапецієподібне тіло - це місце, де більша частина волокон кохлеарного ядра (CN) розпадається (хрест зліва направо і навпаки); це перехрестя сприяє локалізації звуку. [18] CN розпадається на вентральний (VCN) і дорсальний (DCN) регіони. VCN має три ядра. [ потрібні роз’яснення ] Кущі клітини передають інформацію про хронометраж, їх форма в середньому викликає тремтіння часу. Зірчасті (подрібнювальні) клітини кодують звукові спектри (піки та долини) за допомогою просторових нейронних швидкостей випалу на основі слухової вхідної сили (а не частоти). Клітини восьминога мають майже найкращу часову точність під час стрільби, вони декодують слуховий часовий код. DCN має 2 ядра. DCN також отримує інформацію від VCN. Веретеноподібні клітини інтегрують інформацію для визначення спектральних сигналів до місця розташування (наприклад, чи походить звук спереду чи ззаду). Кохлеарні нервові волокна (30000+) мають найбільш чутливу частоту і реагують на широкий діапазон рівнів. [19] [20]
Спрощено, сигнали нервових волокон передаються кущистими клітинами до бінауральних областей в оливковий комплекс, в той час як піки та долини сигналу відзначаються зоряними клітинами, а час сигналу витягується клітинами восьминога. Бічний лемніск має три ядра: спинні ядра найкраще реагують на двосторонній вхід і мають налаштовані на складність відповіді; проміжні ядра мають широкі налаштування відповіді; а вентральні ядра мають широкі та помірно складні криві налаштування. Вентральні ядра бічного лемніскусу допомагають нижньому колікулу (IC) декодувати амплітудно-модульовані звуки, даючи як фазові, так і тонічні відповіді (короткі та довгі ноти відповідно). IC отримує вхідні дані, не показані, в тому числі візуальні (прекуктальна область: рухає очима до звуку. Вищий колікул: орієнтація та поведінка щодо предметів, а також зони рухів очей (саккада), понс (верхні мозочкові плодоніжки: таламус до мозочок зв'язок / почути звук і дізнатися поведінкову реакцію), спинний мозок (периаведуктальний сірий: чути звук і інстинктивно рухатися) і таламус. Вищевказане означає, що ІС впливає на «реакцію на здивування» та очні рефлекси. Окрім мультисенсорної інтеграції, ІС реагує на певні частоти амплітудної модуляції, дозволяючи виявляти висоту тону. IC також визначає різницю в часі бінаурального слуху. [21] Медіальне колінчасте ядро ділиться на вентральне (реле та інгібуючі реле клітини: частота, інтенсивність та бінауральна інформація топографічно передаються), дорсальне (широкі та складні налаштовані ядра: зв’язок із соматосенсорною інформацією) та медіальне (широке, складне та вузьке налаштоване ядра: інтенсивність реле та тривалість звуку). Слухова кора (AC) привносить звук в усвідомлення / сприйняття. AC визначає звуки (розпізнавання імен звуків), а також визначає місце походження звуку. AC - це топографічна карта частоти з пучками, що реагують на різні гармонії, терміни та висоту тону. Правий змінного струму більш чутливий до тональності, лівий змінного струму більш чутливий до хвилинних послідовних відмінностей у звуці. [22] [23] Ростромедіальні та вентролатеральні префронтальні кори беруть участь в активації під час тонального простору та зберіганні короткочасних спогадів відповідно. [24] Викрутка / поперечна скронева звивина Гешля включає область і функціональність Верніке, вона активно бере участь у процесах звучання емоцій, виразів обличчя емоцій та звукової пам'яті. Енторінальна кора є частиною «системи гіпокампу», яка допомагає та зберігає зорові та слухові спогади. [25] [26] Надмаргінальна звивина (SMG) допомагає в розумінні мови і відповідає за співчутливі реакції. SMG пов'язує звуки зі словами з кутовою звивиною та допомагає у виборі слів. SMG об’єднує тактильну, візуальну та слухову інформацію. [27] [28]
Вентральное кохлеарное ядро - Ventral cochlear nucleus
в вентральное кохлеарное ядро (VCN), слуховой нерв волокна попадают в мозг через нервный корешок в VCN. Вентральное кохлеарное ядро делится на переднее вентральное (передне-вентральное) кохлеарное ядро (AVCN) и заднее вентральное (задневентральное) кохлеарное ядро (PVCN). В VCN волокна слухового нерва разветвляются, восходящая ветвь иннервирует AVCN, а нисходящая ветвь иннервирует PVCN, а затем продолжается до дорсальное кохлеарное ядро. Упорядоченная иннервация волокнами слухового нерва дает AVCN тонотопный организация вдоль дорсовентральной оси. Волокна, которые несут информацию от верхушки улитки и настроены на низкие частоты, контактируют с нейронами вентральной части AVCN; те, которые несут информацию от основания улитки и настроены на высокие частоты, контактируют с нейронами в дорсальной части AVCN. AVCN заселяют несколько популяций нейронов. Кустистые клетки получают входной сигнал от волокон слухового нерва через особенно большие окончания, называемые концевые лампочки Held. Они связываются звездчатые клетки через более обычные бутоны.
Содержание
Типы клеток
Переднее ядро улитки содержит несколько типов клеток, которые достаточно хорошо соответствуют различным типам физиологических единиц. Кроме того, эти типы клеток обычно имеют определенные проекционные узоры.
Кустистые клетки
Названы из-за ветвящейся древовидной природы их дендритных полей, видимых с помощью Метод Гольджи они получают большие концевые лампочки Held от волокон слухового нерва. Кустообразные клетки бывают трех подтипов, которые проецируются на разные ядра-мишени в верхнем оливковом комплексе.
Шаровидный
Глобулярные кустистые клетки проецируют большие аксоны на контралатеральное медиальное ядро трапециевидное тело (MNTB), в высший оливарный комплекс где они синапсируют с основными клетками через одну чашечка Хельда и несколько меньших коллатералей синапсов ипсилатерально в заднем (PPO) и дорсолатеральном периоливном ядрах (DLPO), латеральной верхней оливе (LSO) и латеральном ядре трапециевидного тела (LNTB); контралатерально в дорсомедиальное периоливное ядро (DMPO), вентральное ядро трапециевидного тела (VNTB), ядро paragigantocellularis lateralis (PGL) и вентральное ядро латерального лемниска (VNLL). Аксоны всегда посылают коллатерали в MNTB, но не обязательно вызывают коллатерали, которые иннервируют каждое из других ядер. [1]
Большой сферический
Сферические густые клетки проецируются ипсилатерально к LSO, с двух сторон к медиальной верхней оливе (MSO) и LNTB и контралатерально к VNTB и VNLL. Наиболее важной целью этих прогнозов, по-видимому, является наделение MSO и LSO их межураторным временем и чувствительностью уровня (соответственно). [2]
Маленький сферический
Маленькие сферические густые клетки, вероятно, выступают на ипсилатеральную боковую верхнюю оливу. Они не проецируются ни на медиальные верхние оливки, ни на медиальное ядро трапециевидного тела.
Мультиполярные (звездчатые) клетки
Мультиполярные клетки делятся на две отдельные группы. Те, чьи аксоны выходят из aVCN через трапециевидное тело, Т-звездчатые клетки, имеют более длинные дендриты, чем кустистые клетки, которые обычно лежат на одной линии с пучками волокон слухового нерва. Эти основные клетки возбуждающие.
Другое название этих клеток - «чопперы». У них есть собственный ритм, и они будут запускать потенциалы действия с этим ритмом, когда они будут активированы правильным звуком.
Переднее вентральное кохлеарное ядро (AVCN)
Антеровентральное ядро улитки (AVCN) (или аксессуар), находится между двумя разделами кохлеарный нерв, и находится на вентральной части нижний цветонос.
- AVCN можно подразделить на основе цитоархитектура области.
- Типичные подразделения определяются как переднее (AaVCN), заднее (PaVCN), постеродорсальное (PDaVCN) и задневентральное (PVaVCN). [3]
Рекомендации
Эта статья включает текст в всеобщее достояние из стр. 788 20-го издания Анатомия Грея (1918)
Анатомия и физиология вентрального кохлеарного ядра (ВКЯ)
Анатомия и физиология вентрального кохлеарного ядра (ВКЯ)
Как описано отдельной статье «Передача звука во внутреннем ухе в норме», сложный звук распадается на составные частотные компоненты путем механической настройкой улитки. Поскольку акустическая энергия тонотопически распределена вдоль кохлеарной части, выходные частоты отдельных волокон слухового нерва зависят от формы волны. Дополнительный процесс восстановления звукового сигнала из рассеянных периферических сигналов начинается в улитковом ядре, где параллельные потоки восходящей информации, каждый с собственной уникальной функциональной ролью, представлены сходящимися структурами волокон слухового нерва.
Следующее обсуждение строится вокруг самой широкой функциональной дихотомной структуры: вентрального и дорсального разделения комплекса улитковых ядер. Это разделение возникло в самом начале анатомического формирования слухового отдела ствола мозга. Такое разделение сохранилось лишь по причине того, что представленная клеточная морфология обеспечила превосходное функционирование на начальных этапах центральной слуховой обработки.
Вентральное кохлеарное ядро (ВКЯ) служит первичной точкой входа для эфферентных путей к верхней оливе, латеральной петле и нижнему коленцу (НК). Слуховая информация идет к каждой из этих структур в определенных проводящих путях, которые приспособлены для селективного кодирования фаз и уровней частотных компонентов, а также и более глобальных свойств, таких как амплитуда модуляции стимула. Это функциональное разделение происходит из-за различий в синаптических структурах, конвергирующих структурах и внутренних электрических свойств нейронов ВКЯ.
а) Анатомия вентрального кохлеарного ядра. Приблизительно 30000 слуховых нервных волокон соединяют человеческое внутреннее ухо с комплексом кохлеарных ядер. После поступления в ствол мозга пути разделяются на восходящую ветвь, которая идет к переднему ВКЯ и нисходящую ветвь, которая проходит через заднее ВКЯ и дорсальное кохлеарное ядро (ДКЯ). Хотя расположение проекций повторяет тонотопическую организацию улиткового разделения, линейное отображение частоты преобразуется в конечные поля. Посредством каждой двухмерной частотной пластинки ортогональная ось кодирует другие показатели звука.
Завершение слухового нерва в кохлеарном ядре имеет принципиальное значение. Вся информация о звуке должна быть передана к вышестоящим центрам посредством выходных частот слухового ядра. Для каждого слухового нервного волокна есть примерно два нейрона ВКЯ, что предполагает высокую степень схождения в ядре.
Каким образом нейроны будут интегрировать многообразные входящие сигналы слухового нерва, зависит от физических характеристик синапсов улитковых ядер. Известны две базовые структуры. Нейроны с широкими интеграционными свойствами имеют обширные дендритные поля, покрытые шишковидными окончаниями большого числа волокон слухового нерва. Нейроны, которые точно сохраняют образцы сигналов слухового нерва, управляются большими аксональными окончаниями, концевыми утолщениями Хелда. Эти входы малочисленны, но по отдельности достаточно мощные, чтобы активизировать постсинаптическую активность.
Основные подтипы нейронов ВКЯ отличаются тремя основными типами клеток. Мшистые клетки имеют мелкие, похожие на куст, дендриты, глобулярные или сферические тела клеток и синаптические концевые утолщения. Как большинство нейронов в центральной нервной системе, мультиполярные (или звездчатые), клетки имеют длинные, практически без ветвей, дендриты и шишковидные синапсы. Интеграционные свойства этих клеток прежде всего определены тем, лежат ли их дендритные области в пределах плоскости частотной пластины или проходят через нее. Так называемые «осьминогообразные» клетки имеют длинные дендриты, с пучками от одной стороны тела клетки, напоминая головоногих.
Цитоархитектоника ВКЯ организована регионально. Сферические мшистые клетки, располагаясь в самом переднем отделе, окружают точку входа слухового нерва. Глобулярные мшистые клетки занимают промежуточное местоположение, в то время как «осьминогообразные» клетки расположены в заднем отделе. Мультиполярные клетки, как наиболее структурно разнообразные нейроны, в основном рассеяны.
![Комплекс кохлеарного ядра]()
Топографическая группировка морфологических нейрональных классов в комплексе кохлеарного ядра.
Схема показывает относительное положение кохлеарного ядра в пределах центральных слуховых путей.
ДКЯ —дорсальное кохлеарное ядро; НД — нижнее двухолмие; ЛСО — латеральная верхняя олива;
МКТ — медальное коленчатое тело; МЯТТ — медиальное ядро трапециевидного тела;
МВО — медиальная верхняя олива; ВКЯ — вентральное кохлеарное ядро.б) Основные физиологические свойства вентрального кохлеарного ядра (ВКЯ). Анатомическое распределение определенных нейронных групп позволило физиологам связывать анатомическую специализацию нейронов вентрального кохлеарного ядра (ВКЯ) с их свойствами кодирования звука. Ответы в переднем ядре наиболее вероятно регистрируются от мшистых клеток, в то время как реакции в заднем ядре зарегистрируются от «осьминогообразных» клеток. Эти предположения были подтверждены внутриклеточными экспериментами, которые охарактеризовали физиологические свойства нейронов кохлеарного ядра перед маркировкой для последующей визуализации клетки материалом, таким как пероксидаза корней хрена (ПКХ).
Первичные нейроны дают гистограмму с отражением относительно выраженного ответа, но быстро адаптирующегося к более низкой устойчивой частоте. Так как первичные нейроны обнаруживаются в передних отделах ВКЯ, они объединяются для функционирования подобно кустистым клеткам. В соответствии с высокой стабильностью концевых утолщений, которые связывают кустистые клетки с волокнами слухового нерва, физиологические характеристики первичных нейронов тесно связаны с их периферическими выходами.
Первичные нейроны с желобком обнаружены в областях, содержащих большие количества глобулярных мшистых клеток. Их перистимульная гистограмма содержат серию острых пиков, которая следует за кратким периодом неактивности. Этот «желобок» отражает рефракционный период нейрона после возникновения потенциала действия. Первично подобные нейроны с вырезкой обладают этим свойством, потому что измененные концевые окончания синхронизируют их ответы на начало стимула.
«Начальные» нейроны названы таким образом из-за своего свойства возбуждаться в начале стимула и затем показывать низкую активность к моменту стихания стимула. Начальные нейроны-запиратели (начальные-3), как правило, регистрируются в задних отделах ВКЯ, их морфологическая связь с «осьминогообразными» клетками была установлена посредством внутриклеточной маркировки. Точное возникновение ответа, широкая настройка частоты и широкий динамический диапазон начальных нейронов согласуются с высокой конвергенцией афферентных входов «осьминогообразных» клеток.
Начальные нейроны-прерыватели (начальные-П) менее вариабельны, чем начальные-3 нейроны. Поскольку их потенциалы действия возникают с регулярными интервалами, начальные-П нейроны дают «прерывистую» ПСТГ. Эта реакция приписывается D-звездчатым нейронам, которые являются большими мультиполярными клетками, посылающими дорзальные двусторонние проекции в вентральном кохлеарном ядре (ВКЯ). Эти нейроны также называют «лучистыми» нейронами т.к. их дендритные поля располагаются в трех измерениях, объединяющих входы слухового нерва через широкий диапазон частот. Оказывается, они имеют один источник глицинэргического торможения, который играет важную роль в кодирующих свойствах звука в кохлеарном ядре.
Клетки-прерыватели — физиологическая копия Т-звездчатых клеток. Эти маленькие мультиполярные клетки распространяются вентрально через трапециевидное тело к контралатеральным нижним бугоркам. Эти нейроны также описываются как «плоские» нейроны, так как их дендритные поля ориентированы в плоскости частотой пластины. В дополнение к острой настройке на частоту, прерывистые элементы отражают крайне регулярную частоту выброса и поэтому производят «прерывистый» ПСТГ. Объединяя ответы многих слуховых нервных волокон, клетки-прерыватели производят сигналы, которые устойчивы к изменению уровня звука и эффектам фонового шума.
в) Кодирование звука в вентральном кохлеарном ядре. Разнообразные физиологические структуры групп клеток в вентральном кохлеарном ядре (ВКЯ) наделяют нейроны кодирующими способностями с избирательным подходом к акустическим особенностям сложных звуков. Кроме «начальных» нейронов, проекционные нейроны имеют тенденцию к четкой настройке на частоту. Следовательно, в слуховом нерве сложные спектры с многообразными частотными компонентами должны быть закодированы исходящими частотами множества нейронов с дополнительными возможностями настройки.
С точки зрения человеческого слухового опыта обработка сложных спектральных форм — это основа речевого восприятия.
В английском языке самым элементарным спектром является устойчивый гласный. Перцептивное распознавание гласного звука определяется частотами его формант, макроэргически связанными со стимулами. Эти несущие информацию элементы закодированы сотнями нейронов в слуховом нерве и кохлеарном ядре, объединяющиеся для создания удивительно ясной картины, в которой исходящая частота линейно связана с количеством спектральной энергии в пределах частотного диапазона каждого нейрона.
Кривая гласного звука появляется, когда активность, вызванная гласным звуком, отражается на наилучших частотах (НЧ) нейрона. Например, английский гласный /е/ (как русское /э/), как в слове «bet», вызывает высокочастотные выходные сигналы около 0,5 и 1,7 Гц, которые частотно локализуются у его первой и второй формант.
Длительные и кратковременные единицы прерывности обеспечивают превосходные уровни представления спектров гласного звука в широком диапазоне уровней звука. Предполагается, что этот расширенный динамический диапазон обеспечивается плотным схождением слуховых нервных волокон на многополюсных клетках.
Волокна с высокими спонтанными частотами (СЧ) зачастую имеют низкие пороги чувствительности. Единицы прерывности могут ослаблять эти входы на низких уровнях звука. С другой стороны, волокна с низкими СЧ имеют высокие пороги чувствительности, которые могут доминировать над прерывными ответами на высоких уровнях звука. Были зафиксированы обходные пути данного «избирательного слушания», при котором высокочастотные входы локализуются на дистальных дендритах, а низкочастотные входы располагаются около тел прерывающих клеток.
При звуках высокой интенсивности активация промежуточных тормозящих входов из радиальных нейронов отводит насыщенные высокочастотные импульсы от тел клеток. Ответный механизм для низкочастотных импульсов не требуется, так как их действие ограничено порогами чувствительности при низких уровнях шума.
Репрезентация гласных звуков первично-подобными нейронами ВКЯ демонстрирует ограниченность динамического диапазона входов их слуховых нервов. Первично-подобные нейроны с низкой СЧ обеспечивают скудное представление о формантной структуре на низких уровнях звука, потому что они неэффективно управляются стимулом. Структура гласных звуков проявляется на надпороговом уровне, так как наивысшая исходящая частота ограничена формант-настраивающими нейронами. Первично-подобные нейроны с высокой СЧ обеспечивают хорошую репрезентацию формантной структуры на низких уровнях, но плохой профиль на высоких из-за того, что полный набор нейронов реагирует на максимальной частоте.
Пороги и эффекты насыщения первично-подобных нейронов ограничивают их способность к кодированию спектральной формы (исходящей частоты), однако их ответы богаты временной информацией. В частности, временная точность потенциалов действия является критичной для обобщения пространственной локализации слуховых стимулов.
Большая часть естественных звуков представляют собой краткие переходные процессы. Исходные нейроны активно отвечают на эти стимулы и могут участвовать в тесно-связанных сенсорно-двигательных проводящих путях, которые контролируют реакцию на внезапный акустический стимул и ответные движения. Бинауральное сравнение времени начала стимулов также оказывает сильное влияние на пространственное восприятие. Результат не обязательно должен возникать с началом стимула. Резкий акустический переход стимула по огибающей вызовет подобную реакцию.
Если переходные процессы повторяются с постоянной частотой, то реакции исходных нейронов будут определяться периодом стимуляции. Периодичность — фундаментальная характеристика уровня восприятия и важнейший сигнал для выделения звуковых сигналов из фонового шума.
г) Симптомы и клиника патологии вентрального кохлеарного ядра (ВКЯ). Анализ неврологических расстройств человека был важен для объяснения физиологических основ более высоких когнитивных функций. Текущее понимание локализации центров речи и понимания языка в коре головного мозга можно проследить вплоть до описанных Broca и Wernicke в XIX веке пациентов, страдающих афазией. Более широкое определение клинических проявлений проводящих путей, несущих информацию к коре, было получено при хирургических манипуляциях на экспериментальных животных.
Функциональные нарушения ВКЯ были исследованы путем повреждения путей, которые выходят из ядра и проходят через трапециевидное тело. Итоговые отклонения были по тяжести сопоставимы с последствиями удаления улитки или слухового нерва. Если повреждение трапециевидного тела выполнялось в передне-среднем отделе ствола мозга, то устранялись сигналы от обоих кохлеарных ядер и субъект оказывался глухим. При простом латерализованном повреждении терялся сигнал от одного ВКЯ. Субъекты поддерживали слуховую функцию в одном ухе, но они претерпевали серьезную потерю направленного слушания, потому что мозг больше не проводил бинаурального сравнения источника стимулов.
Временное выключение входящих в ВКЯ стимулов может быть причиной долговременных клинических проявлений. На ранних стадиях развития для установления связей с улитковым ядром требовался звук, активирующий слуховой нерв. Если ухо выключено из-за кондуктивной потери слуха или сенсоневральных нарушений, концевые окончания Хелда подвергаются гипертрофии, которая ассоциируется с временным дефицитом обработки. Синапсы и, вероятно, высшие когнитивные функции, могут быть сохранены срочными вмешательствами, направленными на восстановление поступления входных сигналов к кохлеарному ядру. Эти данные, полученные на моделях животных, могут объяснить, почему кохлеарная имплантация и установка слухового аппарата наиболее эффективны в молодом возрасте.
![Схема проводящих путей слуха]()
Схема восходящих путей центральной слуховой системы.
Основные связи между главными ядрами показаны для левого уха.
Симметричная проекция для правого уха не показана.Учебное видео проводящих путей слухового анализатора
Редактор: Искандер Милевски. Дата обновления публикации: 18.3.2021
Проводниковый и корковый отделы слуховой сенсорной системы
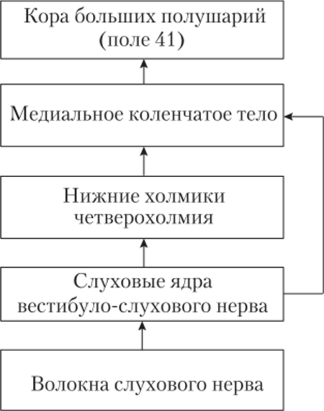
нице продолговатого мозга и моста, — дорсальном и вентральном. Большинство волокон от этих ядер перекрещиваются (переходят на другую сторону) и образуют синапсы на слуховых центрах противоположной стороны. Основная часть волокон от вентрального ядра идет к ядрам верхних олив — слуховым ядрам в варолиевом мосту. Оттуда волокна идут к нижним холмикам четверохолмия среднего мозга. Следующий центр проводникового отдела — проекционные сенсорные ядра таламуса (промежуточный мозг) — медиальные (наружные) коленчатые тела (МКТ). Волокна от этих ядер идут в височную долю коры больших полушарий, образуя так называемую слуховую лучистость.
![Упрощенная схема путей и центров слуховой системы]()
Рис. 14.6. Упрощенная схема путей и центров слуховой системы:
явление перекреста не учитывается
Волокна от дорсальных кохлеарных ядер также совершают перекрест и идут в центры среднего мозга и таламуса, минуя верхние оливы. Небольшая часть волокон от кохлеарных ядер поступает в МКТ напрямую, т.е. без переключения в ядрах верхних олив и нижних холмиках. Перекрест некоторых слуховых волокон осуществляется и на других уровнях слуховой системы.
Волокна, идущие от кохлеарных ядер и верхних олив, в варолиевом мосту собираются в один пучок — латеральный лемниск (латеральная петля). Большинство волокон этого пучка, как ясно из вышесказанного, заканчивается на нейронах нижних холмиков, небольшая часть сразу идет в МКТ.
Каждое проводящее волокно слухового нерва передает информацию от строго определенных волосковых клеток. В результате отдельные волокна слухового нерва также возбуждаются при предъявлении звука только в определенном частотном диапазоне. По ним информация о звуковых сигналах передается в кохлеарные ядра. В них есть полное представительство («карта») улитки; расположение переключающих нейронов тонотопическое (частота звука кодируется «номером канала»). Тонотопия сохраняется и на всех других уровнях слуховой системы.
Локализация источника звука определяется при помощи бинаурального слуха, т.е. слышания обоими ушами. Каждый звук, идущий сбоку, достигает более удаленного уха позже и с меньшей силой. Благодаря перекресту волокон информация от рецепторов правого и левого уха может конвергировать на одних и тех же нейронах. Это впервые происходит в ядрах верхнеоливарного комплекса, затем анализ продолжается на более высо- колежащих уровнях. Бинауральный слух дает возможность определять направление звука с точностью до 3—4°. Человек, глухой на одно ухо (с моноуральным слухом), может сориентироваться в направлении звука, только вращая головой и оценивая громкость сигнала.
Первичная слуховая кора находится в височной доле больших полушарий (поле 41; см. рис. 9.9). Первичная слуховая кора получает проекции от медиального коленчатого тела. В коре есть несколько представительств (карт) улитки, в результате чего осуществляется параллельная обработка информации. В слуховой коре, как и в некоторых других сенсорных зонах, обнаружены слуховые колонки как структурные элементы коры, обеспечивающие первичные процессы опознавания звукового сигнала. Все нейроны одной колонки имеют одну и ту же оптимальную частоту звука, на который они отвечают.
Поле 41 тесно связано с окружающей его вторичной слуховой корой (поля 42 и 22; см. рис. 9.9). У человека повреждение этих зон ведет к нарушению восприятия музыки, речи и т.п. Так, именно здесь в левом полушарии у правшей находится речевой центр Вернике. При поражении этого центра наблюдается сенсорная афазия — нарушение понимания звучащей речи. В экспериментах на обезьянах показано, что здесь находятся нейроны, связанные с внутривидовой коммуникацией (общением) и опознающие издаваемые при этом звуки. По-видимому, большинство нейронов, находящихся в этой зоне, обладают свойством настраиваться на определенные звуковые образы, т.е. проходят процесс обучения.
Нарушения в работе слухового анализатора могут быть связано как с периферическим, так и с центральными отделами. Например, при воспалении среднего уха часто нарушается костная передача звука из-за снижения подвижности слуховых косточек. При воспалении внутреннего уха возможно поражение слуховых рецепторов, в результате чего нарушается трансформация звукового сигнала в нервный импульс. Сильные шумы, постоянно действующие на ухо, наносят ему большой вред, так как барабанная перепонка при этом колеблется с большим размахом и теряет свою эластичность. В результате развивается тугоухость — профессиональная болезнь людей, работающих в условиях повышенного шума.
«Проект: Аудиология» - сборник научно-практических материалов для профессионалов в области аудиологии и слухопротезирования, включающий материалы конференций, статьи по кохлеарной имплантации, отоневрологии и законодательные акты по сурдологии.![Проект - Аудиология]()
Слуховой стволомозговой имплантант (ABI, auditory brainstem implant) представляет собой модифицированный кохлеарный имплант, и предназначен для электрической стимуляции кохлеарных ядер ствола мозга пациента.
ABI был разработан специально для пациентов с заболеванием, известным как нейрофиброматоз II типа (НФ-2). НФ-2 характеризуется наличием двусторонних вестибулярных шванном. Сама опухоль или хирургическое удаление опухоли часто приводит к значительному поражению слухового нерва, поэтому кохлеарная имплантация не является методом выбора для этих пациентов. После удаления опухоли электродный массив ABI вводится в латеральный карман четвёртого желудочка и размещается на поверхности вентрального и дорсального кохлеарных ядер. Стимулируя слуховые нервные структуры в пределах кохлеарного ядра, ABI передаёт важную информацию о частоте, амплитуде и временных характеристиках звуков.
Дизайн и функционирование ABI подобны многоканальному кохлеарному импланту, за исключением различий в дизайне стимулирующего электродного массива 5. Программирование устройства ABI отличается от программирования кохлеарного импланта по некоторым важным аспектам.
В статье внимание сфокусировано на многоканальной высокочастотной системе ABI производства MED-EL.Техническое описание системы C40+ ABI
Система состоит из многоканальной имплантируемой части и речевого процессора.
Имплантируемые части системы C40+ ABI включают в себя ABI-стимулятор (приёмник/передатчик), активный электродный массив и референтный электрод.![Слуховая стволомозговая имплантация]()
Стимулятор
Приблизительные размеры стимулятора С40+ 3,5 х 2,4 см, толщина менее 0,4 мм. Все электронные компоненты помещены в твёрдый и компактный керамический корпус (Рисунок 1), в котором герметично запаяны электронная схема и мощный микрочип. Корпус импланта и электроника ABI аналогична тем, которые используются в кохлеарном импланте С40+.
Рисунок 1. Слуховой стволомозговой имплант.Активный электродный массив ABI.
От стимулятора отходит активный электродный массив, который состоит из 12 активных платиновых контактов, запаянных в овальную силиконовую капсулу-пластинку в форме весла с размерами (5,5 х 3,0 х 0,6) мм (Рисунок 2, середина). На обратной стороне силиконовой капсулы имеется сеточка из дакрона (синтетическая нерассасывающаяся нить), которая облегчает фиксацию электродного массива.
Рисунок 2. Электроды слухового стволомозгового импланта (MED-EL, Combi 40+R). (Верх): Четырёхканальный тест-электрод для локализации кохлеарного ядра. (Середина): Двенадцатиканальный активный электрод, который будет имплантирован.![Слуховая стволомозговая имплантация]()
Электродный массив преформирован идущими поперёк платиновыми проводками. Это позволяет так сформировать этот элемент, чтобы он подходил к контуру (поверхности) кохлеарного ядра (Рисунок 3).
Рисунок 3. Активный электродный массив слухового стволомозгового импланта.Референтный электрод
Референтный электрод в форме листа клевера идентичен тому, который используется в кохлеарном импланте C40+ (Рисунок 2, низ).Внешними компонентами ABI производства MED-EL являются заушный речевой процессор TEMPO+ (Рисунок 4) или карманный речевой процессор CIS PRO+.
Рисунок 4. Полностью заушный речевой процессор TEMPO+.Речевой процессор разработан для того, чтобы кодировать акустические сигналы окружающей среды, которые собираются микрофоном. Акустический сигнал преобразовывается в электрический сигнал и превращается в паттерн электрических импульсов в соответствии с используемой стратегией кодирования речи. От речевого процессора закодированные электрические импульсы передаются через передающую катушку, через интактную (неповреждённую) кожу к импланту. Электроника декодирует сигналы, полученные внутренней вторичной катушкой. Эти сигналы передаются затем активному электродному массиву, и кохлеарное ядро стимулируется.
В ABI MED-EL используется стратегия обработки речи CIS (continuous interleaved sampling, стратегия высокочастотной непрерывной выборки), разработанная в 90-х годах XX столетия Вильсоном (Wilson) с партнёрами. С этим устройством возможна высокочастотная симуляция с максимумом в 18180 имп./с. Максимальная частота стимуляции на канал составляет 515/с при 12 активных электродах. Режим стимуляции является монополярным, против референтного электрода, который помещается под височную фасцию.Показания, отбор кандидатов для ABIs
Когда стволомозговая имплантация только начиналась, ABIs использовались почти исключительно только у пациентов с нейрофиброматозом II типа, которые потеряли слух до или во время удаления двусторонней вестибулярной шванномы. Позже показания в Европе для ABI стали включать аплазию кохлеарного нерва (в том числе и вследствие атрезии внутреннего слухового прохода) и выраженную мальформацию улитки у детей, а также полную оссификацию улитки или разрыв кохлеарного нерва из-за травмы у взрослых 9.
При отборе должна учитываться индивидуальная мотивация пациента для повторных послеоперационных процедур настройки и его/её готовность к ежедневным занятиям с устройством. Пациенты должны быть полностью информированы о возможных результатах чтобы избежать любого разочарования, которое может отразиться на их общей мотивации или готовности к сотрудничеству. Кандидаты должны иметь нормальный уровень интеллектуального развития. Проявления основной болезни не должны помешать процессу настройки или реабилитации. По мнению некоторых авторов, НФ-2 типа Wischard, при котором есть выраженные признаки и симптомы и уменьшенная продолжительность жизни, не должен считаться противопоказанием. За и против имплантации при этом подтипе НФ-2 должны взвешиваться у каждого конкретного пациента. Если пациент в физическом и психическом плане способен и согласен пройти все необходимые послеоперационные процедуры настройки и реабилитации и имеет хорошую поддержку со стороны семьи, нет никакой причины отказывать ему/ей в шансе улучшить слух. [10].Периост и фасция височной мышцы рядом с ним отводятся кпереди чтобы сформировать второй слой, который прикроет имплант. Кость вскрывается в ретромастоидальной области и обнажается твёрдая мозговая оболочка за сигмовидным и ниже поперечного синусов. Полушария мозжечка отводятся после дренирования цереброспинальной жидкости (ЦСЖ) при вскрытии базальных цистерн. Арахноидальная оболочка, покрывающая опухоль, вскрывается, на опухоль накладывается зажим и она постепенно удаляется. В зависимости от размера опухоли и интраоперационных находок, внутренний слуховой проход рассверливается до, во время или после удаления экстрамеатальной порции опухоли. После полного удаления опухоли каудальные черепно-мозговые нервы выделяются из арахноидальной оболочки. Открываются флоккулюс мозжечка и хориоидное сплетение бокового кармана четвертого желудочка. Доступ к отверстию Люшка (Luschka) облегчается поворотом головы в сторону опухоли. В отверстие Люшка вводится четырёхполярный стимулирующий тест-электрод (Рисунок 2, верх) и размещается в латеральном кармане четвёртого желудочка, на поверхности кохлеарного ядра. Главная цель – вентральное кохлеарное ядро.
Расположение тест-электрода ABI выполняется в соответствии с анатомическими ориентирами, которые доступны обзору хирурга. Анатомические ориентиры, использующиеся для размещения импланта, включают корешок VIII нерва, языкоглоточный нерв, лицевой нерв и хориоидальное сплетение, а также вход в латеральный карман (латеральная апертура, отверстие Люшка (Luschka)), где все эти структуры сходятся (Рисунки 6-8). При операции, когда практически всегда ствол мозга искажён опухолью, отверстие Люшка расположено выше IX нерва. IX нерв обычно находится в фиксированном анатомическом положении, приводящем практически всегда в отверстие Люшка. Кроме того, важна луковица яремной вены, потому что её расположение может очень сильно варьироваться.
Рисунок 6. Анатомия кохлеарных ядер: анатомический препарат (левая сторона) показывает внутренний слуховой канал (IAC), VIII и IX черепно-мозговые нервы, сходящиеся ко входу в латеральный карман (отверстие Люшка), где имеется сплетение (Т). Показана локализация вентрального (VCN) и дорсального (DCN) кохлеарных ядер. С = каудально; D = дорсально; R = рострально; V = вентрально.![Слуховая стволомозговая имплантация]()
Рисунок 7. На гистологическом препарате показаны область латерального кармана четвёртого желудочка (Ve), VII (VIIn), VIII (VIIIn), IX (IXn) черепно-мозговые нервы и взаимоотношения между вентральным кохлеарным ядром (VCN), дорсальным кохлеарным ядром (DCN), нижней ножкой мозжечка ICP) и вестибулярными ядрами (VN). Также показаны мозжечок (CE) и нижние оливы (IO). A = кпереди; Р = кзади.
Рисунок 8. Показана область имплантации на препарате. Электрод (Е) виден в латеральном кармане в области кохлеарного ядра; отверстие Люшка (Luschka) окружено (обрисовано) VIII (VIIIn) и IX (IXn) черепно-мозговыми нервами; ножки мозжечка (Се) ограничивают его сверху; хориоидальное сплетение (Ch) обычно выходит из бокового кармана. С = каудально; D = дорсально; R = рострально; V = вентрально.![Слуховая стволомозговая имплантация]()
В затылочно-теменной области выше поперечного синуса формируется костное ложе для приёмника-передатчика (стимулятора) – таким образом, чтобы приёмник/передатчик был погружён в кость по крайней мере на половину своей толщины, на ~ 2 мм. Внимательно следят за тем, чтобы не повредить внутренний кортикальный слой кости. Приёмник/передатчик фиксируется звёздчатым швом, позже он покрывается фасцией и периостом, после чего кожный лоскут укладывается на место и ушивается двумя слоями швов.
Затем тест-электрод в латеральном кармане ещё раз проверяется. Измерения EABR в каждом из вариантов режима биполярной стимуляции в пределах расположения четырёх электродов повторяют, пока результаты всех возможных биполярных комбинаций из четырёх контактов не покажут адекватные волны EABR. После этого тест-электрод удаляют и из стерильной упаковки извлекают 12-ти-канальный электродный массив ABI (Рисунок 2, середина, Рисунок 3) (активный электрод). Затем активный электродный массив размещают в соответствии с положением, намеченным регистрацией EABR при помощи тест-электрода. Перед тем как зафиксировать активный электрод, с него проводится окончательная запись EABR (Рисунок 10).
Рисунок 10. ABR: акустические ABR (а), электрические EABR у пользователя кохлеарным имплантом (b), электрические EABR у пациента с ABI (c)Хотя КТ-сканирования достаточно для контроля за пациентами с ABI, большинство пациентов требуют проведения МРТ в связи с их основным заболеванием (НФ-2). Тайссл (Teissl) с сотрудниками, затем Хохмайр (Hochmair) (Университет Инсбрука, 2001) показали, что МРТ при 0,2 и 1,5 Т не оказывает никакого отрицательного влияния на импланты MED-EL [10, 16].
Доктора Уильям Хауз (William House) и Уильям Хитцельбергер (William Hitselberger) первыми использовали одноканальный имплант, получивший в дальнейшем название ABI, для электрической стимуляции кохлеарного ядра у пациента с НФ-2 в 1979 году 2. Перед имплантацией было доказано, что электрическая стимуляция кохлеарного ядра у людей могла приводить к слуховым ощущениям. Первое устройство ABI состояло из пары шаровидных электродов, которые помещались в толщу кохлеарного ядра. Управлял электродами модифицированный слуховой аппарат Bosch. Однако, эта пара электродов смещалась и вызывала неслуховые побочные эффекты. При операции-ревизии в 1981 году этому пациенту в латеральный карман четвёртого желудочка, на поверхность кохлеарного ядра, был имплантирован электродный массив в форме весла с двумя электродами. Вплоть до 1991 года использовался этот тип электродов и все созданные в последнее время электродные массивы использовали его как прототип и имплантируются также в латеральный карман. С тех пор устройство претерпело несколько модификаций – каналов стало больше (теперь доступны системы с от 8 до 21 каналов) 26, разработаны новые стратегии кодирования звукового сигнала, появляются новые варианты электродов, такие как пенетрирующий ABI. Главной движущей силой для усовершенствования был успех операций кохлеарной имплантации. Другими предпосылками стали развитие микрохирургической техники, интраоперационного мониторинга и нейроанестезии. Также произошли изменения и в отношении хирургов: они больше не удовлетворены просто удалением акустической невриномы, сейчас они хотят сохранить функцию. Первоначально это желание относилось к лицевому нерву, сейчас оно распространяется и на кохлеарный нерв [17].
ABI MED-El был разработан и произведён в ходе сотрудничества инженеров MED-EL Инсбрукского университета, Австрия, и Вюрцбургского университета, Германия на основе кохлеарного импланта Combi 40/40+ (компания MEDEL, Инсбрук, Австрия), который был оценён в европейском мультицентровом клиническом исследовании [18]. Не так давно MED-EL разработан ABI нового поколения – Pulsar i100 ABI.
Стволомозговой имплант фирмы Cochlea был разработан совместно компанией Cochlear, House Ear Institute (Лос-Анджелес, Калифорния, США) и Медицинским Исследовательским Институтом (Хантингтон, США). Авторами считаются Роберт Шеннон и Дералд Брекмен. 20 октября 2000 г. изделие было одобрено FDA (Управлением по контролю за продуктами и лекарствами США).
Исследования не стоят на месте. Ранние исследования электрической стимуляции нижних бугров четверохолмия у человека не обеспечивали ощущения звука [19]. Однако Коллетти (Colletti) и др. [20, 23] недавно имплантировал электроды в нижние бугры четверохолмия пациенту с двусторонним дефектом слухового нерва вследствие удаления вестибулярной шванномы, продемонстрировав, что электрическая стимуляция нижних бугров четверохолмия действительно может обеспечить восприятие звука и некоторую разборчивость речи.
В House Ear Institute (Лос-Анджелес, Калифорния, США) и Huntington Medical Research Institute (Пасадена, Калифорния, США) в сотрудничестве с производителем, Cochlear Corporation, с целью улучшения точности стимуляции слуховых нейронов ствола мозга разрабатывается и проходит испытания PABI (пенетрирующий ABI), состоящий из двух электродных массивов: 12-ти-электродного поверхностного массива и 10-ти-электродного массива с игольчатыми микроэлектродами, который размещается в толще вентральной порции кохлеарного ядра. Считается, что микростимуляция с игольчатыми электродами обеспечит активацию малых скоплений тонотопических групп нейронов в глубоких слоях с восприятием целого диапазона частот (высот звука) [21].
К настоящему времени в мире устройство ABI получили более 500 пациентов. Операции активно проводятся в институтах Европы, США, Ближнего Востока, Азии.Возможные осложнения, побочные эффекты
Кроме осложнений, непосредственно связанных с имплантацией, а именно: хирургических повреждений нижележащих черепно-мозговых нервов, сосудистых повреждений или геморрагий в области ствола мозга, в литературе описано развитие на 7-20 день после операции псевдомембраноцеле. Поскольку важно расстояние между передающей катушкой и имплантированным передатчиком, это осложнение нужно лечить, даже если оно (лечение) не требовалось бы неимплантированным пациентам [10].
Описан случай развития у пациента гидроцефалии, которая потребовала вентрикулоперитонеального шунтирования через 2 месяца после первой операции [10]. У этого пациента была глиома в области brachium pontis мозжечка на контралатеральной от операции стороне и бессимптомная солидная и кистозная глиома нижних отделов ствола мозга, которые, возможно, нарушали циркуляцию цереброспинальной жидкости.
Несмотря на хорошие интраоперационные EABR, иногда только несколько слуховых электродов может функционировать после операции. Это может быть связано с движением (перемещением, смещением) электродного массива после операции или, в некоторых случаях, корешок VIII нерва, возможно, временно давал хорошие ответы EABRs в операционной, но после его атрофии ответы не регистрировались [15].
При подключении речевого процессора и первых его настройках крайне важно следить за жизненными показателями пациента; подключение и настройку следует делать в отделении интенсивной терапии или в присутствии анастезиолога-реаниматолога, т.к. в литературе описаны случаи стимулирования блуждающего нерва, что может привести к остановке сердца [15].
Как было показано в ходе психофизиологического тестирования во время первичной настройки ABI, более 90% пользователей слухового стволомозгового импланта испытывают какие-либо неслуховые побочные эффекты [22]. Они появляются после подключения речевого процессора и активации импланта. К ним относятся покалывание в горле, покалывание или подёргивание в туловище или в верхних и/или нижних конечностях, раздражение лицевого нерва, подёргивание глаз или рта, размытость (нечёткость) полей зрения, затруднения при глотании или боль. Неслуховые эффекты могут быть уменьшены или устранены путём регулирования программных параметров или выключением канала [14].Восстановление слуха с использованием стволомозговых имплантов у пациентов с НФ-2 и другими состояниями, при которых нарушена целостность слухового нерва – это надёжная и многообещающая процедура для тех, кто иначе был бы полностью глухим.
Читайте также: