Лизосомы. Этапы эндоцитоза. Переваривание при эндоцитозе.
Добавил пользователь Morpheus Обновлено: 27.01.2026
Лизосома — (от λύσις — растворяю и sōma — тело) клеточный везикул. Эти одномембранные органоиды - часть эндомембранной системы клетки ). Разные виды лизосом могут рассматриваться как отдельные клеточные компартменты.
Содержание
Распространенность среди царств живой природы
Лизосомы были впервые описаны в 1955 году Кристианом де Дювом в животной клетке, а позже были обнаружены и в растительной. У растений к лизосомам по способу образования, а отчасти и по функциям близки вакуоли. Лизосомы есть также у большинства протистов (как с фаготрофным, так и с осмотрофным типом питания) и у грибов.Таким образом, наличие лизосом характерно для клеток всех эукариот. У прокариот лизосомы отсутствуют, так как у них отсутствует фагоцитоз и нет внутриклеточного пищеварения.
Признаки лизосом
Один из признаков лизосом - наличие в них ряда белки, липиды и LAMP1 и LAMP2. Они присутствуют на мембране лизосом и поздних эндосом, но отсутствуют на мембранах других компартментов вакуома.
Образование лизосом и их типы
Лизосомы формируются из пузырьков (везикул), отделяющихся от аппарата Гольджи, и пузырьков (эндосом), в которые попадают вещества при эндоцитозе. В образовании аутолизосом (аутофагосом) принимают участие мембраны эндоплазматического ретикулума. Все белки лизосом синтезируются на "сидячих" рибосомах на внешней стороне мембран эндоплазматического ретикулума и затем проходят через его полость и через аппарат Гольджи.
Лизосомы - гетерогенные органеллы, имеющие разную форму, размеры, ультраструктурные и цитохимические особенности. "Типичные" лизосомы животных клеток обычно имеют размеры 0,1-1 мкм, сферическую или овальную форму. Число лизосом варьирует от одной (крупная вакуоль во многих клетках растений и грибов) до нескольких сотен или тысяч (в клетках животных).
К сожалению, нет общепринятой классификации и номенклатуры для разных стадий созревания и типов лизосом. Различают первичные и вторичные лизосомы. Первые образуются в области аппарата Гольджи, в них находятся ферменты в неактивном состоянии, вторые же содержат активные ферменты. Обычно ферменты лизосом активируются при понижении рН. Среди лизосом можно также выделить гетеролизосомы (переваривающие материал, поступающий в клетку извне - путем фаго- или пиноцитоза) и аутолизосомы (разрушающие собственные белки или органоиды клетки). Наиболее широко используется следующая классификация лизосом и связанных с ними компартментов:
- Ранняя эндосома - в нее поступают эндоцитозные (пиноцитозные) пузырьки. Из ранней эндосомы рецепторы, отдавшие (из-за пониженного рН) свой груз, возвращаются на наружную мембрану.
- Поздняя эндосома - в нее из ранней эндосомы поступают пузырьки с материалом, поглощенном при пиноцитозе, и пузырьки из аппарата Гольджи с гидролазами. Рецепторы маннозо-6-фосфата возвращаются из поздней эндосомы в аппарат Гольджи.
- Лизосома - в нее из поздней эндосомы поступают пузырьки со смесью гидролаз и перевариваемого материала.
- Фагосома - в нее попадают более крупные частицы (бактерии и т.п.), поглощенные путем фагоцитоза. Фагосомы обычно сливаются с лизосомой.
- Аутофагосома - окруженный двумя мембранами участок цитоплазмы, обычно включающий какие-либо органоиды и образующийся при макроаутофагии. Cливается с лизосомой.
- Мультивезикулярные тельца - обычно окружены одинарной мембраной, содержат внутри более мелкие окруженные одинарной мембраной пузырьки. Образуются в результате процесса, напоминающего микроаутофагию (см. ниже), но содержат материал, полученный извне. В мелких пузырьках обычно остаются и затем подвергаются деградации рецепторы наружной мембраны (например, рецепторы эпидермального фактора роста). По стадии формирования соответствуют ранней эндосоме. Описано образование мультивезикулярных телец, окруженных двумя мембранами, путем отпочковывания от ядерной оболочки.
- Остаточные тельца (телолизосомы) - пузырьки, содержащие непереваренный материал (в частности, Функции лизосом
- переваривание захваченных клеткой при эндоцитозе веществ или частиц (бактерий, других клеток) - уничтожение ненужных клетке структур, например, во время замены старых органоидов новыми, или переваривание белков и других веществ, произведенных внутри самой клетки - самопереваривание клетки, приводящее к ее гибели (иногда этот процесс не является паталогическим, а сопровождает развитие организма или дифференцировку некоторых специализированных клеток)
Внутриклеточное пищеварение и участие в обмене веществ
У многих протистов и у животных, имеющих ЛПНП, поступает внутрь пиноцитозных везикул после соединения ЛПНП с рецепторами ЛПНП на мембране. Рецепторы возвращаются к мембране из ранней эндосомы, а ЛПНП поступают в лизосомы. После этого ЛПНП перевариваются, а высвободившийся холестерин через мембрану лизосом поступает в цитоплазму.
Косвенно лизосомы участвуют в обмене, обеспечивая Аутофагия
Обычно различают два типа аутофагии - микроаутофагия и макроаутофагия. При микроаутофагии, как при образовании мультивезикулярных телец, образуются впячивания мембраны эндосомы или лизосомы, которые затем отделяются в виде внутренних пузырьков, только в них попадают вещества, синтезированные в самой клетке. Таким путем клетка может переваривать белки при нехватке энергии или строительного материала (например, при голодании). Но процессы микроаутофагии происходят и при нормальных условиях и в целом неизбирательны. Иногда в ходе микроаутофагии перевариваются и органоиды; так, у дрожжей описана микроаутофагия пероксисом и частичная микроаутофагия ядер, при которой клетка сохраняет жизнеспособность.
При макроаутофагии участок цитоплазмы (часто содержащий какие-либо органоиды) окружается мембранным компартментом, похожим на цистерну эндоплазматической сети. В результате этот участок оказывается отгорожен от остальной цитоплазмы двумя мембранами. Затем такая аутофагосома сливается с лизосомой, и ее содержимое переваривается. Видимо, макроаутофагия также неизбирательна, хотя часто подчеркивается, что с помощью нее клетка может избавляться от "отслуживших свой срок" органоидов (митохондрий. рибосом и др.).
Третий тип аутофагии - шаперон-зависимая. При этом способе происходит направленный транспорт частично денатурировавших белков из цитоплазмы сквозь мембрану лизосомы в ее полость.
Автолиз
Ферменты лизосом нередко высвобождаются при разрушении насекомых при апоптозом и Клиническое значение. Болезни, связанные с нарушением работы лизосом
Иногда из-за неправильной работы лизосом развиваются болезни накопления, при которых ферменты из-за мутаций не работают или работают плохо. Примером болезней накопления может служить амовротическая идиотия при накоплении гликогена.
Разрыв лизосомы и выход в гиалоплазму расщепляющих ферментов сопровождается резким повышением их активности. Такого рода повышение активности ферментов наблюдается, например, в очагах См. также
Лизосомы. Этапы эндоцитоза. Переваривание при эндоцитозе.
Лизосомы. Этапы эндоцитоза. Переваривание при эндоцитозе.
Лизосомы обнаруживаются почти во всех эукариотических клетках. Они представляют собой простые мембранные мешочки (стенка мешочка состоит из одинарной мембраны), наполненные пищеварительными ферментами — протеазами, нуклеазами и липазами, расщепляющими соответственно белки, нуклеиновые кислоты и липиды. Эти ферменты, катализирующие реакции гидролиза (идущие с присоединением воды), лучше всего работают в кислой среде; содержимое лизосом имеет, следовательно, кислую реакцию. Перечисленные выше ферменты должны быть отделены от всех остальных клеточных компонентов и структур, иначе они разрушат эти компоненты и структуры.
В животных клетках лизосомы имеют обычно округлую форму а диаметр их составляет 0,2—0,5 мкм.
В растительных клетках роль лизосом могут играть крупные центральные вакуоли. Впрочем, в цитоплазме растительных клеток иногда видны тельца, напоминающие по своему виду лизосомы животных клеток.
Заключенные в лизосомах ферменты синтезируются на гранулярном ЭР и транспортируются к аппарату Гольджи. Позже от него отпочковываются пузырьки Гольджи, содержащие ферменты, подвергшиеся необходимым модификациям. Эти пузырьки и превращаются в лизосомы. Лизосомы выполняют ряд функций, которые описаны ниже и перечислены на рисунке.
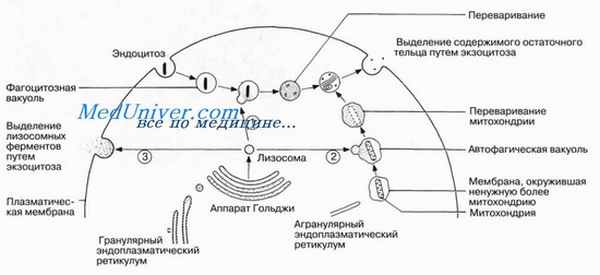
Переваривание материалов, поглощенных посредством эндоцитоза
Процесс эндоцитоза описан в нашей статье. Лизосомы могут сливаться с пузырьками или вакуолями, образующимися в результате эндоцитоза, высвобождать в них свои ферменты и переваривать находящиеся внутри пузырьков или вакуолей материалы. Иногда этот процесс представляет собой способ поглощения пищи, как, например, у некоторых простейших. В других случаях он выполняет защитную функцию, когда, например, специализированные лейкоциты (фагоциты) захватывают и переваривают попавшие в организм бактерии. Продукты переваривания поглощаются и усваиваются цитоплазмой клетки, но часть материала так и остается непереваренной. Вакуоль вместе с этим остаточным материалом направляется к плазматической мембране, и здесь ее содержимое выводится наружу путем экзоцитоза.
Своеобразную роль играют лизосомы в клетках щитовидной железы, которые путем пиноцитоза поглощают тиреоглобулин. Образовавшиеся пиноцитозные пузырьки сливаются с лизосомами, и тиреоглобулин под действием лизосомных ферментов превращается в активный гормон тироксин. После этого лизосомы путем слияния с плазматической мембраной изливают свое содержимое наружу — выделяют этот гормон в кровь.
Автофагия
Автофагией называется процесс, посредством которого клетка поглощает и переваривает внутри лизосом свои собственные не нужные ей структуры. Сначала эти структуры окружаются одинарной мембраной, отделяющейся обычно от агранулярного эндоплазматического ретикулума, а затем такой мембранный мешочек с заключенной в нем структурой сливается с лизосо-мой, образуя так называемую автофагическую вакуоль, в которой структура переваривается. Данная последовательность событий входит как составная часть в естественный круговорот цитоплазматических органелл, при котором старые органеллы заменяются новыми.
Выделение ферментов из клетки (экзоцитоз)
Иногда ферменты, содержащиеся в лизосомах, высвобождаются из клетки наружу. Это происходит, например, в процессе развития организма при замене хряшей костной тканью. Аналогичное явление можно наблюдать, когда основное вещество кости разрушается при перестройке костной ткани в ответ на повреждения, при новой нагрузке и т. п. В спермиях содержатся специализированные лизосомы, называемые акросомами.
Из них наружу непосредственно перед оплодотворением выделяются ферменты, с помощью которых спермий прокладывает себе путь к яйцеклетке сквозь окружающие ее клеточные слои, переваривая отделяющий его от яйцеклетки материал.
Автолиз
Автолиз — это саморазрушение клетки, наступающее в результате высвобождения содержимого ее лизосом внутри самой клетки. Именно в связи с этим лизосомы были в свое время метко названы «орудиями самоубийства» (suicide bags). При некоторых процессах дифференцировки автолиз представляет собой нормальное явление; он может распространяться на всю ткань, как это, например, имеет место при резорбции хвоста головастика во время метаморфоза. Другой пример — превращения, которые претерпевает матка. Во время беременности матка сильно увеличивается, чтобы вместить растущий плод. После родов она постепенно приобретает свои обычные размеры в результате самопереваривания большого числа клеток. Наблюдается автолиз также и в мышцах долго остающихся без работы. После гибели клетки тоже наступает автолиз; именно поэтому пищевые продукты портятся, если они не были заморожены. Иногда автолиз является следствием некоторых лизосомных болезней.
Информация на сайте подлежит консультации лечащим врачом и не заменяет очной консультации с ним.
См. подробнее в пользовательском соглашении.
Вопрос 25. Строение лизосом.
Лизосома — (от греч. λύσις — растворяю и sōma — тело) клеточный органоид размером 0,2 — 0,4 мкм, один из видов везикул. Эти одномембранные органоиды — часть вакуома (эндомембранной системы клетки). Разные виды лизосом могут рассматриваться как отдельные клеточные компартменты. Лизосомы были впервые описаны в 1955 году Кристианом де Дювом в животной клетке, а позже были обнаружены и в растительной. У растений к лизосомам по способу образования, а отчасти и по функциям близки вакуоли. Лизосомы есть также у большинства протистов (как с фаготрофным, так и с осмотрофным типом питания) и у грибов. Таким образом, наличие лизосом характерно для клеток всех эукариот. У прокариот лизосомы отсутствуют, так как у них отсутствует фагоцитоз и нет внутриклеточного пищеварения. Один из признаков лизосом — наличие в них ряда ферментов (кислых гидролаз), способных расщеплять белки, углеводы, липиды и нуклеиновые кислоты. К числу ферментов лизосом относятся катепсины (тканевые протеазы), кислая рибонуклеаза, фосфолипаза и др. Кроме того, в лизосомах присутствуют ферменты, которые способны отщеплять от органических молекул сульфатные (сульфатазы) или фосфатные (кислая фосфатаза) группы. Для лизосом характерна кислая реакция внутренней среды. Обычно pH в лизосомах составляет около 4,5-5 (концентрация протонов на два порядка выше, чем в цитоплазме). Это обеспечивается активным транспортом протонов, который осуществляет встроенный в мембраны лизосом белок-насос протонная АТФаза. Высокая активность кислой фосфатазы ранее использовалась как один из маркеров лизосом. В настоящее время более надежным маркером считается присутствие специфических мембранных гликопротеидов — LAMP1 и LAMP2. Они присутствуют на мембране лизосом и поздних эндосом, но отсутствуют на мембранах других компартментов вакуома. Лизосомы формируются из пузырьков (везикул), отделяющихся от аппарата Гольджи, и пузырьков (эндосом), в которые попадают вещества при эндоцитозе. В образовании аутолизосом (аутофагосом) принимают участие мембраны эндоплазматического ретикулума. Все белки лизосом синтезируются на «сидячих» рибосомах на внешней стороне мембран эндоплазматического ретикулума и затем проходят через его полость и через аппарат Гольджи.Лизосомы — гетерогенные органеллы, имеющие разную форму, размеры, ультраструктурные и цитохимические особенности. «Типичные» лизосомы животных клеток обычно имеют размеры 0,1-1 мкм, сферическую или овальную форму. Число лизосом варьирует от одной (крупная вакуоль во многих клетках растений и грибов) до нескольких сотен или тысяч (в клетках животных).Общепринятой классификации и номенклатуры для разных стадий созревания и типов лизосом нет. Различают первичные и вторичные лизосомы. Первые образуются в области аппарата Гольджи, в них находятся ферменты в неактивном состоянии, вторые же содержат активные ферменты. Обычно ферменты лизосом активируются при понижении рН. Среди лизосом можно также выделить гетеролизосомы (переваривающие материал, поступающий в клетку извне — путем фаго- или пиноцитоза) и аутолизосомы (разрушающие собственные белки или органоиды клетки). Наиболее широко используется следующая классификация лизосом и связанных с ними компартментов:Ранняя эндосома — в нее поступают эндоцитозные (пиноцитозные) пузырьки. Из ранней эндосомы рецепторы, отдавшие (из-за пониженного рН) свой груз, возвращаются на наружную мембрану.Поздняя эндосома — в нее из ранней эндосомы поступают пузырьки с материалом, поглощенном при пиноцитозе, и пузырьки из аппарата Гольджи с гидролазами. Рецепторы маннозо-6-фосфата возвращаются из поздней эндосомы в аппарат Гольджи.Лизосома — в нее из поздней эндосомы поступают пузырьки со смесью гидролаз и перевариваемого материала.Фагосома — в нее попадают более крупные частицы (бактерии и т. п.), поглощенные путем фагоцитоза. Фагосомы обычно сливаются с лизосомой.Аутофагосома — окруженный двумя мембранами участок цитоплазмы, обычно включающий какие-либо органоиды и образующийся при макроаутофагии. Сливается с лизосомой.Мультивезикулярные тельца — обычно окружены одинарной мембраной, содержат внутри более мелкие окруженные одинарной мембраной пузырьки. Образуются в результате процесса, напоминающего микроаутофагию (см. ниже), но содержат материал, полученный извне. В мелких пузырьках обычно остаются и затем подвергаются деградации рецепторы наружной мембраны (например, рецепторы эпидермального фактора роста). По стадии формирования соответствуют ранней эндосоме. Описано образование мультивезикулярных телец, окруженных двумя мембранами, путем отпочковывания от ядерной оболочки.Остаточные тельца (телолизосомы) — пузырьки, содержащие непереваренный материал (в частности, липофусцин). В нормальных клетках сливаются с наружной мембраной и путем экзоцитоза покидают клетку. При старении или патологии накапливаются. Функциями лизосом являются: переваривание захваченных клеткой при эндоцитозе веществ или частиц (бактерий, других клеток). аутофагия — уничтожение ненужных клетке структур, например, во время замены старых органоидов новыми, или переваривание белков и других веществ, произведенных внутри самой клетки. автолиз — самопереваривание клетки, приводящее к ее гибели (иногда этот процесс не является патологическим, а сопровождает развитие организма или дифференцировку некоторых специализированных клеток). Пример: При превращении головастика в лягушку, лизосомы, находящиеся в клетках хвоста, переваривают его: хвост исчезает, а образовавшиеся во время этого процесса вещества всасываются и используются другими клетками тела.
Вопрос 26. Вакуоли и запасные вещества. У растений вакуоли - производные эндоплазматической сети , ограниченные мембраной - тонопластом и заполненные водянистым содержимым - клеточным соком. По- видимому, существенную роль в образовании вакуолей имеет деятельность аппарата Гольджи .В молодых делящихся растительных клетках вакуоли представляют систему канальцев и пузырьков (провакуоли), по мере роста клеток они увеличиваются, а затем сливаются в одну большую центральную вакуоль. Она занимает от 70 до 90% объема клетки, в то время как протопласт располагается в виде тонкого постенного слоя. В основном увеличение размеров клетки происходит за счет роста вакуоли. В результате этого возникает тургорное давление и поддерживается упругость клеток и тканей.Содержимое вакуоли - клеточный сок - представляет собой слабокислый (рН 2-5) водный раствор различных органических и неорганических веществ (в незрелых плодах или в зрелых плодах лимона клеточный сок имеет сильнокислую реакцию). По химическому составу и консистенции клеточный сок существенно отличается от протопласта . Эти различия связаны с избирательной проницаемостью тонопласта, выполняющего барьерную функцию. Большинство органических веществ, содержащихся в клеточном соке, относится к группе эргастических продуктов метаболизма протопласта. В зависимости от потребностей клетки они могут накапливаться в вакуоли в значительных количествах либо полностью исчезать. Наиболее обычны различные углеводы, играющие роль запасных энергетических веществ, а также органические кислоты. Вакуоли семян нередко содержат и белки-протеины. Растительные вакуоли часто служат местом концентрации разнообразных вторичных метаболитов - полифенольных соединений: флавоноидов , антоцианов , таннидов и азотсодержащих веществ - алкалоидов . В клеточном соке растворены также многие неорганические соединения.Функции вакуолей многообразны. Они формируют внутреннюю водную среду клетки, и с их помощью осуществляется регуляция водно-солевого обмена. В этом плане очень важна роль тонопласта , участвующего в активном транспорте и накоплении в вакуолях некоторых ионов.Другая важнейшая роль вакуолей состоит в поддержании тургорного гидростатического давления внутриклеточной жидкости в клетке.Наконец, третья их функция - накопление запасных веществ и "захоронение" отбросов, т.е. конечных продуктов метаболизма клетки. Иногда вакуоли разрушают токсичные или ненужные клетке вещества. Обычно это выполняется специальными небольшими вакуолями, содержащими соответствующие ферменты. Такие вакуоли получили название лизосомных .Тургорное давление в растительных клетках способствует поддержанию формы неодревесневших частей растений. Оно служит также одним из факторов роста, обеспечивая рост клеток растяжением. Потеря тургора вызывает увядание растений. Тургорное давление связано с избирательной проницаемостью тонопласта для воды и явлением осмоса. Осмос - это односторонняя диффузия воды через полупроницаемую перегородку в сторону водного раствора солей большей концентрации. Поступающая в клеточный сок вода оказывает давление на цитоплазму , а через нее - на стенку клетки, вызывая упругое ее состояние, т.е. обеспечивая тургор. Недостаток воды в растении и тем самым в отдельной клетке ведет к плазмолизу , т.е. к сокращению объема вакуоли и отделению протопластов от оболочки. Плазмолиз может быть вызван искусственно при погружении клетки в гипертонический раствор какой-либо соли или сахара. Плазмолиз обычно обратим и может служить показателем живого состояния протопласта . Вещества живого содержимого растительной клетки - протопласта и продукты его жизнедеятельности очень разнообразны. Условно их объединяют в две группы:1) конституционные, входящие в состав живой материи, и участвующие в обмене веществ (белки, нуклеиновые кислоты, липиды, углеводы и др.);2) эргастические включения (греч. эргон - работа) - представляющие собой компоненты протопласта, играющие вспомогательную роль в его жизни и являющиеся либо источниками материи и энергии при росте и работе живой клетки, либо отбросными продуктами ее метаболизма. Одни из них - запасные вещества, т.е. временно исключенные из процесса обмена веществ (белки, липиды, углеводы: крахмал, инулин сахар и др.). Другие вещества - конечные продукты, например, соли кальция.Крахмал (после целлюлозы) является самым распространенным в растительном мире углеводом. Крахмал образуется в хлоропластах во время фотосинтеза (ассимиляционный или первичный крахмал). Позже он разрушается и синтезируется в амилопластах как запасной или вторичный крахмал.Крахмальные зерна имеют разную форму (рис. 21) и образуют слоистость вокруг одной точки, называемой образовательным центром . Возникновение слоистости приписывают чередованию двух углеводов амилазы (линейные молекулы) и амилопектина (разветвленные молекулы). Расположение слоев может быть концентрическим (например, у злаков и бобовых) (рис. 21, Б; 24) и эксцентрическим (например, у картофеля) (рис. 21, А). В последнем случае, точка, вокруг которой откладываются слои, находится не в центре зерна, а сдвинута вбок.Амилопласт может содержать одно (простое зерно) или несколько крахмальных зерен (полусложное и сложное). Если в лейкопласте имеется одна точка, вокруг которой откладываются слои, то образуется простое зерно, если две и более, то образуется сложное зерно, состоящее как бы из нескольких простых. Полусложное зерно образуется в том случае, если крахмал сначала откладывается вокруг нескольких точек, а затем после соприкосновения простых зерен вокруг них возникают общие слои. Форма крахмальных зерен своеобразна у каждого вида.В клубнях георгина, земляной груши, корнях одуванчика и других растений семейства сложноцветных клеточный сок содержит близкий к крахмалу углевод инулин, отличающийся от крахмала растворимостью в воде. При действии спирта инулин кристаллизуется, образует так называемые сферокристаллы.Белки - это основные органические вещества, определяющие строение и свойства живой материи. В определенные фазы развития белки могут откладываться в запас. Запасные белки наиболее часто откладываются в виде зерен округлой или овальной формы, называемых алейроновыми. Это простые белки - протеины. Они откладываются в вакуолях или лейкопластах (алейронопласты).Запасными белками очень богаты семена бобовых и злаковых растений. Большое количество белков находится в клетках, расположенных под семенной кожурой, в так называемом алейроновом слое.Липиды включают большую группу соединений биологического происхождения. Липиды являются структурными компонентами клетки (входят в состав мембран, образуют липидные капли в цитоплазме) или эргастическими веществами. Запасные масла обычно откладываются в лейкопластах, называемых олеопластами.
Лизосома
Электронная микрофотография клетки HeLa, на которой видны компоненты эндоцитозного пути. Лизосома отмечена буквой «L», ранняя эндосома — буквой «Е», поздняя эндосома — буквой «М».
Лизосо́ма (от греч. λύσις — растворяю и sōma — тело) — окружённый мембраной клеточный органоид, в полости которого поддерживается кислая среда и находится множество растворимых гидролитических ферментов [1] . Лизосома отвечает за внутриклеточное переваривание макромолекул, в том числе при аутофагии; лизосома способна к секреции своего содержимого в процессе лизосомного экзоцитоза; также лизосома участвует в некоторых внутриклеточных сигнальных путях, связанных с метаболизмом и ростом клетки [2] . Лизосома является одним из видов везикул и относится к эндомембранной системе клетки [3] . Разные виды лизосом могут рассматриваться как отдельные клеточные компартменты.
Лизосомы были открыты в 1955 году бельгийским биохимиком Кристианом де Дювом [4] . Лизосомы есть во всех клетках млекопитающих, за исключением эритроцитов [5] . У растений к лизосомам по способу образования, а отчасти и по функциям близки вакуоли. Лизосомы есть также у большинства протистов (как с фаготрофным, так и с осмотрофным типом питания) и у грибов. Таким образом, наличие лизосом характерно для клеток всех эукариот. У прокариот лизосомы отсутствуют, так как у них отсутствует фагоцитоз и нет внутриклеточного пищеварения.
С нарушением функций лизосом связан ряд наследственных заболеваний у человека, называемых лизосомными болезнями накопления [6] .
Содержание
В 1949—1952 годах биохимик Кристиан де Дюв и его студенты, изучавшие действие инсулина в клетках печени крыс, случайно обнаружили неожиданное различие в активности кислой фосфатазы в зависимости от способа выделения. Кислая фосфатаза использовалась ими в качестве стандарта, основным предметом их изучения был фермент глюкозо-6-фосфатаза, вовлечённый в метаболизм инсулина. В ходе экспериментов выяснилось, что при фракционировании клеточного содержимого на центрифуге кислая фосфатаза была ассоциирована с микросомальной фракцией, но проявляла только десятую часть активности в сравнении с простым клеточным экстрактом, причём после нескольких дней хранения микросомальной фракции в холодильнике активность кислой фосфатазы возрастала. При обнаружении этого феномена первым объяснением было то, что произошла какая-то техническая ошибка. Однако повторение эксперимента неизменно воспроизводило первоначальную картину. Это позволило предположить существование неких окружённых мембраной клеточных частиц, которые содержат внутри себя фермент. С 1952 по 1955 год было открыто ещё несколько кислых гидролаз, связанных с микросомальной фракцией. В 1955 году, который считается годом открытия лизосом, К. де Дюв предложил название «лизосома» для клеточной органеллы, которая окружена мембраной, внутри которой поддерживается низкий pH и внутри которой находится ряд ферментов, оптимально работающих в кислой среде [7] [8] . В том же 1955 году американский цитолог Алекс Новиков (англ.) русск. из Вермонтского университета США , блестяще владевший техникой микроскопии, посетил лабораторию К. де Дюве и смог получить первые электронные фотографии этих органелл, используя препарат частично очищенных лизосом. Позднее в 1961 году Алекс Новиков с помощью гистохимического выявления кислой фосфатазы и электронной микроскопии подтвердил локализацию этого фермента в лизосомах [9] [10] . В 1963 году бельгийский биохимик Генри Хэрс, ранее работавший в группе К. де Дюве, обнаружил недостаточность лизосомного фермента α-глюкозидазы у пациентов с болезнью Помпе и высказал предположение о связи других генетических заболеваний с нарушением работы лизосом [11] . В настоящее время более 50 наследственных заболеваний связывают с лизосомной недостаточностью [12] .
В 1974 году за свой вклад в раскрытие структурной и функциональной организации клетки К. де Дюв был удостоен Нобелевской премией по медицине [13] .
Лизосомы являются гетерогенными по форме, размеру, ультраструктурным и цитохимическим особенностям. В клетках животных размер лизосом составляет обычно менее 1 мкм, хотя в некоторых типах клеток, например, в макрофагах, размер лизосом может превышать несколько микрон. Лизосомы, как правило, имеют сферическую, овальную, иногда тубулярную форму [14] . Число лизосом варьирует от одной (крупная вакуоль во многих клетках растений и грибов) до нескольких сотен или тысяч (в клетках животных). Лизосомы у животных обычно составляют не более 5 % внутриклеточного объёма [15] .
Один из признаков лизосом — наличие в них ряда ферментов (кислых гидролаз), способных расщеплять белки, углеводы, липиды и нуклеиновые кислоты. К числу ферментов лизосом относятся катепсины (тканевые протеазы), кислая рибонуклеаза, фосфолипаза и др. Кроме того, в лизосомах присутствуют ферменты, которые способны отщеплять от органических молекул сульфатные (сульфатазы) или фосфатные (кислая фосфатаза) группы. Всего полость лизосомы содержит около 60 растворимых кислых гидролитических ферментов [2] .
Для лизосом характерна кислая реакция внутренней среды, которая обеспечивает оптимум работы лизосомных гидролаз [14] . Обычно pH в лизосомах составляет около 4,5-5, то есть концентрация протонов в них на два порядка выше, чем в цитоплазме. Это обеспечивается активным транспортом протонов, который осуществляет встроенный в мембраны лизосом белок-насос протонная АТФаза [15] . Помимо протонного насоса в мембрану лизосом встроены белки-переносчики для транспорта в цитоплазму продуктов гидролиза макромолекул: аминокислот, сахаров, нуклеотидов, липидов [16] .
Высокая активность кислой фосфатазы ранее использовалась как один из маркеров лизосом. В настоящее время более надежным маркером считается присутствие специфических мембранных гликопротеидов — LAMP1 и LAMP2. Они присутствуют на мембране лизосом и поздних эндосом, но отсутствуют на мембранах других компартментов вакуома.
Лизосомы формируются из пузырьков (везикул), отделяющихся от аппарата Гольджи, и пузырьков (эндосом), в которые попадают вещества при эндоцитозе [17] . В образовании аутолизосом (аутофагосом) принимают участие мембраны эндоплазматического ретикулума. Все белки лизосом синтезируются на «сидячих» рибосомах на внешней стороне мембран эндоплазматического ретикулума и затем проходят через его полость и через аппарат Гольджи.
Общепринятой классификации и номенклатуры для разных стадий созревания и типов лизосом нет. Различают первичные и вторичные лизосомы. Первые образуются в области аппарата Гольджи, в них находятся ферменты в неактивном состоянии, вторые же содержат активные ферменты. Обычно ферменты лизосом активируются при понижении рН. Среди лизосом можно также выделить гетеролизосомы (переваривающие материал, поступающий в клетку извне — путём фаго- или пиноцитоза) и аутолизосомы (разрушающие собственные белки или органоиды клетки). Наиболее широко используется следующая классификация лизосом и связанных с ними компартментов:
- Ранняя эндосома — в неё поступают эндоцитозные (пиноцитозные) пузырьки. Из ранней эндосомы рецепторы, отдавшие (из-за пониженного рН) свой груз, возвращаются на наружную мембрану.
- Поздняя эндосома — в неё из ранней эндосомы поступают пузырьки с материалом, поглощённом при пиноцитозе, и пузырьки из аппарата Гольджи с гидролазами. Рецепторы маннозо-6-фосфата возвращаются из поздней эндосомы в аппарат Гольджи.
- Лизосома — в неё из поздней эндосомы поступают пузырьки со смесью гидролаз и перевариваемого материала.
- Фагосома — в неё попадают более крупные частицы (бактерии и т. п.), поглощённые путём фагоцитоза. Фагосомы обычно сливаются с лизосомой.
- Аутофагосома — окружённый двумя мембранами участок цитоплазмы, обычно включающий какие-либо органоиды и образующийся при макроаутофагии. Сливается с лизосомой.
- Мультивезикулярные тельца — обычно окружены одинарной мембраной, содержат внутри более мелкие окружённые одинарной мембраной пузырьки. Образуются в результате процесса, напоминающего микроаутофагию (см. ниже), но содержат материал, полученный извне. В мелких пузырьках обычно остаются и затем подвергаются деградации рецепторы наружной мембраны (например, рецепторы эпидермального фактора роста). По стадии формирования соответствуют ранней эндосоме. Описано образование мультивезикулярных телец, окруженных двумя мембранами, путём отпочковывания от ядерной оболочки.
- Остаточные тельца (телолизосомы) — пузырьки, содержащие непереваренный материал (в частности, липофусцин). В нормальных клетках сливаются с наружной мембраной и путем экзоцитоза покидают клетку. При старении или патологии накапливаются.
Функциями лизосом являются:
- переваривание захваченных клеткой при эндоцитозе веществ или частиц (бактерий, других клеток)
- аутофагия — уничтожение ненужных клетке структур, к примеру, во время замены старых органоидов новыми, или переваривание белков и других веществ, произведенных внутри самой клетки
- автолиз — самопереваривание клетки, приводящее к её гибели (иногда этот процесс не является патологическим, а сопровождает развитие организма или дифференцировку некоторых специализированных клеток). Пример: При превращении головастика в лягушку, лизосомы, находящиеся в клетках хвоста, переваривают его: хвост исчезает, а образовавшиеся во время этого процесса вещества всасываются и используются другими клетками тела.
- растворение внешних структур (см, например, остеокласты)
У многих протистов и у животных, имеющих внутриклеточное пищеварение, лизосомы участвуют в переваривании пищи, захваченной путем эндоцитоза. При этом лизосомы сливаются с пищеварительными вакуолями. У протистов непереваренные остатки пищи обычно удаляются из клетки при слиянии пищеварительной вакуоли с наружной мембраной.
Многие клетки животных, у которых преобладает полостное пищеварение (например, хордовые) получают питательные вещества из межклеточной жидкости или плазмы крови с помощью пиноцитоза. Эти вещества также вовлекаются в обмен веществ клетки после их переваривания в лизосомах. Хорошо изученный пример такого участия лизосом в обмене веществ — получение клетками холестерина. Холестерин, приносимый кровью в виде ЛПНП, поступает внутрь пиноцитозных везикул после соединения ЛПНП с рецепторами ЛПНП на мембране. Рецепторы возвращаются к мембране из ранней эндосомы, а ЛПНП поступают в лизосомы. После этого ЛПНП перевариваются, а высвободившийся холестерин через мембрану лизосом поступает в цитоплазму.
Косвенно лизосомы участвуют в обмене, обеспечивая десенсибилизацию клеток к воздействию гормонов. При длительном действии гормона на клетку часть рецепторов, связавших гормон, поступают в эндосомы и затем деградируют внутри лизосом. Снижение числа рецепторов понижает чувствительность клетки к гормону.
Для крупных вакуолей растений характерна запасающая функция — в них могут накапливаться ионы, пигменты (например, антоцианы), вторичные метаболиты, белки (в алейроновых зернах эндосперма злаков). Внутри вакуолей (например, в прорастающих семенах) у растений происходят и процессы переваривания запасенных белков.
Обычно различают два типа аутофагии — микроаутофагия и макроаутофагия. При микроаутофагии, как при образовании мультивезикулярных телец, образуются впячивания мембраны эндосомы или лизосомы, которые затем отделяются в виде внутренних пузырьков, только в них попадают вещества, синтезированные в самой клетке. Таким путём клетка может переваривать белки при нехватке энергии или строительного материала (например, при голодании). Но процессы микроаутофагии происходят и при нормальных условиях и в целом неизбирательны. Иногда в ходе микроаутофагии перевариваются и органоиды; так, у дрожжей описана микроаутофагия пероксисом и частичная микроаутофагия ядер, при которой клетка сохраняет жизнеспособность.
При макроаутофагии участок цитоплазмы (часто содержащий какие-либо органоиды) окружается мембранным компартментом, похожим на цистерну эндоплазматической сети. В результате этот участок оказывается отгорожен от остальной цитоплазмы двумя мембранами. Затем такая аутофагосома сливается с лизосомой, и её содержимое переваривается. Видимо, макроаутофагия также неизбирательна, хотя часто подчеркивается, что с помощью неё клетка может избавляться от «отслуживших свой срок» органоидов (митохондрий, рибосом и др.).
Третий тип аутофагии — шаперон-зависимая. При этом способе происходит направленный транспорт частично денатурировавших белков из цитоплазмы сквозь мембрану лизосомы в её полость.
Ферменты лизосом нередко высвобождаются при разрушении мембраны лизосомы. Обычно при этом они инактивируются в нейтральной среде цитоплазмы. Однако при одновременном разрушении всех лизосом клетки может произойти её саморазрушение — автолиз. Различают патологический и обычный автолиз. Распространенный вариант патологического автолиза — посмертный автолиз тканей.
В норме процессы автолиза сопровождают многие явления, связанные с развитием организма и дифференцировкой клеток. Так, автолиз клеток описывается как механизм разрушения тканей у личинок насекомых при полном превращении, а также при рассасывании хвоста у головастика. Правда, эти описания относятся к периоду, когда различия между апоптозом и некрозом ещё не были установлены, и в каждом случае требуется выяснять, не лежит ли на самом деле в основе деградации органа или ткани апоптоз, не связанный с автолизом.
У растений автолизом сопровождается дифференциация клеток, которые функционируют после смерти (например, трахеид или члеников сосудов). Частичный автолиз происходит и при созревании клеток флоэмы- члеников ситовидных трубок.
Иногда из-за неправильной работы лизосом развиваются болезни накопления, при которых ферменты из-за мутаций не работают или работают плохо. Примером лизосомных болезней накопления может служить болезнь Гоше, болезнь Помпе, Болезнь Тея — Сакса. Всего известно более 50 наследственных заболеваний, связанных с нарушением функции лизосомы [12] .
Лизосомы. Этапы эндоцитоза. Переваривание при эндоцитозе.
Организация клетки. Лизосомы
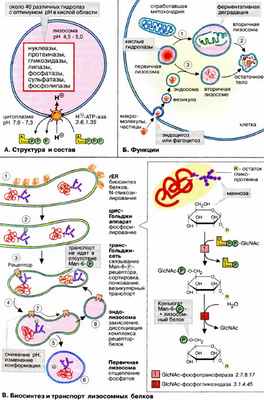
А. Структура и состав
Лизосомы — это органеллы диаметром 0,2-2,0 мкм, окруженные простой мембраной, способные принимать самые разные формы. Обычно на клетку приходится несколько сотен лизосом. Функция лизосом заключается в деградации клеточных компонентов. Деградация достигается за счет присутствия в лизосомах около 40 типов различных расщепляющих ферментов — гидролаз с оптимумом действия в кислой области. Главный фермент лизосом — кислая фосфатаза. В мембране лизосом находятся АТФ-зависимые протонные насосы вакуольного типа. Они обогащают лизосомы протонами, вследствие чего для внутренней среды лизосом рН 4,5-5,0 (в то время как в цитоплазме рН 7,0-7,3). Лизосомные ферменты имеют оптимум рН около 5,0, т. е. в кислой области. При рН, близких к нейтральным, характерным для цитоплазмы, эти ферменты обладают низкой активностью. Очевидно, это служит механизмом защиты клеток от самопереваривания о том случае, если лизосомный фермент случайно попадет в цитоплазму.
Главная функция лизосом — ферментативная деградация попавших в них макромолекул и органелл. Примером может служить деградация отработавших митохондрий по механизму аутофагии (захвата органеллы) ( 1 ). После захвата органеллы первичные лизосомы превращаются во вторичные, в которых и идет процесс гидролитического расщепления ( 2 ). В итоге образуются «остаточные тела» , состоящие из негидролизовавшихся фрагментов. Лизосомы ответственны также за деградацию макромолекул и частиц, захваченных клетками путем эндоцитоза и фагоцитоза, например липопротеинов, протеогормонов и бактерий (гетерофагия). В этом случае лизосомы сливаются с эндосомами ( 3 ), содержащими вещества, подлежащие деградации.
В. Биосинтез и транспорт лизосомных белков
Первичные лизосомы образуются в аппарате Гольджи.
Лизосомные белки синтезируются в ШЭР, где они гликозилируются путем переноса олигосахаридных остатков ( 1 , см. с. 226). На последующей стадии, типичной для лизосомных белков, терминальные маннозные остатки (Man) фосфорилируются no C-6 (на схеме справа). Реакция протекает в две стадии. Сначала на белок переносится GlcNAc-фосфат, а затем идет отщепление GlcNAc. Таким образом, лизосомные белки в процессе сортировки приобретают концевой остаток маннозо-6-фосфата (Man-6-P, 2 ).
В мембранах аппарата Гольджи имеются молекулы-рецепторы, специфичные для Man-6-P-остатков и за счет этого специфически узнающие и селективно связывающие лизосомные белки ( 3 ). Локальное накопление этих белков происходит с помощью клатрина. Этот белок позволяет вырезать и транспортировать подходящие мембранные фрагменты в составе транспортных везикул к эндолизосомам ( 4 ), которые затем созревают с образованием первичных лизосом ( 5 ) в заключение от Man-6-P отщепляется фосфатная группа ( 6 ).
Man-6-P-рецепторы используются вторично в процессе рецикла. Снижение рН а эндолизосомах приводит к диссоциации белков от рецепторов ( 7 ). Затем рецепторы с помощью транспортных везикул переносятся обратно в аппарат Гольджи ( 8 ).
Некоторые редко встречающиеся заболевания связаны с генетическими дефектами лизосомных ферментов, так как эти ферменты участвуют в деградации гликогена (гликогенозы), липидов (липидозы) и протеогликанов (мукополисахаридозы). Продукты, которые не могут участвовать в метаболизме из-за дефектов или отсутствия соответствующих ферментов, накапливаются в остаточных телах, что приводит к необратимому повреждению клеток и как результат к нарушению функций соответствующих органов.
Читайте также: