Механизмы формирования митотического веретена деления
Добавил пользователь Валентин П. Обновлено: 30.01.2026
• Хромосомы разделяются посредством митотического веретена
• Веретено представляет собой симметричную биполярную структуру, состоящую из микротрубочек, расположенных между двумя полюсами. На каждом полюсе находится центросома
• Центросомы прикрепляются к веретену за счет взаимодействия своих кинетохоров с микротрубочками
Веретено представляет собой сложную динамическую структуру, которая быстро образуется в начале процесса деления и при его окончании так же быстро разрушается. Веретено необходимо для митоза и служит для выполнения двух отдельных функций: (1) обеспечение разделения реплицированных хромосом по дочерним ядрам при делении ядра (кариокинез) и (2) управление процессом деления цитоплазмы (цитокинез).
Если заблокировать образование веретена (например, обработав клетки различными химическими соединениями), то хромосомы конденсируются, но не движутся, как это обычно происходит в митозе, и процесс деления останавливается. Во многом веретено представляет собой род биологического мотора, который превращает химическую энергию в механическую работу, необходимую для перемещения хромосом и деления клетки. Функции веретена отражаются в его строении. Симметричная структура с двумя полюсами необходима для успешного прохождения митоза.
Действительно, она отражает принцип парности клеточного деления, при котором одна клетка и реплицированная ДНК делятся на две отдельных дочерних клетки.
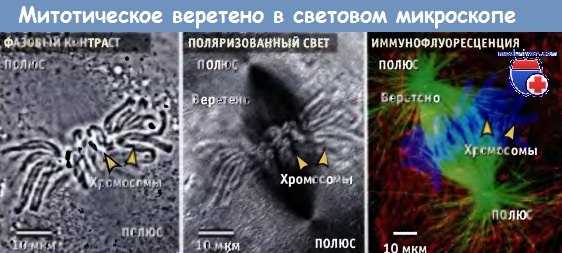
Метафазное веретено в живой клетке тритона, видимое в фазовоконтрастном и поляризационном микроскопе.
Показана часть такой же клетки с веретеном в той же ориентации после иммунофлуоресцентного окрашивания микротрубочек (зеленым), хромосом (синим) и кератиновых филаментов (красным).
Отметим, что веретено не видно в фазовом контрасте, но видно в поляризованном свете. Микротрубочки веретена наиболее отчетливо видны после иммунофлуоресцентного окрашивания.
Веретено можно увидеть с помощью различных методов. Основной структурный элемент веретена — микротрубочки, слишком малы для того, чтобы их можно было видеть с помощью светового микроскопа (т. е. из-за недостаточного разрешения). Поэтому, хотя в клетках высших организмов часто можно наблюдать с помощью обычного светового микроскопа конденсированные хромосомы, веретена при этом не видно. Однако во многих случаях можно сделать вывод о наличии веретена по косвенному признаку, поскольку эта структура вытесняет видимые клеточные органеллы. При этом, как показано на рисунке ниже, пространство, занимаемое веретеном, по сравнению с окружающей цитоплазмой кажется более прозрачным. Хотя вначале исследователи считали, что веретено состоит из волокон, до начала 1950-х гг. это предположение не было подтверждено прямыми наблюдениями.
К этому времени усовершенствования техники поляризационной световой микроскопии позволили увидеть веретено на препаратах клеток. Типичная фотография веретена, полученная с помощью светового микроскопа, представлена на рисунке ниже. Структура приобрела черную окраску из-за взаимодействия поляризованного света с микротрубочками. В течение 1970-х гг. была разработана техника с использованием флуоресцентных зондов, которая позволила наблюдать компоненты веретена в трехмерном пространстве даже в живой клетке. Эта техника позволяет прослеживать положения одного или нескольких специфических белков веретена в ходе митоза. Одним из таких белков почти всегда является тубулин, поскольку он обеспечивает визуализацию микротрубочек.
При наблюдении в электронном микроскопе веретено типичной клетки млекопитающих состоит из трех структурных компонентов. Как показано на рисунке ниже, в каждом из двух полярных участков находится центросома. Эта красивая органелла включает пару небольших, интенсивно окрашенных структур, называемых центриолями.
Они окружены более или менее плотным диффузным материалом. Между центросомами расположены хромосомы, которые в большинстве клеток являются самыми крупными структурами веретена. Хромосомы состоят из компактных, плотно скрученных и сильно базофильных фибрилл хроматина диаметром 25 нм. Каждая хромосома содержит две небольших структуры, называемых кинетохоры (от греч. kineto — подвижный; chora — пространство). Кинетохоры прикрепляются к противоположным сторонам своей центромеры. Между полюсами веретена проходит плотный пучок расположенных параллельно друг другу микротрубочек.
На рисунке ниже это видно наиболее отчетливо. Один из концов микротрубочек веретена обычно расположен на самом полюсе или рядом с ним. Другой находится в области веретена в свободном состоянии или связан с кинетохором. Микротрубочки растут от каждого полюса, что делает веретено симметричной структурой, образованной двумя параллельными и перекрывающимися пучками микротрубочек. Каждый из этих пучков называется полуверетено. У большинства позвоночных полуверетено состоит из 600-750 микротрубочек, 30-40% которых заканчиваются на кинетохорах.
В каждом полуверетене, наряду с основными микротрубочками, из каждого полюса выходят другие микротрубочки. Эти микротрубочки распространяются во всех направлениях, образуя радиальные структуры, которые называются звездами (астерами) и располагаются в центре каждого полюса. Так же как и микротрубочки веретена, все астральные микротрубочки одним концом ориентированы на полюс, а другим на отдаленную точку в цитоплазме. В митозе астральные звезды обладают несколькими функциями. Наряду с позиционированием веретена в клетке, которое определяет плоскость цитокинеза, они также участвуют в отделении полюсов (центросом) при образовании веретена в анафазе В.
Критическую роль в митозе также играют два кинетохора каждой хромосомы. Их роль в перемещении хромосом обнаружилась очень давно, поскольку оказалось, что фрагменты хромосом, не содержащие кинетохора, не способны к направленному движению. Особенно важно, каким образом располагаются кинетохоры по отношению друг к другу. Поскольку они расположены на противоположных сторонах центромеры, то обращены к противоположным полюсам веретена, что позволяет реплицированным хромосомам присоединяться к обоим полюсам. Наличие такой позиционной взаимосвязи между двумя кинетохорами существенно для сегрегации двух хроматид в разные ядра. При образовании веретена каждый кинетохор связывается с концами множества микротрубочек, исходящих из одного полюса, и образует пучок, называемый кинетохорным пучком, который проходит между кинетохором и полюсом.
Нити кинетохора и сами кинетохоры не являются всего лишь транспортной системой канатов, позволяющих хроматидам продвигаться к полюсам. Скорее всего, они играют более важную активную роль, не только определяя направление движения хромосомы, но и генерируя усилия, необходимые для этого движения.
Для того чтобы понять молекулярные механизмы митоза, необходимо ответить на следующие кардинальные вопросы. Каким образом формируется веретено и как обеспечивается его биполярная структура? Каким образом генерируются усилия, обеспечивающие движение хромосом и как это движение регулируется? Как обеспечивается точность процесса сегрегации хромосом? Каким образом после сегрегации хромосом происходит разделение цитоплазмы с образованием двух дочерних клеток?
Электронная микрофотография, демонстрирующая основные структурные элементы митотического веретена.
Крупные пучки микротрубочек соединяют каждую центросому с кинетохорами на хромосомах.
В центре фотографии кинетохоры, помеченные стрелками, иллюстрируют, что два кинетохора на хромосоме обращены к противоположным полюсам веретена. На основной фотографии помещено изображение центросомы в электронном микросокопе.
Две центриоли расположены под прямым углом друг к другу, так, что одна выглядит как круг, а другая как прямоугольник.
Вокруг первой центриоли находится скопление гранулярного материала
(сравните область, примыкающую к центриоли, с более удаленными частями цитоплазмы,
которые окрашены менее интенсивно и где заметно присутствие многих мембранных везикул). Нити кинетохора, прикрепленные к сестринским хроматидам.
Окрашивание иммунофлуоресцентным методом (слева) и фотография, сделанная в электронном микроскопе (в центре и справа). Последовательность событий мейоза включает два клеточных деления.
При первом делении происходит разделение гомологичных хромосом,
при втором разделяются индивидуальные хроматиды (каждой хромосомы).
При митозе происходит только разделение хроматид.
Редактор: Искандер Милевски. Дата обновления публикации: 18.3.2021
Митотическое веретено: структура, формирование, функции и эволюция
Видео: Митотическое веретено: структура, формирование, функции и эволюция
Содержание:
В митотическое веретено или ахроматический, также называемый митотическим механизмом, представляет собой клеточную структуру, состоящую из микротрубочек белковой природы, которые образуются во время деления клеток (митоз и мейоз).
Термин ахроматический означает, что он не окрашивается красителями орсеина А или В. Веретено участвует в справедливом распределении генетического материала между двумя дочерними клетками в результате деления клеток.
Деление клеток - это процесс, при котором из зиготы образуются как гаметы, которые являются мейотическими клетками, так и соматические клетки, необходимые для роста и развития организма.
Переход между двумя последовательными делениями составляет клеточный цикл, продолжительность которого широко варьируется в зависимости от типа клетки и стимулов, которым она подвергается.
Во время митоза эукариотической клетки (клетки, которая имеет истинное ядро и мембранно-ограниченные органеллы) происходит несколько фаз: S-фаза, профаза, прометафаза, метафаза, анафаза, телофаза и интерфейс.
Хромосомы первоначально конденсируются, образуя две идентичные нити, называемые хроматидами. Каждая хроматида содержит одну из двух ранее созданных молекул ДНК, связанных вместе областью, называемой центромерой, которая играет фундаментальную роль в процессе миграции к полюсам до деления клетки.
Митотическое деление происходит на протяжении всей жизни организма. Подсчитано, что в течение жизни человека в организме происходит около 10 17 деления клеток. Мейотическое деление происходит в клетках, производящих гаметы, или половых клетках.
Структура и формирование
Связь с цитоскелетом
Ахроматическое веретено считается продольной системой белковых микрофибрилл или клеточных микротрубочек. Он образуется во время деления клеток между центромерами хромосом и центросомами на полюсах клеток и связан с миграцией хромосом с образованием дочерних клеток с таким же количеством генетической информации.
Центросома - это область, в которой микротрубочки берут начало как от ахроматического веретена, так и от цитоскелета. Эти микротрубочки веретена состоят из димеров тубулина, заимствованных из цитоскелета.
В начале митоза сеть микротрубочек цитоскелета клетки расчленяется и образуется ахроматическое веретено. После деления клетки веретено расчленяется, и сеть микротрубочек цитоскелета реорганизуется, возвращая клетку в состояние покоя.
Важно различать три типа микротрубочек в митотическом аппарате: два типа микротрубочек веретена (кинетохоры и полярные микротрубочки) и один тип микротрубочек звездочки (астральные микротрубочки).
Двусторонняя симметрия ахроматического веретена обусловлена взаимодействиями, которые удерживают его две половинки вместе. Эти взаимодействия бывают: либо латеральными, между перекрывающимися положительными концами полярных микротрубочек; или они являются терминальными взаимодействиями между микротрубочками кинетохоры и кинетохор сестринских хроматид.
Клеточный цикл и ахроматическое веретено: S-фаза, профаза, прометафаза, метафаза, анафаза, телофаза и интерфаза.
Репликация ДНК происходит во время S-фазы клеточного цикла, затем во время профазы центросомы мигрируют к противоположным полюсам клетки, а хромосомы также конденсируются.
Прометафаза
В прометафазе происходит формирование митотического аппарата благодаря сборке микротрубочек и их проникновению внутрь ядра. Сестринские хроматиды, связанные центромерами, образуются, а они, в свою очередь, связываются с микротрубочками.
Метафаза
Во время метафазы хромосомы выравниваются в экваториальной плоскости клетки. Веретено состоит из центрального митотического веретена и пары звездочек.
Каждая звездочка состоит из микротрубочек, расположенных в форме звезды, которые простираются от центросом в кору клетки. Эти астральные микротрубочки не взаимодействуют с хромосомами.
Затем говорится, что астра излучается от центросомы в кору клетки и участвует как в расположении всего митотического аппарата, так и в определении плоскости деления клеток во время цитокинеза.
Анафаза
Позже, во время анафазы, микротрубочки ахроматического веретена закрепляются на положительном конце хромосом через их кинетохоры, а на отрицательном конце - к центросоме.
Происходит разделение сестринских хроматид на независимые хромосомы. Каждая хромосома, прикрепленная к микротрубочке кинетохоры, движется к полюсу клетки. Одновременно происходит разделение полюсов ячеек.
Телофаза и цитокинез
Наконец, во время телофазы и цитокинеза ядерные мембраны образуются вокруг дочерних ядер, и хромосомы теряют свой уплотненный вид.
Митотическое веретено исчезает по мере деполимеризации микротрубочек и клеточного деления, выходящего на поверхность раздела.
Механизм хромосомной миграции
Однако механизм, вовлеченный в миграцию хромосом к полюсам и последующее отделение полюсов друг от друга, точно не известен; Известно, что в этом процессе участвуют взаимодействия кинетохоры и прикрепленной к ней микротрубочки веретена.
По мере того как каждая хромосома мигрирует к соответствующему полюсу, происходит деполимеризация прикрепленной микротрубочки или кинетохорной микротрубочки. Считается, что эта деполимеризация может вызывать пассивное движение хромосомы, прикрепленной к микротрубочке веретена.
Считается также, что с кинетохорой могут быть связаны другие моторные белки, в которых будет использоваться энергия гидролиза АТФ.
Эта энергия будет способствовать миграции хромосомы вдоль микротрубочки к ее концу, называемому «меньше», где расположена центросома.
В унисон может происходить деполимеризация конца микротрубочки, который связывается с кинетохорой, или «плюсового» конца, что также будет способствовать перемещению хромосомы.
Функция
Ахроматическое или митотическое веретено - это клеточная структура, которая выполняет функцию закрепления хромосом через их кинетохоры, выравнивания их по экватору клетки и, наконец, направления миграции хроматид к противоположным полюсам клетки перед их делением, обеспечивая распределение выравнивание генетического материала между двумя образующимися дочерними клетками.
Если в этом процессе возникают ошибки, возникает недостаток или избыток хромосом, что приводит к аномальным паттернам развития (возникающим во время эмбриогенеза) и различным патологиям (возникающим после рождения человека).
Другие функции для проверки
Есть данные, что микротрубочки ахроматического веретена участвуют в определении местоположения структур, ответственных за деление цитоплазмы.
Основное свидетельство состоит в том, что разделение клеток всегда происходит по средней линии веретена, где полярные волокна перекрываются.
Эволюция механизма
Эволюционно он был выбран как механизм с высокой степенью избыточности, в котором каждый шаг выполняется моторными белками микротрубочек.
Считается, что эволюционное приобретение микротрубочек произошло из-за процесса эндосимбиоза, при котором эукариотическая клетка абсорбировала из окружающей среды прокариотическую клетку, которая проявляла эти ахроматические структуры веретена. Все это могло произойти до начала митоза.
Эта гипотеза предполагает, что белковые структуры микротрубочек могли первоначально выполнять функцию движения. Затем, когда они стали частью нового организма, микротрубочки составили цитоскелет, а позже - митотический аппарат.
В истории эволюции были вариации базовой схемы деления эукариотических клеток. Клеточное деление представляет собой лишь некоторые фазы клеточного цикла, который является важным процессом.
Механизмы формирования митотического веретена деления
• Изменения в центросомах и в цитоплазме в начале митоза приводят к образованию вокруг каждой центросомы астральных структур, состоящих из коротких высокодинамичных микротрубочек
• Образование митотического веретена инициируется взаимодействием между астральными структурами, образованными двумя центросомами
• Разделение центросом зависит от белковых моторов микротрубочек
• Путь образования веретена зависит оттого, произошло расхождение центросом до или после разрыва оболочки ядра
По мере продвижения клетки из интерфазы в митоз, структура микротрубочек проходит через ряд быстрых и глубоких изменений. В цитоплазме происходит разборка длинных микротрубочек, типичных для интерфазной клетки, и каждая из двух центросом нуклеирует плотную радиальную структуру, состоящую из коротких микротрубочек (звезду, или звездчатую структуру).
Как показано на рисунке ниже, в конце концов две эти звездчатые структуры своими микротрубочками участвуют в образовании веретена. В начале митоза происходят изменения количества и распределения микротрубочек, которое определяется разными процессами, в том числе изменениями центросомы.
Незадолго до того как клетка коммитируется к митозу, меняются обе центросомы. Они становятся способны к нуклеации гораздо большего числа микротрубочек, чем в течение интерфазы. Как только это происходит, связанные с центросомой белки становятся более фосфорилированными, в центросомах возрастает содержание у-тубулина, и происходит расширение области перицентриолярного материала. Остается неясным, каким образом происходит процесс «созревания». Вероятно, в нем участвуют специфические киназы, которые активируются по мере перехода клетки из фазы G2 в митоз, включая главную регуляторную киназу митоза.
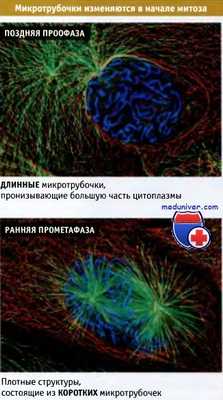
Для превращения одной структуры в другую требуется всего несколько минут.
Микротрубочки флуоресцируют зеленым цветом, хромосомы синим, а промежуточные филаменты — красным.
Примерно в то же время из-за изменения условий в цитоплазме происходит дестабилизация микротрубочек. Это приводит к замещению сети, состоящей из длинных интерфазных микротрубочек, двумя звездчатыми структурами, состоящими из коротких микротрубочек и образующимися на центросомах, как описано выше. В результате, к началу прометафазы общая длина микротрубочек в клетке уменьшается, и скорость, с которой образуются новые и разрушаются старые микротрубочки (т. е. оборот микротрубочек), увеличивается.
Это означает, что, как только начинается разрушение ядерной оболочки, область, окружающая ядро, постоянно зондируется большим количеством крайне динамичных микротрубочек, растущих от каждой звездчатой структуры. Как мы увидим далее, столь динамичный характер обеспечивает связывание звездчатых структур с хромосомами.
По мере расхождения центросом, в результате взаимодействия между окружающими их звездчатыми структурами начинается образование веретена. Это в высшей степени гибкий процесс, который осуществляется по одному из двух путей, в зависимости от того, разделились центросомы до или после разрушения ядерной оболочки. Оба пути включают взаимодействие микротрубочек с белковыми моторами. В случаях, когда ядерная оболочка разрушается до начала расхождения центросом, высвободившиеся хромосомы распределяются по цитоплазме, и возникает только одна большая звездообразная структура.
Как показано на рисунке ниже, образуется «однополюсное» полуверетено; оно сохраняется до тех пор, пока нс разойдутся две центросомы, после чего образуется двухполюсное веретено. Расхождение центросом после разрушения оболочки ядра включает эффект двух сил: одна их толкает за счет ки-незиноподобного белка Eg5, который взаимодействует с примыкающими микротрубочками противоположной полярности, расположенными между двумя центросомами. Вторая сила тянет центросомы за счет цитоплазматического динеина, закрепленного на периферии клетки (т. е. в ее кортексе).
В отсутствие препятствий эти две силы раздвигают центросомы до тех пор, пока не разойдутся две звездчатые структуры. Однако расхождение центросом ограничивается существованием других моторов, которые связаны со звездчатыми структурами, а также образованием нитей кинетохора на сестринских кинетохорах.
Веретено образуется другим способом, если две звездчатые структуры уже разошлись и произошло разрушение ядерной оболочки. При этом расхождение звездчатых структур происходит без участия Eg5, который оказывается недоступным, поскольку находится в ядре. Вместо него с микротрубочками, отходящими из каждой центросомы, взаимодействует цитоплазматический динеин, и когда укрепилась связь между двумя центросомами, он способен обеспечить их расхождение. В этом случае динеин локализован в клеточном кортексе и на поверхности ядерной оболочки. Актиновые филаменты определяют направление, по которому расходятся две центросомы, взаимодействуя с миозином, расположенным или в самих центросомах или вдоль микротрубочек
Когда расхождение двух звездчатых структур происходит до разрушения ядерной оболочки, то часто в области, где перекрываются микротрубочки, образуется первичное веретено. Однако до разрушения ядерной оболочки эта структура является неустойчивой, и микротрубочки центросом могут разойтись настолько далеко, что не будут перекрываться. Это объясняется тем, что для устойчивости веретена необходимы белки, которые находятся в интерфазном ядре, и выходят в цитоплазму только после разрушения оболочки ядра. В результате во многих клетках, находящихся в поздней профазе, две центросомы и звездчатые структуры расположены в противоположных участках ядра и не взаимодействуют между собой.
В таких клетках веретено образуется лишь после того, как кинетохоры станут доступными для микротрубочек и смогут связать две звездчатых структуры.
Кадр видеофильма, иллюстрирующий изменение организации микротрубочек в митотические структуры. Два механизма образования веретена различаются на этапе расхождения центросом по отношению к процессу разрушения ядерной оболочки.
Способность клетки к образованию веретена, независимо от того, когда расходятся центросомы, подчеркивает необычайно гибкий характер процесса его формирования. Слева представлено монополярное веретено, образующееся в клетке кенгуровой крысы,
при блокировании процесса расхождения центросом (вид сбоку).
Хромосомы (флуоресцируют оранжевым цветом) присоединены к одному из полюсов.
Видны толстые нити кинетохоров. Для сравнения на врезке показано обычное биполярное веретено.
Справа показано аналогичное монополярное веретено в клетке человека (вид спереди).
В центре находятся центросомы (флуоресцируют синим цветом). Процесс образования веретена начинается, как только произошел разрыв оболочки ядра.
Вначале обе центросомы примыкают друг к другу. Белки Eg5 и HSET представляют собой моторы микротрубочек и входят в семейство кинезинов.
Однако Eg5 движется к плюс-концу микротрубочки, a HSET — к ее минус-концу.
Длина веретена определяется балансом усилий, развиваемых тремя моторами.
Митотический цикл. Митоз
1. Клеточный цикл. Митоз, как механизм бесполого размножения. Фазы митоза. Биологическое значение митоза.
2. Мейоз как цитологическая основа развития гамет. Фазы и стадии мейотических делений. Биологическое значение мейоза.
3. Гаметогенез – образование половых клеток.
4. Размножение. Биологическое значение размножения. Виды размножения.
Нерегулярный тип полового размножения: партеногенез, апомиксис, гиногенез, андрогенез.
Клеточный цикл. Митоз, как механизм бесполого размножения. Фазы митоза. Биологическое значение митоза.
Материальной преемственностью между поколениями организмов, размножающихся половым путем, является клетка, как универсальная единица структурно-функциональной организации живой материи. Материальные носители наследственности находятся в ядре, хотя небольшая часть наследственного материала находится в цитоплазме, т.е. ядро является основным хранителем наследственной информации. В 80 года 19 века В. Флемингом, Э Страсбургером в ядре были обнаружены нитевидные структуры, которые в 1888г. В. Вальдейер назвал хромосомами, в переводе в греческого – окрашенное тело. В дальнейшем было подтверждено, что именно хромосомы являются носителями наследственной информации, которые наиболее четко видны во время деления клеток.
Хромосомы (от греч.хрома – цвет и сома - тело) – интенсивно окрашивающиеся нитевидные или палочковидные структуры, четко выявляемые в клетке во время митоза. Они состоят из ядерного хроматина, главным компонентом которого служат ДНК. В связи с этим хромосомы являются материальным субстратом наследственности клеток. Число х. в ядрах всех клеток данного вида постоянно. Каждая ч. в хромосомном наборе имеет определенный генный состав и может передать только ей присущую наследственную информацию.
Хроматиды (от греч.хрома – цвет и идос - подобный) – продольные половинки хромосом, состоящие в свою очередь из хромонем. Х. в качестве составной части хромосом выступают в период профазы и метофазы мейоза. Позднее во время анафазы после расщепления хромосом на хроматиды каждая из хроматид становится самостоятельным образованием и обозначается уже как дочерняя или сестринская хромосома.
Хроматин (от греч.хрома - цвет) – сильно окрашивающееся основными красителями вещество клеточного ядра. В химическом отношении х. является сложным белком, состоящим из ДНК и гистонов.
Промежуток времени между окончанием одного клеточного деления – митоза и окончанием последующего называется клеточным циклом или митотическим циклом.
Митотический цикл. Митоз
Митоз — основной способ деления эукариотических клеток, при котором сначала происходит удвоение, а затем равномерное распределение между дочерними клетками наследственного материала.
Митоз представляет собой непрерывный процесс, в котором выделяют четыре фазы: профазу, метафазу, анафазу и телофазу. Перед митозом происходит подготовка клетки к делению, или интерфаза. Период подготовки клетки к митозу и собственно митоз вместе составляют митотический цикл. Ниже приводится краткая характеристика фаз цикла.
Интерфаза состоит из трех периодов: пресинтетического, или постмитотического, — G1, синтетического — S, постсинтетического, или премитотического, — G2.
Пресинтетический период (2n 2c, где n — число хромосом, с — число молекул ДНК) — рост клетки, активизация процессов биологического синтеза, подготовка к следующему периоду.
Синтетический период (2n 4c) — репликация ДНК.
Постсинтетический период (2n 4c) — подготовка клетки к митозу, синтез и накопление белков и энергии для предстоящего деления, увеличение количества органоидов, удвоение центриолей.
Профаза (2n 4c) — демонтаж ядерных мембран, расхождение центриолей к разным полюсам клетки, формирование нитей веретена деления, «исчезновение» ядрышек, конденсация двухроматидных хромосом.
Метафаза (2n 4c) — выстраивание максимально конденсированных двухроматидных хромосом в экваториальной плоскости клетки (метафазная пластинка), прикрепление нитей веретена деления одним концом к центриолям, другим — к центромерам хромосом.
Анафаза (4n 4c) — деление двухроматидных хромосом на хроматиды и расхождение этих сестринских хроматид к противоположным полюсам клетки (при этом хроматиды становятся самостоятельными однохроматидными хромосомами).
Телофаза (2n 2c в каждой дочерней клетке) — деконденсация хромосом, образование вокруг каждой группы хромосом ядерных мембран, распад нитей веретена деления, появление ядрышка, деление цитоплазмы (цитотомия). Цитотомия в животных клетках происходит за счет борозды деления, в растительных клетках — за счет клеточной пластинки.
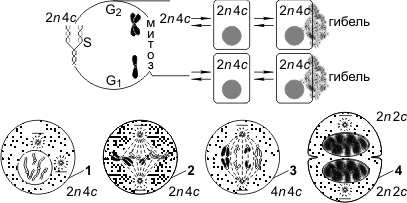
Митотический цикл, митоз: 1 — профаза; 2 — метафаза; 3 — анафаза; 4 — телофаза.
Биологическое значение митоза. Образовавшиеся в результате этого способа деления дочерние клетки являются генетически идентичными материнской. Митоз обеспечивает постоянство хромосомного набора в ряду поколений клеток. Лежит в основе таких процессов, как рост, регенерация, бесполое размножение и др.
Амитоз — прямое деление интерфазного ядра путем перетяжки без образования хромосом, вне митотического цикла. Описан для стареющих, патологически измененных и обреченных на гибель клеток. После амитоза клетка не способна вернуться в нормальный митотический цикл.
• В отсутствие хромосом цитастеры расходятся полностью и не могут образовать веретена
• Хромосомы стабилизируют геометрию веретена и входящие в него микротрубочки путем связывания микротрубочек цитастеров со своими кинетохорами
• Веретено может образоваться в отсутствие центросом, хотя при этом оно образуется медленнее и не содержит астральных микротрубочек
• Образование центросомального веретена включает процесс нуклеации микротрубочек хромосомами, а также функционирование нескольких различных типов моторных белков микротрубочек
По мере образования веретено стабилизируется хромосомами, их кинетохорами и моторными белками, которые связываются с микротрубочками и доставляют необходимые компоненты веретена.
Для образования основных форм веретена особенно важны белковые моторы, которые связывают соседние микротрубочки противоположной полярности. Такие микротрубочки находятся там, где перекрываются две звездчатые структуры, и при образовании сшивок, даже в отсутствие поблизости хромосом, создается веретенообразная структура по длине близкая к обычному веретену. Однако в отсутствие хромосом веретено неустойчиво и постепенно теряет микротрубочки.
Каким образом связывание хромосом с образующимся веретеном предотвращает потерю микротрубочек? Ответить на этот вопрос непросто, и, вероятно, в процесс вовлекаются несколько разных механизмов. Каждый из кинетохоров хромосомы организует микротрубочки звездчатых структур в специальный пучок, называемый кинетохорным пучком, который соединяет кинетохор с полюсом. Включение микротрубочек в состав волокна кинетохора ноги пучка увеличивает их устойчивость по сравнению с другими микротрубочками веретена.
Это характерно для значительной части микротрубочек в каждой астральной структуре; обычно в метафазе около 30-40% из 1200— 1500 микротрубочек в веретене стабилизируются, связываясь с кинетохором. Поскольку каждая хромосома содержит два кинетохора, образование кинетохорного пучка связывает два полюса веретена вместе, что приводит к еще большему взаимодействию между микротрубочками астральных структур.
Пока происходит организация кинетохорного пучка, каждая астральная структура также отбирает и накапливает разнообразные белки, способствующие стабилизации микротрубочек. Некоторые из них являются структурными белками и организованы в рыхлый матрикс веретена, окружающий микротрубочки. Например, когда из ядра выходит белок ядерного митотического аппарата (NuMA) и фосфорилируется CDK1, он накапливается в веретене.
Этот белок присоединяется к различным белковым моторам веретена, тем самым связывая микротрубочки и стабилизируя их. Стабилизирующая роль компонентов матрикса существенна, однако не настолько, как кинетохоров. В зрелом веретене продолжительность существования микротрубочек, связанных с кинетохором, в 10 раз превышает таковую для других микротрубочек веретена.
Интересно, что биполярное веретено может образовываться в отсутствие центросом. Это происходит в процессе самоорганизации, при котором случайно нуклеированные микротрубочки собираются в двухполюсную структуру с участием хромосом и белковых моторов микротрубочек. Такой «ацентросомальный» путь сборки веретена характерен для всех высших растений, а также для мейоза, происходящего на ранних стадиях развития некоторых животных. Возможно, что он представляет собой эволюционно более древний предшественник образования веретена с участием центросом и что он маскируется присутствием центросом.
В пользу такой точки зрения говорит тот факт, что даже клетки млекопитающих, обычно содержащие центросомы, при их отсутствии могут формировать двухполюсное веретено. Этот факт прекрасно иллюстрирует способность клеток вырабатывать множество различных механизмов для выполнения одной и той же задачи.
При ацентросомальной сборке веретена поблизости от каждой хромосомы образуются короткие микротрубочки. В этом процессе принимают участие белки, расположенные на поверхности хромосом. Эти микротрубочки вначале ориентируются случайно, а затем под действием моторных белков организуются в параллельные пучки. Главную роль в процессе играют моторы, которые сразу связываются с двумя противоположно ориентированными микротрубочками и одновременно движутся в направлении плюс-конца каждой из них. Вначале эти белки ориентируют микротрубочки поблизости от хромосомы, просто располагаясь по их длине и образуя сшивки.
Затем, двигаясь к плюс-концу каждой микротрубочки, моторы сортируют их на две группы параллельных поляризованных микротрубочек, с совпадающими плюс-концами. Белковый мотор может оставаться там в связанном состоянии, тем самым поддерживая структуру. В этом процессе сборки веретена также участвуют белковые моторы, находящиеся на плечах хромосом и называемые хромокинезинами. Отчасти их роль заключается в том, чтобы связать микротрубочки, находящиеся поблизости от хромосом, с тем чтобы они по мере образования веретена оставались в его центре.
После сортировки микротрубочки в пределах каждой из структур связываются вместе своими минус-концами с помощью моторов, движущихся к этим концам (например, цитоплазматического динеина и HSET), тем самым придавая всей конструкции форму веретена. Процесс самосборки происходит вблизи каждой хромосомы и носит независимый характер. Образующиеся при этом многочисленные веретена затем сливаются в одно большое веретено с широкими полюсами. Структурные белки и другие компоненты матрикса (например, NuMA), которые транспортируются к минус-концам микротрубочек, т. е. к формирующемуся полюсу, скрепляют структуру и стабилизируют область полюсов.
Если центросомы не являются необходимыми компонентами образования двухполюсного веретена, то почему тогда они присутствуют на полюсах у большинства клеток млекопитающих в митозе? Одна из причин может заключаться в том, что центросомы более выгодны с кинетической точки зрения и обеспечивают более быстрое образование веретена. Это важно, поскольку при развитии организма часто веретено должно формироваться синхронно и очень быстро. Другая причина в том, что полюса веретена, образованные центросомами, содержат цитастеры, которые отсутствуют у полюсов веретена, образующегося по ацентросомальному пути.
При позиционировании веретена в клетке эти структуры определяют положение борозды деления при цитокинезе. Эта сторона функционирования астральных структур имеет критическое значение в последующем развитии организма. Наконец, наряду со своей ролью центра организации микротрубочек, центросома выполняет в клетке и другие функции (например, участвует в образовании первичных ресничек и в продвижении клетки по циклу). Ассоциация центросом на каждом из полюсов веретена служит удобным и надежным механизмом, гарантирующим, что каждая новая клетка унаследует копию этой важной органеллы.
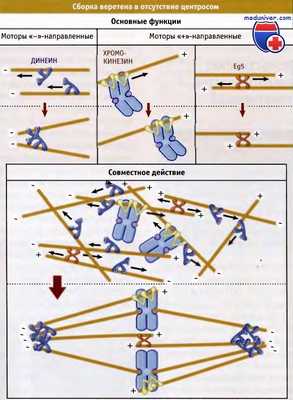
Процесс нуклеации микротрубочек вокруг хромосом происходит случайным образом.
Когда микротрубочки формировались, три типа белковых моторов принимают совместное участие в их организации в биполярные структуры, расположенные вокруг хромосом.
Читайте также: