Тогавирусы. Репродукция тогавирусов. Ретровирусы. Репродукция ретровирусов.
Добавил пользователь Morpheus Обновлено: 29.01.2026
Вирусы относят к царству Vira. Это мельчайшие микроорганизмы, не имеющие клеточного строения, белоксинтезирующей системы, содержащие только один тип нуклеиновой кислоты (ДНК или РНК).
Они отличаются особым разобщенным (дисъюнктив- ным) способом размножения (репродукции): в клетке отдельно синтезируются нуклеиновые кислоты вирусов и их белки и затем происходит их сборка в вирусные частицы. Вирусы, являясь облигатными внутриклеточными паразитами, размножаются в цитоплазме иди ядре клетки. Сформированная вирусная частица называется вирионом (рис. 2.9).
Морфологию и структуру вирусов изучают с помощью электронного микроскопа, так как их размеры малы и сравнимы с толщиной оболочки бактерий. Форма вирионов может быть различной (рис. 2.10): палочковидной (вирус табачной мозаики), пу- левидной (вирус бешенства), сферической (вирусы полиомиелита, ВИЧ), в виде сперматозоида (многие бактериофаги).
Размеры вирусов определяют с помощью электронной микроскопии, методом ультрафильтрации через фильтры с известным диаметром пор, методом ультрацентрифугирования. Одним из самых мелких вирусов является вирус полиомиелита (около 20 нм), наиболее крупным — натуральной оспы (около 350 нм).
Вирусы имеют уникальный геном, так как содержат либо ДНК, либо РНК. Поэтому различают ДНК-содержащие и РНК- содержащие вирусы (табл. 2.1). Они обычно гаплоидны, т.е. имеют один набор генов. Геном вирусов представлен различными видами нуклеиновых кислот: двунитчатыми, однонитчатыми, линейными, кольцевыми, фрагментированными. Среди РНК-
Капсид, состоящий из капсомеров
Рис.2.9. Строение вириона (капсомерного). Суперкап- сид у некоторых вирусов отсутствует (схема).
Рис.2.10. Формы вирионов.
1 — вирус оспы; 2 — вирус герпеса; 3 — аденовирус; 4 — паповавирус; 5 — гепаднавирус; 6 — парамиксовирус; 7 — вирус гриппа; 8 — коронавирус; 9 — аренавирус; 10 — ретровирус; 11 — реовирус; 12 — пикорнавирус; 13 — вирус бешенства; 14 — тогавирус, флавивирус; 15 — буньявирус.
Классификация и некоторые свойства основных вирусов Семейство вирусов Наличие супер- капсида Размер вириона Типовые представители Дї [К-содержащие вирусы 70-90 45-50 200
Вирус гепатита В
Вирусы простого герпеса, цитоме-
галии и др. Вирусы папилломы, полиомы Аденоассоциированный вирус Вирусы осповакцины, натураль-ной оспы РНК-содержащие вирусы Аренавирусы + 50- -300 Вирусы Ласса, Мачупо Буньявирусы + 90- -100 Вирусы лихорадки с почечным синдромом Калицивирусы — 20- -30 Калицивирусы человека Коронавирусы + 80- -130 Коронавирусы человека Оргомиксовирусы + 80- -120 Вирусы гриппа Парамиксовирусы + 150- -300 Вирусы кори, паротита, парагриппа, респираторно-синци- тиальный Пикорнавирусы — 20- -30 Вирусы полиомиелита, ECHO, Коксаки, гепатита А Рабдовирусы + 70- -175 Вирус бешенства Реовирусы — 60- -80 Реовирусы, ротавирусы Ретровирусы + 80- -100 Вирусы лейкоза человека, иммунодефицита человека Тогавирусы + 30- -90 Вирусы Синдбис, лошадиных эн цефалитов Флавивирусы + 30- -90 Вирус клещевого и японского энцефалитов, денге, желтой лихорадки, краснухи
содержащих вирусов различают вирусы с положительным (плюс- нить РНК) геномом. Плюс-нить РНК этих вирусов выполняет наследственную функцию и функцию информационной РНК (иРНК). Имеются также РНК-содержащие вирусы с отрицательным (минус-нить РНК) геномом. Минус-нить РНК этих вирусов выполняет только наследственную функцию.
Геном вирусов способен включаться в состав генетического аппарата клетки в виде провируса, проявляя себя генетическим паразитом клетки. Нуклеиновые кислоты некоторых вирусов (ви-
русы герпеса и др.) могут находиться в цитоплазме инфицированных клеток, напоминая плазмиды.
Различают просто устроенные (например, вирус полиомиелита) и сложно устроенные (например, вирусы гриппа, кори) вирусы. У просто устроенных вирусов нуклеиновая кислота связана с белковой оболочкой, называемой капсидом (от лат.
capsa — футляр). Капсид состоит из повторяющихся морфологических субъединиц — капсомеров. Нуклеиновая кислота и капсид, вза-имодействуя друг с другом, образуют нуклеокапсид. У сложно устроенных вирусов капсид окружен дополнительной липопро- теидной оболочкой — суперкапсидом (производное мембранных структур клетки-хозяина), имеющей «шипы». Для вирионов характерен спиральный, кубический и сложный тип симметрии капсида. Спиральный тип симметрии обусловлен винтообразной структурой нуклеокапсида, кубический тип симметрии — образованием изометрически полого тела из капсида, содержащего вирусную нуклеиновую кислоту.
Капсид и суперкапсид защищают вирионы от влияния окру-жающей среды, обусловливают избирательное взаимодействие (адсорбцию) с клетками, определяют антигенные и иммуноген- ные свойства вирионов. Внутренние структуры вирусов называются сердцевиной.
В вирусологии используют следующие таксономические категории: семейство (название оканчивается на viridae), подсемейство (название оканчивается на virinae), род (название оканчивается на virus).
Однако названия родов и особенно подсемейств сформулированы не для всех вирусов. Вид вируса биноминального названия, как у бактерий, не получил.
В основу классификации вирусов положены следующие категории:
тип нуклеиновой кислоты (ДНК или РНК), ее структура, количество нитей (одна или две), особенности воспроизводства вирусного генома;
размер и морфология вирионов, количество капсомеров и тип симметрии;
чувствительность к эфиру и дезоксихолату;
место размножения в клетке;
антигенные свойства и пр.
Вирусы поражают позвоночных и беспозвоночных животных, а также растения и бактерии. Являясь основными возбудителями инфекционных заболеваний человека, вирусы также участвуют в процессах канцерогенеза, могут передаваться различными путями, в том числе через плаценту (вирус краснухи, цитомега- ловирус и др.), поражая плод человека. Они могут приводить к постинфекционным осложнениям — развитию миокардитов, панкреатитов, иммунодефицитов и др.
Кроме обычных вирусов, известны и так называемые неканонические вирусы — прионы — белковые инфекционные частицы, являющиеся агентами белковой природы, имеющие вид фибрилл размером 10—20x100—200 нм. Прионы, по-видимому, являются одновременно индукторами и продуктами автономного гена человека или животного и вызывают у них энцефалопатии в условиях медленной вирусной инфекции (болезни Крейтц- фельдта—Якоба, куру и др.).
Другими необычными агентами, близкими к вирусам, являются вироиды — небольшие молекулы кольцевой, суперспи- рализованной РНК, не содержащие белка, вызывающие заболевания у растений.
Тогавирусы. Репродукция тогавирусов. Ретровирусы. Репродукция ретровирусов.
Тогавирусы. Репродукция тогавирусов. Ретровирусы. Репродукция ретровирусов.
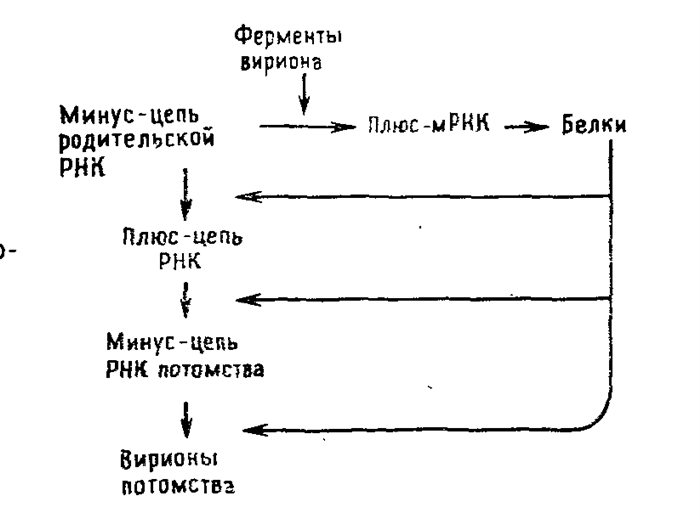
После проникновения вириона тогавируса в клетку синтезируется вирусная полимераза (РНК-зависимая РНК-полимераза), катализирующая образование -РНК на матрице +РНК. Образовавшеся цепи -РНК служат матрицей для синтеза двух типов +РНК (полная и короткая нити). Каждая +РНК транслируется в большой полипептид, подвергающийся последовательному расщепленния и процессингу. Полная нить (42 S) служит шаблоном для синтеза вирусных полипетидов или идёт на построение геномов дочерних популяций вируса; короткая нить (26 S) кодирует белки капсида и два оболочечных белка.
Последние гликолизируются, расщепляются и встраиваются в клеточную мембрану, модифицируя ее. Дочерние вирионы тогавируса почкуются через изменённые участки мембраны с образованием суперкапсида.
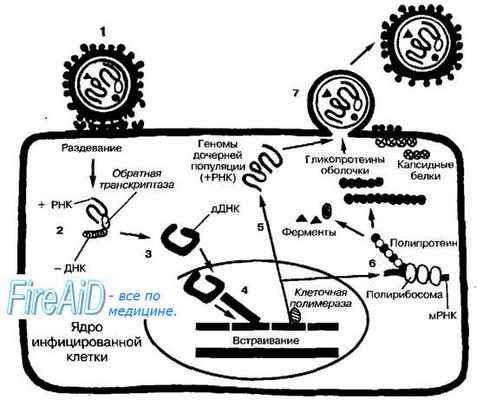
Рис. 5-2. Репродуктивный цикл ретровирусов. Инфекционная вирусная частица (1) проникает в клетку путём слияния с клеточной стенкой после адсорбции на ней. Затем вирус «раздевается», а обратная транскриптаза индуцирует синтез -ДНК, используя в качестве матрицы молекулу +РНК (2) +ДНК копируется с синтезированной молекулы -ДНК, в результате чего образуется двойная цепь ДНК (дДНК) (3). дДНК транспортируется в ядро клетки, где клеточная ДНК подвергается ДНК-сплайсингу (связывание одной молекулы ДНК с другой) с образованием рекомбинантов с вирусной ДНК (4). Интегрированная молекула ДНК транскрибируется клеточной ДНК-зависимой РНК-полимеразой с образованием вирусной +РНК (5). Часть молекул +РНК используется для создания геномов дочерних популяций, другая часть подвергается РНК-сплайсингу
Ретровирусы. Репродукция ретровирусов
Репродуктивный цикл +РНК-ретровирусов уникален, так как при его реализации как промежуточный продукт образуются молекулы ДНК (рис. 5-2). Поскольку обратная транскрипция и интеграция вирусного генома предшествуют репликации, то (в противоположность прочих молекулам вирусных +РНК) плюс-молекулы РНК ретровирусов не проявляют инфекционных свойств, а служат матрицей для синтеза молекулы -ДНК РНК-зависимой ДНК-полимеразой (обратной транскриптазой, ревертазой), входящей в состав вирусной частицы. С молекул -ДНК копируются цепи +ДНК. Затем обе цепи соединяются с образованием двойной цепи и транспортируются в ядро клетки. Клеточные эндонуклеазы и лигазы (интегразы) катализируют сплайсинг ДНК клетки с вирусной ДНК. В результате образуются новые, рекомбинантные молекулы ДНК. Вновь образованная ДНК транскрибируется клеточной ДНК-зависимой РНК-полимеразой в +РНК.
Полная копия +РНК способна формировать геномы дочерних популяций, а также транслироваться как мРНК с образованием структурных белков и ферментов, участвующих в сборке вирионов. Эти копии могут быть подвергнуты РНК-сплайсингу — устранению интронов из мРНК-предшественника; в результате образуются малые молекулы мРНК, кодирующие поверхностные, регуляторные и добавочные белки вируса. Самосборка вирусов и созревание вирусных частиц происходит в цитоплазме. Зрелые вирионы выходят из клетки, отделяясь от неё почкованием (высвобождение вируса в окружающую среду может длиться годами).
Информация на сайте подлежит консультации лечащим врачом и не заменяет очной консультации с ним.
См. подробнее в пользовательском соглашении.
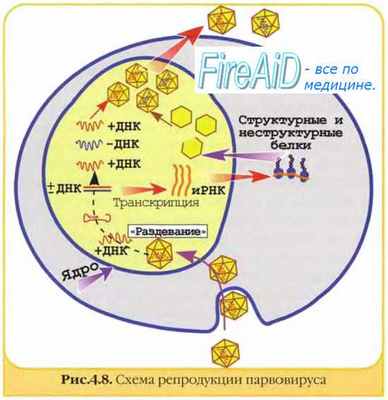
Репродукция ДНК-вирусов. Репликативный цикл ДНК-содержащих вирусов. Репродукция паповавирусов. Репродукция аде
Оглавление темы "Вирусология. Репродукция вирусов. Генетика вирусов.":
1. Вирусология. История вирусологии. Шамберлан. Ру. Пастер. Ивановский.
2. Репродукция вирусов. Репродукция +РНК-вирусов. Пикорнавирусы. Репродукция пикорнавирусов.
3. Тогавирусы. Репродукция тогавирусов. Ретровирусы. Репродукция ретровирусов.
4. Репродукция -РНК-вирусов. Репродукция вирусов с двухнитевыми РНК.
5. Репродукция ДНК-вирусов. Репликативный цикл ДНК-содержащих вирусов. Репродукция паповавирусов. Репродукция аденовирусов.
6. Репродукция герпесвирусов. Репликативный цикл герпесвирусов. Поксвирусы. Репродукция поксвирусов.
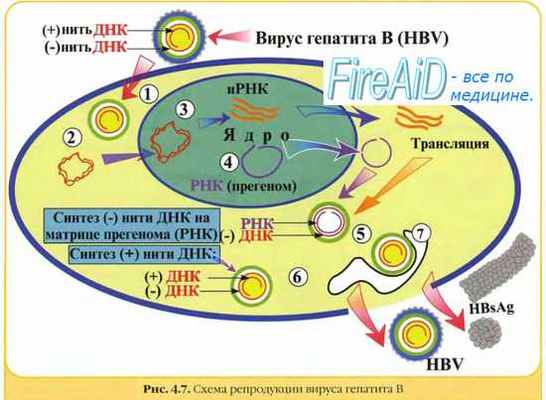
7. Репродукция вируса гепатита В. Репликативный цикл вируса гепатита В.
8. Генетика вирусов. Характеристика вирусных популяций. Генофонд вирусных популяций.
9. Мутации вирусов. Спонтанные мутации вирусов. Индуцированные мутации вирусов. Проявление мутаций вирусов в фенотипе.
10. Генетические взаимодействия между вирусами. Рекомбинации и перераспределение генов вирусами. Обмен фрагментами генома вирусами. Антигенный шифт.
Репродукция ДНК-вирусов. Репликативный цикл ДНК-содержащих вирусов. Репродукция паповавирусов. Репродукция аденовирусов.
Вирусы, лишённые суперкапсида (например, аденовирусы) проникают в клетки путём виропексиса, а имеющие таковой (покс- и герпесвирусы) — за счёт слияния суперкапсида с клеточной мембраной. Репродуктивный цикл ДНК-содержащих вирусов включает раннюю и позднюю стадии (рис. 5-4). У крупных ДНК-вирусов имеется явное несоответствие между кодирующе ёмкостью генома и молекулярной массой вирусиндуцированных белков и белков, входящих состав вирионов. Например, у герпесвирусов лишь 15% ДНК кодирует все белки вирионов и их предшественников. Возможно, значительная часть генома содержит гены, кодирующие синтез ферментов и регуляторных белков. Папова-, адено- и герпесвирусы репродуцируются относительно однотипно, в то время как репродукция поксвирусов имеет некоторые особенности.
Ранняя стадия репродукции. Вирусная ДНК проникает в ядро клетки, где транскрибируется клеточной ДНК-зависимой РНК-полимеразой. При этом считываетеся, а затем транслируется часть вирусного генома («paнние гены»). В результате синтезируются «ранние белки» (регуляторные и матричные белки вирусные полимеразы).
Регуляторные белки выполняют различные функции. При заражении клетки они блокируют синтез клеточных РНК, ДНК и белка и одновременно способствуют экспрессии вирусного генома, изменяя специфичность реагирования клеточных полимераз и полирибосом. Они так же запускают репликацию клеточной ДНК, модифицированной встроенными геномами ДНК содержащих вирусов и ретровирусов, то есть репликацию вирусных геномов. Вирусспецифические полимеразы. В репликацию вирусных геномов также вовлечены вирусоспецифические ДНК-полимеразы, участвующие в образовании молекул ДНК дочерних популяций.
Матричные белки необходимы для репликации нуклеиновых кислот и сборки дочерних популяций. Они образуют электронно-плотные скопления в клетке, известные как тельца включений (например, тельца Гварнери при натуральной оспе).
Поздняя стадия репродукции. На этом этапе происходит синтез нуклеиновых кислот вируса. Не вся вновь синтезированная вирусная ДНК упаковывается в вирионы дочерней популяции. Часть ДНК («поздние гены») используется для синтеза «поздних белков», необходимых для сборки вирионов. Их образование катализируют вирусные и модифицированные клеточные полимеразы.
Паповавирусы и аденовирусы. Репродукция паповавирусов. Репродукция аденовирусов.
Адсорбция, проникновение и депротеинизация аналогичны таковым у РНК-содержащих вирусов, но у папова- и аденовирусов депротеинизация протекает в ядре, а у РНК-вирусов — в цитоплазме.
Ранняя фаза репродукции. Вирусная ДНК («ранние гены») транскрибируется в ядре клетки. На одной из нитей ДНК реализуется транскрипция вирусной «ранней» мРНК. Механизмы транскрипции вирусной ДНК аналогичны считыванию информации с клеточной ДНК. Специфическая мРНК транслируется, начинается синтез ферментов, необходимых для образования дочерних копий ДНК. Синтез клеточной ДНК может временно усиливаться, но затем обязательно подавляется регуляторными белками вируса.
Поздняя фаза репродукции. В течение поздней фазы дочерняя вирусная ДНК продолжает активно транскрибироваться клеточными РНК-полимеразами, в результате чего появляются продукты поздних вирусспецифических синтезов. «Поздняя» мРНК мигрирует в цитоплазму и транслируется на рибосомах. В результате синтезируются капсидные белки дочерней популяции, которые транспортируются в ядро и собираются вокруг молекул дочерней ДНК новых вирусных частиц. Выход полных дочерних популяций сопровождается гибелью клетки.
- Читать далее "Репродукция герпесвирусов. Репликативный цикл герпесвирусов. Поксвирусы. Репродукция поксвирусов."
Вирусология_конспект лекций. Конспект лекций
ГЛАВА 4. РЕПРОДУКЦИЯ ВИРУСОВ
Процесс репродукции вирусов может быть условно разделен на две фазы. Первая фаза охватывает события, которые ведут к адсорбции и проникновению вируса в клетку, освобождению его внутреннего компонента и модификации его таким образом, что он способен вызвать инфекцию. Соответственно, первая фаза включает в себя три стадии: 1) адсорбция вируса на клетках; 2) проникновение вируса в клетки; 3) раздевание вируса в клетке. Эти стадии направлены на то, чтобы вирус был доставлен в соответствующие клеточные структуры, и его внутренний компонент был освобожден от защитных оболочек. Как только эта цель достигнута, начинается вторая фаза репродукции, в течение которой происходит экспрессия вирусного генома. Эта фаза включает в себя стадии: 1) транскрипции, 2) трансляции информационных РНК, 3) репликации генома, 4) сборки вирусных компонентов. Заключительной стадией репродукции является выход вируса из клетки.
ПЕРВАЯ ФАЗА РЕПРОДУКЦИИ
АДСОРБЦИЯ ВИРУСА НА КЛЕТКАХ
Взаимодействие вируса с клеткой начинается с процесса адсорбции, т. е. прикрепления вирусных частиц к клеточной поверхности. Процесс адсорбции возможен при наличии соответствующих рецепторов на поверхности клетки и «узнающих» их субстанций на поверхности вируса. Самые начальные процессы адсорбции имеют неспецифический характер, и в основе их может лежать электростатическое взаимодействие положительно и отрицательно заряженных группировок на поверхности вируса и клетки. Однако узнавание клеточных рецепторов вирусными белками, ведущее к прикреплению вирусной частицы к клетке, является высоко специфическим процессом. Белки на поверхности вируса, узнающие специфические группировки на плазматической мембране клетки и обусловливающие прикрепление к ним вирусной частицы, называются прикрепительными белками.
Вирусы используют рецепторы, предназначенные для прохождения в клетку необходимых для ее жизнедеятельности веществ: питательных веществ, гормонов, факторов роста и т. д. Рецепторы могут иметь разную химическую природу и представлять собой белки, углеводный компонент белков и липидов, липиды. Рецепторами для вирусов гриппа и парамиксовирусов является сиаловая кислота в составе гликопротеидов и гликолипидов (ганглиозидов), для рабдовирусов и реовирусов — также углеводный компонент в составе белков и липидов, для пикорнавирусов и аденовирусов — белки, для некоторых вирусов — липиды. Специфические рецепторы играют роль не только в прикреплении вирусной частицы к клеточной поверхности. Они определяют дальнейшую судьбу вирусной частицы, ее внутриклеточный транспорт и доставку в определенные участки цитоплазмы и ядра, где вирус способен инициировать инфекционный процесс. Вирус может прикрепиться и к неспецифическим рецепторам и даже проникнуть в клетку, однако только прикрепление к специфическому рецептору приведет к возникновению инфекции.
Прикрепление вирусной частицы к клеточной поверхности вначале происходит путем образования единичной связи вирусной частицы с рецептором. Однако такое прикрепление непрочно, и вирусная частица может легко оторваться от клеточной поверхности — обратимая адсорбция (рис. 7, а). Для того чтобы наступила необратимая адсорбция, должны появиться множественные связи между вирусной частицей и многими молекулами рецепторов, т. е. должно произойти стабильное мультивалентное прикрепление. Количество молекул клеточных рецепторов в участках адсорбции может доходить до 3000. Стабильное связывание вирусной частицы с клеточной поверхностью в результате мультивалентного прикрепления происходит благодаря возможности свободного перемещения молекул рецепторов в липидном бислое плазматической мембраны, которое определяется подвижностью, «текучестью» белково-липидного слоя. Увеличение текучести липидов является одним из наиболее ранних событий при взаимодействии вируса с клеткой, следствием которого является формирование рецепторных полей в месте контакта вируса с клеточной поверхностью и стабильное прикрепление вирусной частицы к возникшим группировкам — необрат
имая адсорбция (рис. 7, б).
Рис. 7. Адсорбция вируса осповакцины на клетке (электронно-микроскопическое изображение): а – обратимая; б – необратимая.
Количество специфических рецепторов на поверхности клетки колеблется между 10 4 и 10 5 на одну клетку. Рецепторы ряда вирусов могут быть представлены лишь в ограниченном наборе клеток-хозяев, и этим может определяться чувствительность организма к данному вирусу. Например, пикорнавирусы адсорбируются только на клетках приматов. Рецепторы для других вирусов, напротив, широко представлены на поверхности клеток различных видов, как, например, рецепторы для ортомиксовирусов и парамиксовирусов, представляющие собой сиалил-содержащие соединения. Поэтому эти вирусы имеют относительно широкий диапазон клеток, на которых может происходить адсорбция вирусных частиц. Рецепторами для ряда тогавирусов обладают клетки исключительно широкого круга хозяев: эти вирусы могут адсорбироваться и инфицировать клетки как позвоночных, так и беспозвоночных.
Наличие специфических рецепторов на поверхности клетки в ряде случаев обусловливает феномен зависимого от хозяина ограничения, т. е. способность вируса заражать лишь определенные виды животных. В целом ограничения при взаимодействии рецепторных систем вируса и клетки биологически оправданы и целесообразны, хотя в ряде случаев они являются «перестраховкой». Так, многие линии клеток, устойчивых к вирусам полиомиелита и Коксаки, можно заразить депротеинизированными препаратами РНК, выделенными из этих вирусов. Такое заражение клеток идет в обход естественных входных путей инфекции через взаимодействие с клеточными рецепторами. Известна потенциальная способность вирусов животных реплицироваться в протопластах дрожжей, грибов и бактерий, а бактериофагов — в клетках животных. Таким образом, вирусные ДНК и РНК обладают способностью заражать и более широкий круг хозяев, чем вирусы.
ПРОНИКНОВЕНИЕ ВИРУСОВ В КЛЕТКУ
Исторически сложилось представление о двух альтернативных механизмах проникновения в клетку вирусов животных — путем виропексиса (эндоцитоза) и путем слияния вирусной и клеточной мембран. Однако оба эти механизма не исключают, а дополняют друг друга.
Термин «виропексис», означает, что вирусная частица попадает в цитоплазму в результате инвагинации участка плазматической мембраны и образования вакуоли, которая содержит вирусную частицу.
Рецепторный эндоцитоз. Виропексис представляет собой частный случай рецепторного или адсорбционного эндоцитоза. Этот процесс является обычным механизмом, благодаря которому в клетку поступают питательные и регуляторные белки, гормоны, липопротеины и другие вещества из внеклеточной жидкости. Рецепторный эндоцитоз происходит в специализированных участках плазматической мембраны, где имеются специальные ямки, покрытые со стороны цитоплазмы особым белком с большой молекулярной массой — клатрином. На дне ямки располагаются специфические рецепторы. Ямки обеспечивают быструю инвагинацию и образование покрытых клатрином внутриклеточных вакуолей. Полупериод проникновения вещества внутрь клетки по этому механизму не превышает 10 мин с момента адсорбции. Количество образующихся в одну минуту вакуолей достигает более 2000. Таким Образом, рецепторный эндоцитоз представляет собой хорошо слаженный механизм, который обеспечивает быстрое проникновение в клетку чужеродных веществ.
Покрытые вакуоли сливаются с другими, более крупными цитоплазматическими вакуолями, образуя рецептосомы, содержащие рецепторы, но не содержащие клатрин, а те в свою очередь сливаются с лизосомами. Таким путем проникшие в клетку белки обычно транспортируются в лизосомы, где происходит их распад на аминокислоты; они могут и миновать лизосомы, и накапливаться в других участках клетки в недеградированной форме. Альтернативой рецепторного эндоцитоза является жидкостный эндоцитоз, когда инвагинация происходит не в специализированных участках мембраны. Большинство оболочечных и безоболочечных вирусов животных проникает в клетку по механизму рецепторного эндоцитоза. Эндоцитоз обеспечивает внутриклеточный транспорт вирусной частицы в составе эндоцитарной вакуоли, поскольку вакуоль может двигаться в любом направлении и сливаться с клеточными мембранами (включая ядерную мембрану), освобождая вирусную частицу в соответствующих внутриклеточных участках. Таким путем, например, ядерные вирусы попадают в ядро, а реовирусы — в лизосомы. Однако проникшие в клетку вирусные частицы находятся в составе вакуоли и отделены от цитоплазмы ее стенками. Им предстоит пройти ряд этапов, прежде чем они смогут вызвать инфекционный процесс.
Слияние вирусной, и клеточной мембран. Для того чтобы внутренний компонент вируса мог пройти через клеточную мембрану, вирус использует механизм слияния мембран. У оболочечных вирусов слияние обусловлено точечным взаимодействием вирусного белка слияния с липидами клеточной мембраны, в результате которого вирусная липопротеидная оболочка интегрирует с клеточной мембраной, а внутренний компонент вируса оказывается по другую ее сторону. У безоболочечных вирусов один из поверхностных белков также взаимодействует с липидами клеточных мембран, в результате чего внутренний компонент проходит через мембрану. Большинство вирусов животных выходит в цитозол из рецептосомы.
Если при эндоцитозе вирусная частица является пассивным пассажиром, то при слиянии она становится активным участником процесса. Белком слияния является один из ее поверхностных белков. К настоящему времени этот белок идентифицирован лишь у парамиксовирусов и ортомиксовирусов. У парамиксовирусов этот белок (Р-белок) представляет собой один из двух гликопротеидов, находящихся на поверхности вирусной частицы. Функцию белка слияния у вируса гриппа выполняет малая гемагглютинирующая субъединица.
Парамиксовирусы вызывают слияние мембран при нейтральном рН, и внутренний компонент этих вирусов может проникать в клетку непосредственно через плазматическую мембрану. Однако большинство оболочечных и безоболочечных вирусов вызывают слияние мембран только при низком значении рН — от 5,0 до 5,75. Если к клеткам добавить слабые основания (хлорид аммония, хлороквин и др.), которые в эндоцитарных вакуолях повышают рН до 6,0, слияния мембран не происходит, вирусные частицы остаются в вакуолях, и инфекционный процесс не возникает. Строгая зависимость слияния мембран от значений рН обусловлена конформационными изменениями вирусных белков слияния.
В лизосоме постоянно имеется низкое значение рН (4,9). В эндоцитарной вакуоли (рецептосоме) закисление создается за счет АТФ-зависимого «протонового насоса» еще на клеточной поверхности при образовании покрытой вакуоли. Закисление эндоцитарной вакуоли имеет большое значение для проникающих в клетку физиологических лигандов, так как низкое значение рН способствует диссоциации лиганда от рецептора и рециркуляции рецепторов.
Схематическое изображение возможных способов проникновения вирусов в клетку показано на рис. 8.
Рис. 8. Проникновение вируса в клетку:
а — проникновение вируса путем слияния оболочки с плазматической мембраной;
Раздевание
Проникшие в клетку вирусные частицы должны раздеться для того, чтобы вызвать инфекционный процесс. Смысл раздевания заключается в удалении вирусных защитных оболочек, которые препятствуют экспрессии вирусного генома. В результате раздевания освобождается внутренний компонент вируса, который способен вызвать инфекционный процесс. Раздевание сопровождается рядом характерных особенностей: в результате распада вирусной частицы исчезает инфекционная активность, в ряде случаев появляется чувствительность к нуклеазам, возникает устойчивость к нейтрализующему действию антител, теряется фоточувствительность при использовании ряда препаратов.
Конечными продуктами раздевания являются сердцевины, нуклеокапсиды или нуклеиновые кислоты. Для ряда вирусов было показано, что продуктом раздевания являются не голые нуклеиновые кислоты, а нуклеиновые кислоты, связанные с внутренним вирусным белком. Например, конечным продуктом раздевания пикорнавирусов является РНК, ковалентно связанная с белком VPg, конечным продуктом раздевания аденовирусов, вируса полиомы и SV40 является ДНК, ковалентно связанная с одним из внутренних вирусных белков.
В ряде случаев способность вирусов вызвать инфекционный процесс определяется возможностью их раздевания в клетке данной системы. Тем самым эта стадия является одной из стадий, лимитирующих инфекцию.
Раздевание вирусной частицы ^осуществляется постепенно в результате серии последовательных реакций. Например, в процессе раздевания пикорнавирусы проходят ряд стадий с образованием промежуточных субвирусных частиц с размерами от 156s до 12s. В случае папова-, адено- и герпесвирусов, вероятнее всего, клеточные ферменты дезагрегируют капсид и к моменту экспрессии вирусных функций от вирусной частицы остаются только ДНК или комплекс ДНК—белок. При реовирусной инфекции в клетках удаляется только часть капсида, и, несмотря на то что вирусный геном так и не освобождается из капсида полностью, он экспрессирует все свои функции. Поксвирусы раздеваются в две стадии: на первой ферменты хозяина удаляют наружное покрытие, а на второй для освобождения вирусной ДНК из сердцевины требуется участие продуктов вирусных генов, синтезированных после заражения. Вирус гриппа вначале теряет липопротеидную оболочку и превращается в субвирусную частицу, из которой после удаления М-белка освобождается нуклеокапсид.
Стратегия размножения вируса
Требования и ограничения
В ходе эволюции вирусов сложилось несколько стратегий, обеспечивающих: а) организацию вирусных генов и их кодирующую функцию, б) экспрессию вирусных генов, в) репликацию вирусных геномов и г) сборку и созревание вирусного потомства.
Прежде чем мы рассмотрим каждое из этих положений в деталях, стоит напомнить, что ключевым моментом в репликации вирусов является использование для синтеза вирусных белков хозяйских структур, синтезирующих белки клетки. Независимо от размеров, состава и организации своего генома вирус должен предоставить белоксинтезирующему аппарату эукариоти-ческой клетки информационную РНК, которую клетка должна распознать и транслировать. В этом отношении клетка навязывает вирусу два ограничения.
Во-первых, клетка синтезирует в ядре свою собственную мРНК путем транскрипции своей ДНК и последующего постранскрипционного процессинга транскрипта. Поэтому в клетке: а) ни в ядре, ни в цитоплазме нет фер* ментов, необходимых для транскрипции мРНК с вирусного РНК-генома, и б) в цитоплазме нет ферментов, способных транскрибировать вирусную ДНК. В связи с этим клеточную транскриптазу для синтеза вирусных мРНК могут использовать только вирусы, содержащие ДНК и способные проникать в ядро. Все другие вирусы вынуждены создавать собственные ферменты для синтеза мРНК.
Второе ограничение состоит в том, что синтезирующий аппарат эукариотических клеток приспособлен только для трансляции моноцистронных мРНК, так как он не распознает внутренних участков инициации в мРНК. В результате вирусы вынуждены синтезировать либо отдельные мРНК для каждого гена (функционально моноцистронная мРНК), либо мРНК, включающую несколько генов и кодирующую большой «полипротеин», который затем разрезается на индивидуальные белки.
Экспрессия и репликация вирусного генома
Вирусные гены кодированы либо в РНК, либо в ДНК, которые могут быть либо одно-, либо двухцепочечными. Кроме того, геномы могут быть либо монолитными, когда все гены вируса содержатся в одной хромосоме, либо состоять из раздельных блоков, которые все вместе и составляют геном вируса. Во избежание ошибок мы обозначаем как «геномную» только нуклеиновую кислоту, которая заключена в вирионах.
Для удобства сначала лучше обсудить РНК-содержащие вирусы, фокусируя внимание прежде всего на функции геномной РНК.
Вирусы, содержащие одноцепочечную РНК
Вирусы с одноцепочечной РНК можно подразделить на три группы. В первую группу входят пикорнавирусы и тогавирусы. Их геномы выполняют две функции (рис. 5.2 и 5.3). Во-первых, они функционируют как мРНК. Вирусы, геном которых может служить в качестве мРНК, принято называть вирусами с позитивным геномом. РНК пикорнавирусов после проникновения в клетку связывается с рибосомами и полностью транслируется. Затем продукт этой трансляции — полипротеин — расщепляется. Во-вторых, геномные РНК выполняют функцию матрицы для синтеза на ней комплементарных минус-цепей при участии полимеразы, появившейся в результате расщепления полипротеина. Образуется двуспиральная репликативная форма. На ее минус-цепях синтезируются новые плюс-цепи, которые могут использоваться в качестве а) мРНК, б) матриц для синтеза новых минус-цепей и в) составной части вирусных частиц потомства.
Тогавирусы и ряд других вирусов с позитивным геномом отличаются от пикорнавирусов в одном отношении: для трансляции в первом цикле синтеза белков доступна только часть их геномной РНК. Вероятной функцией образующихся при этом белков является способность транскрибировать геномную РНК. Они осуществляют синтез минус-цепи, которая в свою очередь служит матрицей для синтеза двух различных по размеру классов молекул плюс-РНК. В клетках, зараженных тогавирусами, РНК первого класса представлены небольшими молекулами мРНК, фланкирующими участок геномной РНК, который не транслировался в первом цикле. Образующиеся на них полипротеины расщепляются на белки, которые играют структурную роль в вирионах. Плюс-РНК второго класса состоит из полноразмерных цепей, которые упаковываются в вирионы.
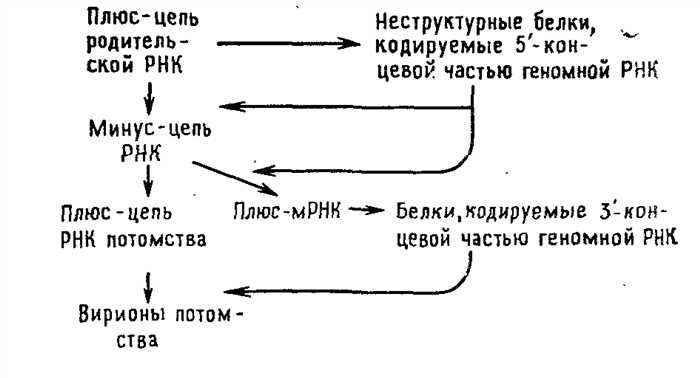
Главным в репликации вирусов с позитивным геномом является способность геномной РНК служить в качестве мРНК после заражения. Это имеет двоякие последствия. Во-первых, ферменты, ответственные за репликацию генома, синтезируются после заражения, и нет необходимости в их внесении в зараженную клетку вместе с вирионом. Вот почему «голая» РНК, экстрагированная из вирионов, инфекционна. Во-вторых, так как все позитивные геномы относятся к монолитным и все их гены сосредоточены в одной хромосоме, первичный продукт трансляции обеих РНК (как геномной, так и мРНК) обязательно представляет собой единый белок. Продукты трансляции пикорнавиру-сов и тогавирусов должны быть затем расщеплены на индивидуальные белки, которые и обнаруживаются в вирионе или в зараженной клетке.
РНК-содержащие вирусы с негативным геномом: ортомиксовирусы, парамиксовирусы, буньявирусы, аренавирусы и рабдовирусы.
Их геномная РНК выполняет две матричные функции: во-первых, для транскрипции и, во-вторых, для репликации. В связи с тем что для синтеза мРНК должен транскрибироваться вирусный геном, а в клетках соответствующие ферменты отсутствуют, все вирусы с негативным геномом содержат в вирионе кроме вирусного генома транскриптазу. Транскрипция вирусного генома — первое событие после проникновения вируса в клетку, в результате которого накапливаются функционально активные моноцистронные мРНК [позитивные, или плюс-цепи], кодирующие один белок. Репликацию начинают новосинтезированные вирусные белки, катализирующие образование полной плюс-цепи, которая служит матрицей для синтеза геномной минус-РНК (рис. 5.4).
Главное в репликации вирусов с негативным геномом заключается в том, что геномная РНК функционирует как матрица и для транскрипции, и для репликации. Отсюда следует, что, во-первых, вирус должен внести с собой в зараженную клетку транскриптазу; во-вторых, «голая» РНК, экстрагированная из вирионов, неинфекционна; в-третьих, синтезируемые мРНК имеют длину одного гена, они кодируют один белок — единичный полипептид. Присутствие сигналов сплайсинга в определенных участках может обеспечить формирование нескольких мРНК (каждая из которых кодирует особый белок) с одного и того же участка генома. Следовательно, плюс-транскрипт, функционирующий в качестве мРНК, отличается от плюс-РНК, служащей матрицей для вирусного потомства, хотя и первый, и вторая синтезируются на геномной РНК. Кроме РНК-транскриптазы в репликации вирусной РНК участвуют вирусиндуцированнные: репликаза 1 (обрзование репликативной формы) и репликаза 2 (синтез вирионных -нитей).
Размножение ортомиксовирусов и парамиксовирусов.
– РНК зависимая РНК полимераза (синтез +нитей,
– две разновидности РНК полимераз
РНК транскриптаза (синтез +нитей в начале инфекции,
РНК репликаза ( – нитей в конце инфекции)
Ретровирусы входят в третью группу РНК-содержащих вируссов.
Характерно, что геномы ретровирусов монолитны, но имеют диплоидную структуру, и обе цепи либо частично соединены водородными связями друг с другом, либо спарены неизвестным до настоящего времени образом. Единственная известная функция геномной РНК — матричная функция для синтеза вирусной ДНК. Поскольку эукариотические клетки не имеют для этого соответствующих ферментов, вирион кроме генома содержит еще и РНК-зависимую ДНК-полимеразу (обратную транскриптазу 1), а также смесь тРНК хозяина, одна из которых служит в качестве затравки. В цикле репродукции можно выделить следующие ключевые ступени: а) связывание комплекса тРНК —обратная транскриптаза с геномной РНК; б) синтез ДНК-копии, комплементарной по отношению к РНК, с переходом полимеразы с одной молекулы РНК-матрицы на другую, что приводит к образованию кольцевой одноцепочечной молекулы ДНК, связанной водородными связями с линейной геномной РНК; в) расщепление геномной РНК нуклеазой, атакующей только РНК в ДНК—РНК-гибридах (рибонуклеазои Н, также содержащейся в вирионе), и г) синтез комплементарной копии вирусной ДНК. Затем кольцевая двухцепочечная ДНК перемещается в ядро, где интегрирует с геномом хозяина, но последующая экспрессия вирусных генов не обязательна. Если экспрессия происходит, то интегрированная.вирусная ДНК транскрибируется транскриптазой клетки-хозяина. Продуктами транскрипции являются молекулы РНК, как равные по длине молекуле генома, так и более короткие мРНК-транскрипты нескольких соседних генов, которые транслируются с образованием полипротеинов. Полипротеины затем расщепляются на отдельные вирусные белки. В состав вирионов включаются только транскрипты, содержащие весь геном.
Читайте также: