
Какой белок рецептор является необходимым для присоединения вич к
После заражения ВИЧ проникает в лимфоидные клетки Лангерганса, которые локализуются в слизистой и в коже, затем инфицируются макрофаги и CD4+-лимфоциты, с которыми вирус распространяется по всему организму. Клетками-мишенями ВИЧ являются Т-лимфоциты, дендритные лимфоциты и клетки Лангерганса (их незрелые предшественники), моноциты/макрофаги, эозинофилы, мегакариоциты, тимоциты, некоторые клоны В-лимфоцитов, клетки нервной системы - нейроны, микроглиальные клетки/макрофаги. Все эти клетки объединяет наличие на их мембране рецептора СD4* (* от англ. Cell Differentiation Antigen 4 - дифференцировочный клеточный антиген № 4. Номер соответствует порядковому номеру очередного препарата (ОКТ-4) серии моноклональны.х антител, полученных фирмой "ORTHO" (США) для исследования рецепторов лимфоцитов), к которому имеет большое сродство вирусный эпимембранный гликопротеин gp120 (ВИЧ-1) и gp105 (ВИЧ-2) [25].
Вместе с тем следует заметить, что существует ряд клеток, которые, не имея рецептора СD4, селективно сорбируют, транспортируют на мембране или проводят через себя ВИЧ. К такого типа клеткам относятся М-клетки слизистой прямой кишки, граничащие с лимфоидной тканью стенки кишки, и сперматозоиды.
Из перечисленных клеток-мишеней, несущих на мембране CD4, важнейшими в патогенетическом плане являются Т-лимфоциты, выполняющие функцию хелперов/индукторов и эффекторов, их количество является наибольшим среди клеток, несущих рецептор CD4, в системе крови человека (табл. 9.1.)
| Таблица 9.1. Прямые клетки-мишени для ВИЧ в системе крови человека, имеющие на мембране рецептор CD4 | |
| Тип дифференцированных клеток | Содержание в крови. %* (норма) |
Т-лимфоциты:
|
|
| Дендритные лимфоциты | 1 |
| Моноциты/макрофаги | 3-1 |
| Эозинофилы | 0,5-5 |
| В-лимфоциты (некоторые клоны) | 1,8-8,5 |
| * от числа ядросодержащнх клеток крози. ** в основном несут на мембране рецептор СD8. | |
По тропизму изоляты ВИЧ делят на моноцитотропные и лимфоцитотропные. Первые преобладают на начальных стадиях болезни, вторые - в период разгара.
Процесс взаимодействия ВИЧ с клеткой-мишенью включает ряд последовательных стадий. Проникновению вируса внутрь клетки предшествует взаимодействие оболочечных белков вируса с молекулами, экспрессированными на мембране клетки. Необходимым условием фиксации ВИЧ на клеточной мембране и последующего проникновения его в клетку является не только наличие рецептора CD4, с которым соединяется белок gp120 (ВИЧ-1) или gp105 (ВИЧ-2), но и корецептора. Для макрофаготропной разновидности ВИЧ таковым является хемокиновый рецептор ССR5, для лимфоцитотроп-ой - СХСR4.
Хемокиновый рецептор обеспечивает конформационное изменение трансмембранпого белка gр41 (ВИЧ-1) и gр36 (ВИЧ-2), необходимое для слияния вируса с клеточной мембраной. Некоторые варианты ВИЧ могут использовать оба корецептора.
У 2% населения на макрофагах отсутствует хемокиновый рецептор ССR5 вследствие генетических мутаций, что обусловливает невосприимчивость таких людей к ВИЧ-инфекции.
На следующей стадии оболочка зафиксированного на клеточной мембране вириона сливается с клеточной мембраной, сердцевина освобождается от оболочки (стадия "раздевания") и проникает в клетку. При этом вполне возможно, что вирусная оболочка становится частью наружной мембраны клетки-хозяина. Проникшая в цитоплазму свободная сердцевина вируса раскрывается. Далее с помощью обратной транскриптазы по матрице вирусной геномной РНК осуществляется синтез ДНК, для чего используются азотистые основания из цитоплазмы. Молекулы вирусной ДНК приобретают кольцевидную форму (циклизация) и проникают из цитоплазмы в ядро, где встраиваются (интеграция) в состав гепома клетки-хозяина [16].
Таким образом, процесс инфицирования вирусом иммунодефицита человека клетки-мишени можно разделить на следующие стадии:
- Связывание вириона с поверхностью клетки. Рецепция вируса.
- Слияние мембран вируса и клетки. Проникновение вируса внутрь клетки.
- Высвобождение нуклеоида и геномной РНК вируса.
- Синтез провирусной ДНК по матрице геномной РНК вируса.
- Интеграция генома провируса в геном клетки.
- Активация процесса транскрипции с ДНК провируса, трансляция белков вируса.
- Активная репликация вируса, то есть продукция всех компонентов вируса и формирование из них зрелых дочерних вирионов.
- Высвобождение вирионов и отдельных белков ВИЧ из клетки-хозяина во внешнюю среду и беспрепятственное заражение других клеток. Цитопатогенные эффекты ВИЧ.
Ведущим звеном в патогенезе ВИЧ-инфекции является поражение Т-хелперов [19], которое обусловлено:
- преждевременным старением и гибелью инфицированных клеток
- уничтожением зараженных клеток лимфоцитами-эффекторами антителозависимой клеточной цитотоксичности;
- блокадой рецепторов CD4 вирусным гликопротеином gр120;
- аутоиммунными процессами.
На Т-хелперы ВИЧ оказывает прямой цитопатогенный эффект. Истощение пула Т-хелперов приводит к тому, что они не могут полноценно обеспечивать функцию и взаимодействие других иммунокомпетентных клеток. Однако и на ранних этапах течения ВИЧ-инфекции, когда еще нет выраженного снижения содержания CD4+-клеток, а доля инфицированных CD4+-лимфоцитов не превышает 0,01% их числа [21], основное значение в развитии дисбаланса иммунного ответа, формировании иммунодефицита принадлежит нарушениям регуллториых функций Т-хелперов/индукторов |2б|. Причиной указанных нарушений является блокада рецептора СD4.
Распознавание Т-хелперами антигенов на поверхности антигенпрезептирующих клеток (например, макрофагов) происходит с обязательным участием двух структур - рецептора CD4, который взаимодействует с главным комплексом гистосовместпмостп II класса (ГКГС II) макрофага, и рецептора для антигена. Связывание только молекулы CD4 каким-либо лигандом (например, анти-CD4-антителом или вирусным белком gp120) в отсутствие последующего задействования рецептора для антигена является для клетки "отрицательным сигналом". При этом происходит быстрая и существенная элиминация молекул CD4 с поверхности клетки. В результате такой лимфоцит не может нормально взаимодействовать с белками главного комплекса гистосовместимости II класса (ГКГС II), находящегося на антигенпрезентирующих клетках, что является необходимым условием для формирования нормального иммунного ответа.
Взаимодействие поверхностного гликопротеина gр120 с мембраной CD4+-клеток может не только вызывать негативные сигналы, но и приводит к программированной клеточной гибели - апоптозу зрелых CD4+-лимфоцитов или гематопоэтнческих СD34+-клеток-предшественников даже при отсутствии инфицирования клеток ВИЧ.
G. Furlini с соавт. [18] установили, что спустя три часа после воздействия ВИЧ-1 (или очищенного рекомбинаитпого белка gp120) на CD4+-клетки in vitro наблюдается пик увеличения синтеза и ядерной транслокации белков теплового шока семейства БТШ-70. Эти данные свидетельствуют о способности gp120 запускать каскад процессов, используя сигнальную активность мембран. Одним из таких внутриклеточных процессов является активация системы белков теплового шока, что в свою очередь указывает на нахождение клетки в неблагоприятных условиях и формирование клеточной стресс-реакции.
Таким образом, не только полноценные вирусы, прямо инфицирующие Т-лимфоциты-хелпeры, но и отдельный растворимый вирусный белок gp120 вызывают иммуносупрессию путем связывания молекул CD4. Особенно значительный иммуносупрессорный эффект оказывает агрегированный (например, специфическими антителами) белок gp120 [26].
Гомология аминокислотных последовательностей молекул вирусного белка gp120 и рецепторпых структур клеток (HLА класса II и CD4) обусловливает появление перекрестно реагирующих антител (аутоантител), которые нарушают кооперативные взаимодействия иммунокомпетентных клеток. Аутоиммунная реакция формируется и в отношении иммуноглобулинов, под структуру которых мимикрирует gp120.
Нарушение функции Т-хелперов влечет за собой снижение цитотоксической активности Т-супрессоров, на которые ВНЧ также оказывает цитопатогенный эффект, что, в свою очередь, обусловливает появление оппортунистических инфекции и СПИД-индикаторных опухолей.
С развитием выраженной виремии количество инфицированных клеток в крови и интенсивность их гибели возрастает. От момента инфицирования до терминальной стадии СПИД содержание CD4+-клеток уменьшается более чем в 20 раз.
В моноцитах/макрофагах, в отличие от Т-лимфоцитов, ВИЧ реплицируется с умеренной интенсивностью, вирионы оформляются в округлые частицы еще в цитоплазме клетки и, выходя из нее, не оказывают цитонекротического действия. Однако моноциты/макрофаги, инфицированные ВИЧ, хотя и не подвергаются быстро наступающему цитолизу при массовом высвобождении дочерних вирионов, как Т-лимфоциты, но претерпевают значительные ультраструктурные преобразования. Указанные изменения, видимо, являются причиной пониженной бактерицидной (фунгицидной) активности и способности к хемотаксису моноцитов и макрофагов, а также ослабления взаимодействия их рецепторов с Fс-фрагментами иммуноглобулинов.
В-лимфоциты, учитывая отсутствие или невысокую плотность CD4-реиепторов на их цитоплазматнческой мембране, должны повреждаться в меньшей степени. Косвенным подтверждением этого, казалось бы, служит тот факт, что суммарная концентрация иммуноглобулинов IgG и IgА сыворотки в условиях ВИЧ-инфекции оказывается повышенной. Однако у больных отмечается характерная диспропорция уровней подклассов иммуноглобулинов IgG. Так, показано, что содержание IgG1 и IgG3 у таких пациентов увеличено, тогда как концентрация IgG2 и IgG4 существенно уменьшена. Прогрессирующее снижение уровня IgG2 может объяснить возрастающую восприимчивость больных ВИЧ-инфекцией к патогенному действию таких микроорганизмов, как Haemophilus influenzae, Streptococcus pneumoniae и Staphylococcus aureus.
В-лимфоциты на фоне активной секреции антител характеризуются слабой реакцией на митогeны и на неоантигены. Таким образом, несмотря на гипергаммаглобулинемию, функциональное состояние В-системы иммунитета у больных ВИЧ-инфекцией сходно с состоянием, развивающимся на фоне выраженной гипогаммаглобулинемни [4].
Функционирование В-лимфоцитов контролируется Т-лимфоиитами. Поэтому, вероятно, дисфункции В-системы в целом могут быть вторичными по отношению к дисфункции Т-хелперов и Т-супрессоров. Кроме того, ВИЧ способен прямо инфицировать В-лимфоциты и вызывать их разрушение. Опыт показывает, что содержание В-лимфоцитов в периферической крови больных в стадии СПИД может быть в три с лишним раза ниже нормы.
Вторым объектом воздействия ВИЧ после иммунной системы является ЦНС. ВИЧ инфицирует нейроны, астроциты, микроглиальные клетки (макрофаги), эндотелий кровеносных сосудов, фибробластоподобные клетки мозга. Однако прямого цитопатогенного эффекта вируса на указанные клетки не отмечено. Поражение ЦНС, проявляющееся деменцией и другими неврологическими нарушениями при ВИЧ-инфекции, скорее связано с нейротоксическим действием gp120 и появлением аутоантптел против антигенов мозговой ткани [15].
Таким образом, прогрессированию течения ВИЧ-инфекции предшествует активация репликативного процесса вируса иммунодефицита человека. Клинические проявления заболевания обусловлены непосредственным патогенным эффектом вируса и его белков на клетки-мишени, истощением пула CD4+-клеток крови, а также нарушением кооперативных связей и функций иммунокомпетентных клеток, что приводит к формированию иммунодефицита.
| Страница 1 2 3 4 5 6 7 8 | всего страниц: 8 |
- Федеральный закон РФ "О предупреждении распространения в Российской Федерации заболевания, вызываемого вирусом иммунодефицита человека" от 30 марта 1995 года.
- Змушко Е. И., Белозеров Е. С. ВИЧ-инфекция / Руководство для врачей. - СПб: Питер, 2000. - 320 с.
- Исаков В. А., Аспель Ю. В., Богоявленский Г. В. и др. Опыт применения циклоферона в терапии ВИЧ-инфекцни и СПИДа / Руководство для врачей.- СПб, 1997.- 60 с.
- Кожемякин Л. А., Бондаренко И. Г., Тяптин А. А. Синдром приобретенного иммунодефицита / Пособие для врачей.- Л.: Знание, 1990.- 112 с.
- Лобзин Ю. В., Казанцев А. П. Руководство по инфекционным болезням. - СПб., 1996. - 712 с.
- Лысенко А. Я., Турьянов М. X., Лaвдовская М. В., Подольский В.М. ВИЧ-инфекция и СПИД-ассоциированные заболевания / Монография.- М.: ТОО "Рарогъ", 1996,- 624 с.
- Новохатский Л. С., Хлябич Г. Н. Теория и практика лабораторной диагностики синдрома приобретенного иммунодефицита (СПИД). - М.: ВИНИТИ, 1992,- 221 с.
- Покровский В. И., Покровский В. В. СПИД: синдром приобретенного иммунодефицита.- М.: Медицина, 1988.- 43 с.
- Покровский В. И. ВИЧ-инфекция или СПИД // Терапевт, арх. - 1989. - Т. 61, № 11. - С. 3-6.
- Покровский В. В. ВИЧ-инфекция: клиника, диагностика / Под общ. ред. В. В. Покровского.- М.: ГЭОТАР МЕДИЦИНА, 2000.- 496 с.
- Рахманова А. Г. ВИЧ-инфекция (клиника и лечение).- СПб: "ССЗ", 2000.- 367 с.
- Рекомендации по применению антиретровирусных препаратов у взрослых и подростков, инфицированных вирусом иммунодефицита человека // Consilium Medicum приложение. Январь 2000,- 22 с.
- Смольская Т. Т., Ленинская П. П., Шилова Э.А. Серологическая диагностика ВИЧ-инфекции / Методическое пособие для врачей.- СПб, 1992.- 80 с.
- Смольскал Т. Т. Второе десятилетие жизни в условиях СППДа: уроки и проблемы / Актовая речь.- СПб., 1997.- 56 с.
- Хаитов Р.М., Игнатьева Г. А. СПИД.- М., 1992.- 352 с.
- Connor S. Research shows how HIV exhausts the body // Brit. Mod. J.- 1995.- Vol.310.- P. 6973-7145.
- Burcham J., Marmor M., Dubin N. et al. CD4 is the best predictor of development of AIDS in a cohort of HIV-infecteci homosexual men // J. AIDS.- 1991.- jN"9. - P.365.
- Furlini G., Vignoli M., Re M. C., Gibellini D., Ramazzotti E., Zauli G.. La Placa M. Human immunodeficiency virus type I interaction with the membrane of CD4+ cells induces the synthesis and nuclear translocation of 70K heat shock protein // J.Gen. Virol.- 1994.- Vol.75, pt 1.- P. 193-199.
- Gallo R. C. Mechanism of disease induction by HIV // J.AIDS.- 1990.- N3.- P. 380-389.
- Gottlieb M. S., Schroff R., Schanker H. et al. Pneumocystis carinii pneumonia and mucosal candidiasis in previously homosexual mon // Now England J. Med. - 1981. - Vol. 305. - P. 1425-1430.
- Harper M. E., Marselle L. M., Gallo R.C., Wong-Staal F. Detection of lymfocytes expressing human T-lymphotropic virus type III in limph nodes and peripheral blood from infected individuals by in situ hybridization // Proc. Natl. Acad. Sci. U. S. A. - 1986. - Vol. 83. - N 2. - P. 772-776.
- Hess G. Clinical and diagnostic aspects of HIV-infection.- Mannheim: Boehringer Mannheim GmbH, 1992.- 37 p.
- Hu D.J., Dondero T.J., Ryefild M. A. et al. The emerging genetic diversity of HIV // JAMA.- 1996. - N 1.- P. 210-216.
- Lambin P., Desjobert H., Debbia M. et al. Serum neopterin and beta-2-microglobulin in anti-HIV positive blood donors // Lancet.- 1986.- Vol.8517. - P. 1216.
- Maldonado I. A., Retru A. Diagnosis of pediatric HIV disease // The AIDS knowledge base, Fd. Cohen P.T.; Sande M. A. Voiberding. 1994.- P. 8.2.1-8.2.10.
- Mc Dougal J.S., Kennedy M.S., Sligh J.M. et al. Binding of the HTLV-III/LAV to T4+ T cells by a complex of the 110K molecule and the T4 molecule // Science.- 1985.- Vol.23.- P. 382-385.
- Montagnier L., Gougeon M. L., Olivier R. et al. Factors and mechanisms of AIDS pathogenesis // Science challenging AIDS. Basel: Karger, 1992.- P. 51-70.
- Paterlini P., Lallemant-Le C., Lallemant M. et al. Polimerase chain reaction for studies of mother to child transmission of HIV-I in Africa // J.Med. Virol. - 1990.- Vol.30, N 10.- P. 53-57.
- Polis M. A., Masur H. Predicting the progression to AIDS // Amor. J. Med. - 1990.- Vol.89, N 6.- P. 701-705.
- Roddy M.M., Grieco M. H. Elevated soluble IL-2 receptor levels in serum of HIV infected populations // AIDS Res. Hum. Retrovir. - 1988.- Vol.4, N 2. - P. 115-120.
- Van dor Groen. G., Van Kerckhoven I. et al. Упрощенный и менее дорогой, в сравнении с традиционным, способ подтверждения инфекции ВИЧ // Бюлл. ВОЗ.- 1991.- Т. 69, №6.- С. 81-86.
Источник: Медицинская лабораторная диагностика, программы и алгоритмы. Под ред. проф. Карпищенко А.И., СПб, Интермедика, 2001
Медицинский эксперт статьи

Возбудитель ВИЧ-инфекции
Описано два серотипа - ВИЧ-1 и ВИЧ-2, различающиеся по структурным и антигенным характеристикам. На территории Украины эпидемиологическое значение имеет ВИЧ-1 (основной возбудитель заболевания).
Систематика ВИЧ
Вирус иммунодефицита человека относят к царству Viridae, семейству Retroviridae, подсемейству Lentiviridae.
Основные свойства вируса иммунодефицита человека
По данным электронной микроскопии, вирус имеет округлую форму и сложную структуру. Диаметр вириона составляет 100-120 нм.
Группы белков ВИЧ-1 и ВИЧ-2
Белки оболочки вируса (env)
gp160, gp120, gp41
gр140, gр105, gр36
Белки сердцевины (gag)
Ферменты вируса (pol)
Молекулярный вес белков измеряют в килодальтонах (кДа): gp - гликопротеины; р - протеины.
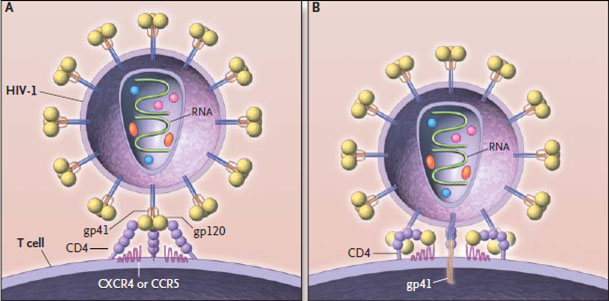
В центре вириона располагается геном вируса, представленный двумя нитями РНК, внутренними протеинами р7 и р9, а также ферментами - обратной транскриптазой (ревертазой), протеазой, РНКазой и интегразой (эндонуклеазой). Геном окружает внутренняя белковая оболочка. В состав внутренней оболочки ВИЧ-1 входят протеины р17, р24 и р55. Протеины р16, р25 и р56 образуют внутреннюю оболочку ВИЧ-2. Наружная липидная оболочка ВИЧ-1 пронизана гликопротеином gpl60. состоящим из трансмембранного (gp41) и высокоиммуногенного (gpl20) фрагмента. Оболочечные белки gp41 и gpl20 соединены нековалентной связью и формируют на поверхности вириона отростки, обеспечивающие присоединение ВИЧ к рецепторам клеток-мишеней человека.
Геном вируса содержит девять генов - три структурных и шесть регуляторных. Геном является изменчивой структурой за счёт антигенного дрейфа. Существует ряд серологических вариантов вируса (например, А, В, С, D, E, F, G, Н).

[1], [2], [3], [4], [5], [6], [7], [8], [9], [10], [11], [12], [13], [14], [15]
Устойчивость вируса в окружающей среде
В естественных условиях ВИЧ (в высушенном состоянии) сохраняет активность в течение нескольких часов; в жидкостях, содержащих большое количество вирусных частиц, таких, как кровь и эякулят, - в течение нескольких дней.
В замороженной сыворотке крови активность вируса определяют на протяжении нескольких лет.
Нагревание до 56 °С в течение 30 мин приводит к снижению инфекционного титра вируса в 100 раз. При более высокой температуре (70-80 °С) вирус погибает через 10 мин. При обработке вирионов 70% раствором этилового спирта в течение минуты происходит их инактивация. При воздействии 0.5% раствора гипохлорита натрия, 1% раствора глутаральдегида, 6% раствора пероксида водорода, 5% раствора лизола, эфира или ацетона также отмечают гибель вирусных частиц.
ВИЧ относительно мало чувствителен к ультрафиолетовому облучению и ионизирующей радиации.
Патогенез ВИЧ-инфекции
Дифференцировочный антиген CD4+ (CD - аббревиатура от Cell Differention antigen) и неспецифические (независящие от наличия CD4+) компоненты - рецепторы для ВИЧ. CD4+ - гликопротеид с молекулярной массой 55 000 кД. по своему строению схожий с определёнными участками иммуноглобулинов. Аналогичное строение имеет и белок вируса gpl20 - это и определяет способность проникновения вирионов в клетку.
Рецептор CD4+, расположенный на мембране иммунокомпетентных клеток, выполняет функцию распознавания антигенов (вместе с белками HLA - главного комплекса гистосовместимости II класса).
Типы клеток, поражаемых вирусом иммунодефицита человека
Фолликулярные дендритные клетки
Толстый кишечник, почки
Клетки шейки матки
Оболочка ВИЧ содержит белки человеческого комплекса гистосовместимости I и II классов, поэтому проникновение вируса в организм не вызывает реакции отторжения. Фиксация вирионов на поверхности клетки-мишени происходит при участии гликопротеина gpl20. Гликопротеин gp41 обеспечивает слияние вирусной оболочки с мембраной клетки-мишени. Двуцепочечная РНК вируса проникает в клетку, где с помощью обратной транскриптазы происходит синтез одноцепочечной провирусной ДНК. Затем происходит формирование двуцепочечной ДНК, встраивающейся в ДНК клетки с помощью интегразы. Вирусная ДНК выполняет функцию матрицы для синтеза РНК, собирающей новую вирусную частицу.
Нередко при репликации ВИЧ возникают генетические ошибки. Таким образом происходит образование разных подтипов вируса.
После проникновения ВИЧ в СD4+-клетки начинается его репликация: чем активнее СD4+-клетки, тем интенсивнее репродукция вируса. Поэтому регуляторы, активирующие CD4+-клетки обеспечивают увеличение репликации вируса. К подобным регуляторам относят ФНО: фактор, стимулирующий колонии гранулоцитов и макрофагов (колониестимулирующий фактор), и ИЛ-6.
Интерферон и трансформирующий фактор роста - регуляторы, тормозящие репликацию вируса. Как показали исследования. ФНО-а активирует транскрипцию провирусной ДНК ВИЧ-1 в хронически инфицированных Т-клетках и макрофагах. Моноциты, синтезирующие ФНО-а. не только индуцируют экспрессию ВИЧ-инфицированными клетками, но и стимулируют активацию латентного провируса. Регистрируют одновременную активность ФНО-а. ИЛ-6 и фактора, стимулирующего колонии гранулоцитов и макрофагов.
Иммунопатогенетические признаки ВИЧ-инфекции - дефицит Т-звена и В-звена иммунной системы: недостаток компонентов комплемента и фагоцитов; снижение функций неспецифических факторов защиты. Поликлональная активация В-лимфоцитов приводит, с одной стороны, к гипергаммаглобулинемии, а с другой - к ослаблению способности клеток продуцировать вируснейтрализующие антитела. Происходит увеличение количества ЦИК и образование антител к лимфоцитам; это вызывает ещё большее уменьшение числа СD4+-Т-лимфоцитов. Отмечают развитие аутоиммунных процессов. Поражение иммунной системы при ВИЧ-инфекции носит системный характер.
Наряду с дефицитом СD4+-лимфоцитов в течение болезни нарастает функциональная недостаточность СD8+-лимфоцитов, NK-клеток (естественных киллеров) и нейтрофилов. При ухудшении иммунного статуса развиваются различные инфекционные, аллергические, аутоиммунные и лимфопролиферативные заболевания, а также синдром, свойственный болезни иммунных комплексов (эти факторы определяют клиническую картину ВИЧ-инфекции).
На начальных стадиях заболевания в организме вырабатываются вируснейтрализующие антитела, подавляющие активность свободно циркулирующих вирусов. Однако такие антитела не действуют на вирусы, находящиеся в клетках (провирусы). С течением времени (обычно после 5-6 лет) происходит истощение защитных возможностей иммунной системы и, следовательно, накопление вируса в крови.
14 октября 2013
- 19572
- 15,7
- 2
- 3
Распространенность СПИДа в мире на 2009 год.

Спонсор конкурса — дальновидная компания Thermo Fisher Scientific. Спонсор приза зрительских симпатий — фирма Helicon.
Иммунная система
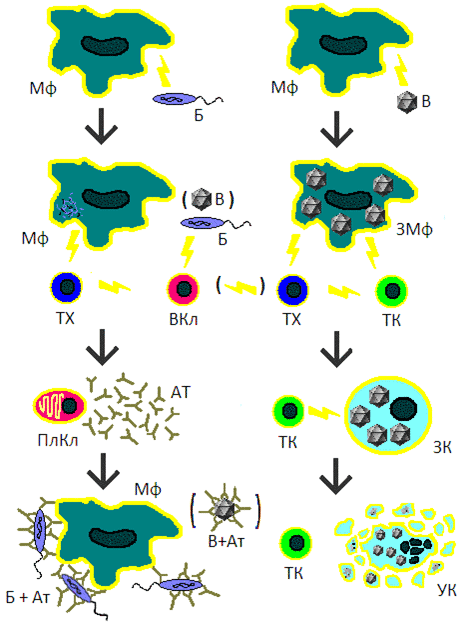
Рисунок 1. Упрощенная схема иммунного ответа. Молниями указано, как один тип клеток активирует другие. Мф — макрофаг, Б — бактерия, ТХ — Т-хелпер, ВКл — B-клетка, ПлКл — плазматическая клетка, Ат — антитела, Б+Ат — бактерия, покрытая антителами, привлекающими макрофагов, В — вирус, ЗМф — зараженный макрофаг, ТК — Т-киллер, ЗК — клетка, зараженная вирусом, УК — умирающая (апоптотирующая) клетка, В+Ат — вирус, покрытый антителами.
рисунок автора статьи
Жизненный цикл ВИЧ
Видео. Жизненный цикл ВИЧ.
Обратная транскрипция осуществляется вирусным белком обратной транскриптазой [31]. Обратная она потому, что обычно в клетке все наоборот — информация переносится с ДНК на РНК (а с РНК — в последовательность аминокислот белков). РНК по сравнению с ДНК очень нестабильна, и поэтому для ВИЧ характерна огромная скорость мутации — в десятки тысяч раз быстрее, чем для человека. Если скорость нейтральных мутаций для ДНК составляет у различных видов и для различных генов в среднем меньше 10 −9 замен на сайт в год, то скорость мутирования генетического материала РНК-вирусов — около 10 −3 замен на сайт в год (у гена env ВИЧ: 10 −2 –10 −3 замен на сайт в год [21]). Это одна из причин, почему иммунная система не может справиться с ВИЧ — он слишком быстро изменяется.
Таким образом, после того как копия вируса в геноме клетки начинает действовать, на её поверхности появляются знакомые нам белки gp41 и gp120, в цитоплазме — остальные вирусные белки и вирусная РНК. И через некоторое время от зараженной клетки начинают отпочковываться всё новые и новые копии ВИЧ.
Способы уничтожения иммунной системы
При отпочковывании вирус использует клеточную мембрану клетки, и со временем это должно вызвать ее разрушение. К тому же, вирусный белок Vpu вызывает увеличение проницаемости мембраны клетки [6]. При активации вируса в клетке появляется неинтегрированная в геном двуцепочечная ДНК [19], наличие которой может рассматриваться клеткой как повреждение ее генетического материала и индуцировать ее смерть через апоптоз (С точки зрения организма такая клетка потенциально может стать раковой). Помимо этого, вирусные белки напрямую нарушают баланс про- и противоапоптотических белков в клетке. К примеру, вирусная протеаза p10 может разрезать противоапоптотический фактор Bcl-2. Белки Nef [26], Env [22] и Tat [24] вовлечены в возрастание количества клеточных белков CD95 и FasL, служащих индукторами апоптоза по т.н. Fas-опосредованному пути. Белок Tat положительно регулирует каспазу 8 [2] — ключевой фактор индукции апоптоза.
Если же клетки не погибли сами, они активно уничтожаются Т-киллерами. (Это стандартный ответ иммунитета на заражение любыми вирусами.)
Помимо этого, при производстве gp120 и Tat часть их выбрасывается зараженной клеткой в кровоток, а это оказывает токсическое действие на организм [4], [28]. gp120 оседает на все СD4 + -клетки (в том числе и здоровые), что имеет три последствия:
- Белок СD4 важен для взаимодействия Т-хелпера с фагоцитом, при слипании же СD4 с gp120 T-хелпер перестает выполнять эту функцию.
- На gp120 как на чужеродный белок образуются антитела; после оседания gp120 на здоровые клетки, последние маркируются антителами, и иммунная система их уничтожает [9].
- После обильного связывания Т-хелпером gp120 клетка умирает сама. Дело в том, что CD4 — это корецептор. Он усиливает сигнал от TCR (T-cell receptor) — главного белка, позволяющего выполнять Т-клеткам функцию иммунитета. В норме эти рецепторы активируются вместе. Если же активируется один тип рецепторов, но не активируется другой, это является сигналом, что что-то пошло не так, и клетка умирает посредством апоптоза.
Антитела, призванные препятствовать инфекции, в случае с ВИЧ часто, наоборот, ее усиливают [28]. Почему так происходит — не совсем понятно. Одну причину мы рассмотрели выше. Возможно, дело в том, что антитела сшивают несколько вирусных частиц вместе, и в клетку попадает не один, а сразу много вирусов. Помимо этого, антитела активируют клетки иммунитета, а в активированной клетке вирусные белки синтезируются быстрее.
Вдобавок, gp120 и gp41 имеют участки, похожие на участки некоторых белков, участвующих в иммунитете (например, MHC-II [25], IgG [23], компонент системы комплемента Clq-A [14]). В результате на эти участки образуются антитела, способные помимо вируса маркировать совершенно здоровые клетки (если на них будут эти белки), с соответствующими для них последствиями.
Так как на поверхности зараженной клетки появляется gp120, то она, как и вирусная частица, будет сливаться с другими СD4-содержащими клетками, образуя огромную многоядерную клетку (синцитий), неспособную выполнять какие-либо функции и обреченную на смерть.
На самом деле, помимо Т-хелперов, СD4 содержатся во множестве других клеток — предшественниках Т-киллеров/Т-хелперов, дендритных клетках, макрофагах/моноцитах, эозинофилах, микроглии (последние четыре — разновидности фагоцитов), нейронах (!), мегакариоцитах, астроцитах, олигодендроцитах (последние две — клетки, питающие нейроны), клетках поперечно-полосатых мышц и хорионаллантоиса (присутствует в плаценте) [29]. Все эти клетки способны заражаться ВИЧ, но сильно токсичен он лишь для Т-хелперов. Это и определяет основное клиническое проявление СПИДа — сильное снижение количества Т-хелперов в крови. Однако для других клеток заражение тоже не проходит бесследно — у больных СПИДом часто снижена свертываемость крови, имеются неврологические и психиатрические отклонения [16], наблюдается общая слабость.
Пути заражения ВИЧ
Внимание! Ввиду того, что СПИД является венерическим заболеванием, информация, приведенная в этом разделе, может вас оскорбить либо вызвать неприятные эмоции при прочтении. Будьте осторожны.
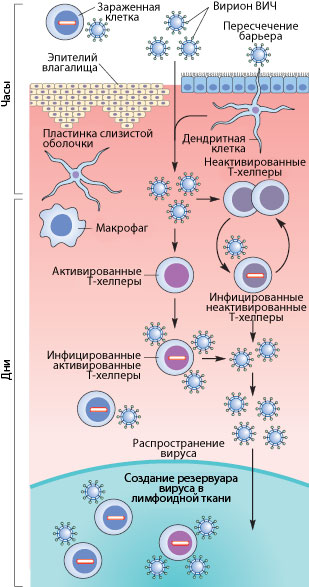
Рисунок 3. Пути проникновения ВИЧ в организм при половом контакте. Слева указано примерные временные рамки каждого этапа.
Как же происходит первичное заражение человека ВИЧ? Думаю, что с переливанием крови или с использованием общего шприца при употреблении наркотиков читателю все понятно — вирус доставляется напрямую из зараженной крови в здоровую. При вертикальном способе передачи инфекции (от матери к ребенку) заражение может произойти во время внутриутробного развития, в момент родов или в период кормления ребенка грудью (т.к. вирус и зараженные вирусом клетки содержатся в молоке матери) [17], [20]. Тем не менее, даже у ребенка, рожденного от ВИЧ-инфицированной матери, есть шанс остаться здоровым, особенно если мать использует ВААРТ (высокоактивную антиретровирусную терапию) [5]. А почему же люди заражаются при половом контакте?
Механизмы передачи вируса при половом акте не до конца ясны. Тем не менее, они понятны в общих чертах. Итак, рассмотрим барьеры, которые необходимо преодолеть вирусу, чтобы добраться до макрофагов и Т-хелперов. К сожалению, рассматривать особо нечего: барьер только один — это слой эпителиальных клеток и вырабатываемая ими слизь, находящиеся во влагалище (а также пенисе, прямой кишке, глотке). Барьер этот тонок — он может быть всего одну клетку в толщину, — но для вируса труднопреодолим, доказательством чего может служить сравнительно небольшой процент заражений — примерно 1–2 случая на 100 половых контактов. Итак, и какими же путями преодолевает вирус это препятствие?
Наиболее очевидный механизм — это микротравмы. При обычном половом контакте, а особенно при анальном сексе, почти неизбежно появляются микротрещины; при наличии же венерических заболеваний часто возникают изъязвления слизистой. Через эти микротравмы вирус, содержащийся в сперме и влагалищном секрете, может проникнуть к иммунокомпетентным клеткам [17], [20].
Помимо дендритных клеток, в кишечнике существует другой тип клеток, предназначенный для ознакомления организма с антигенами внешней среды. Это так называемые М-клетки, пропускающие через себя довольно крупные объекты неповрежденными из просвета кишечника. В т.ч. через них может проходить и вирион ВИЧ.
Свой вклад в заражение вносит и провоспалительный процесс, в норме присутствующий в женских половых путях из-за наличия в них спермы. Благодаря ему Т-клетки подвержены активации, что способствует их восприимчивости к инфекции.
Итак, риск заразиться ВИЧ возрастает в ряду (Классический секс → Анальный секс → переливание крови / использование общих шприцов при принятии наркотиков). Что же касается орального секса, то весьма небольшой шанс заразиться есть только у принимающей стороны — слюна и тем более желудочный сок инактивирует ВИЧ. Теоретически, этим способом может заразиться и мужчина, но для этого необходимы ранки на пенисе.
При поцелуях и обычных бытовых контактах вирус не передается — он очень неустойчив в окружающей среде .
Заключение
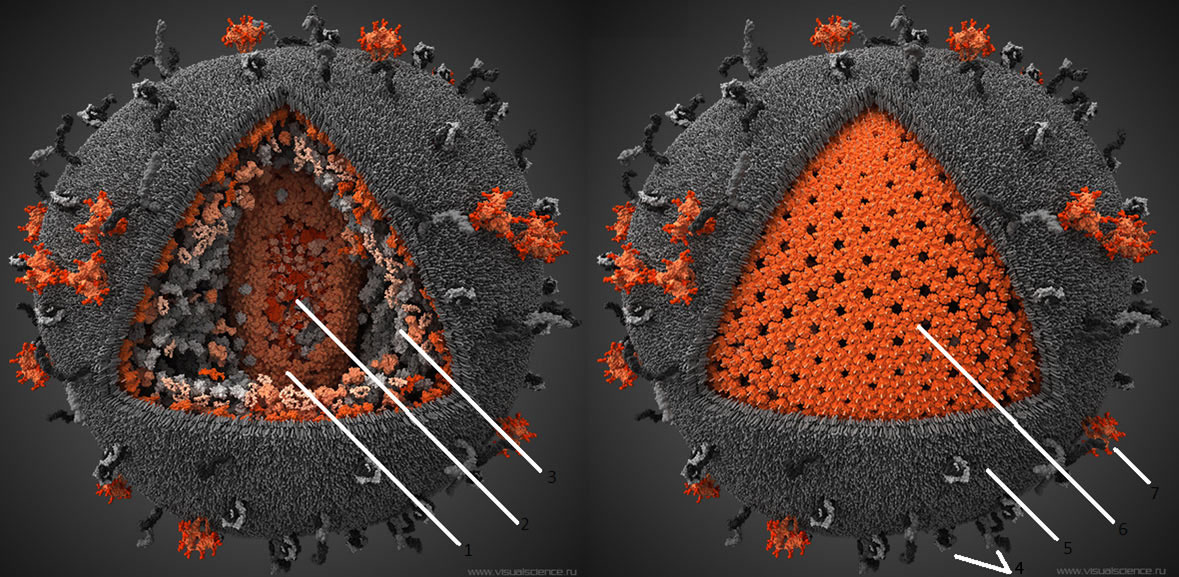
10000 нуклеотидов в каждой. Всего геном включает девять генов, которые в результате альтернативного сплайсинга кодируют 15 различных белков. 3 — Различные белки, захваченные вирусом из хозяйской клетки. 4 — Человеческие белки, обычно находящиеся на поверхности мембраны клеток. Захваченые в результате отпочковывания, они увеличивают вирулентность вируса. 5 — Внешняя оболочка ВИЧ, ведущая свое происхождение от цитоплазматической мембраны клетки, от которой когда-то отпочковался вирион. 6 — Матрикс, образованный тримерами белка р17. 7 — Тримерные комплексы белков gp120 и gp41, при помощи которых вирус проникает в клетку. На поверхности среднего вириона насчитывается около 20 таких тримеров. Чтобы увидеть рисунок в полном размере, нажмите на него.
. Все эти факты позволяют надеяться, что вскоре ВИЧ, как и оспа, останется в прошлом.
Читайте также: