
Афферентный рефлекторный путь вегетативной нервной системы
Изменения интенсивности деятельности внутренних органов могут быть вызваны раздражением как экстеро-, так и интерорецепторов, особенно собственных рецепторов органа. Афферентные пути от внутренних органов идут в ЦНС в составе вегетативных (симпатических и парасимпатических) и соматических нервов. При этом большинство афферентных волокон связывает орган с центрами спинного и стволом мозга по сегментарному принципу. Значительная часть афферентной импульсации от одного и того же органа поступает в ЦНС по нескольким нервным стволам в различные отделы ЦНС. Такой принцип афферентной иннервации внутренних органов обеспечивает более надежную их регуляцию. При повреждении отдельных афферентных путей оставщиеся афференты могут компенсировать роль утраченных проводников.
Периферические чувствительные пути ВНС. Симпатические стволы на 50% состоят из афферентных А-, В- и С-волокон. Однако большинство из них являются С-афферентами интра- и экстраорганных ганглиев. Часть этих афферентных волокон образует синапсы на вставочных клетках экстраорганных ганглиев, формируя афферентное звено периферической экстраорганной вегетативной рефлекторной дуги. Вставочный нейрон контактирует с эффекторным симпатическим нейроном этого же ганглия. Часть афферентных волокон симпатических нервов идет в спинной мозг в составе задних корешков.
Блуждающий нерв на 90% состоит из афферентных волокон групп А, В и С, подавляющее большинство которых — тонкие безмиелиновые С-волокна нейронов узловатого ганглия.
Тазовый нервна 50% состоит из афферентных волокон.
Во многих афферентах вегетативной и соматической нервной системы медиатором является глутамат, а кроме того, нейропептиды: ангиотензин II, вещество Р, ХЦК, энкефалин и др.
Вегетативные центры реагируют также на импульсацию чувствительных волокон соматической нервной системы, являющихся отростками афферентных нейронов межпозвоночных спинальных ганглиев и гомологичных им черепных узлов.
Сенсорные рецепторы ВНС (интерорецепторы) являются первичными рецепторами и локализуются в серозных оболочках и в стенках органов. Они воспринимают изменения внутренней среды организма, состояния внутренних органов, желез и сосудов. Это дает возможность ЦНС с помощью ВНС и эндокринной системы поддерживать гомеостазис. Хеморецепторы воспринимают изменения pH, осмотического давления, электролитного состава содержимого внутренних органов, Рсо2, Ро2, концентрации глюкозы, аминокислот. Механорецепторы реагируют на растяжение стенок органов, воспринимают давление: например, в артериях, в мочевом пузыре, в желудке и т.д. Терморецепторы располагаются в основном в ЖКТ, имеются в ЦНС. Болевые рецепторы имеются в тканях внутренних органов, мелких кровеносных и лимфатических сосудах.
Центральные афферентные пути ВНС. До ствола мозга афферентные пути ВНС несут информацию только от внутренних органов. В стволе мозга промежуточные нейроны являются полисенсорными — они реагируют на импульсацию от чувствительных соматических и вегетативных путей. В таламусе зоны представительства блуждающего и чревного нервов разделены, но в обеих зонах одни и те же нейроны возбуждаются от импульсов, поступающих по соматическим и вегетативным афферентным путям. Аналогичным образом реагируют и нейроны коры большого мозга. Это означает, что от таламуса в кору большого мозга поступает интегрированная информация. Специализированных зон в коре головного мозга, воспринимающих импульсацию только от внутренних органов, не обнаружено. ВП регистрируются в одних и тех же участках I и II соматосенсорных областей при раздражении экстеро- и интерорецепторов. В соматосенсорной зоне (постцентральная извилина) представительство чревного нерва совпадает с представительством нижних конечностей, блуждающего нерва — с представительством верхних конечностей. Корковые структуры вносят важный вклад в процесс интеграции соматической и висцеральной информации, что необходимо для согласования соматических и вегетативных компонентов при осуществлении поведенческих реакций организма для получения полезного приспособительного результата.
Афферентная импульсация от биологически активных точек, по-видимому, играет важную роль для человека. Таких точек насчитывают до 700. По некоторым данным, раздражение этих точек (акупунктура, пальцевой точечный массаж) может вызвать изменение работы тех или иных внутренних органов, снять головную боль, боль в области сердца и др. Свойства биологически активных точек, их физиологическая роль активно изучаются. Разработаны различные приборы для выявления и стимуляции этих точек с целью получения целебного эффекта.
Дата добавления: 2016-02-16 ; просмотров: 1951 ; ЗАКАЗАТЬ НАПИСАНИЕ РАБОТЫ
А. Общая характеристика. Вегетативные реакции могут быть вызваны раздражением как экстеро-, так и интерорецепторов, но легче всего они вызываются раздражением собственных рецепторов органа. Афферентные пути от внутренних органов идут в ЦНС в составе вегетативных (симпатических и парасимпатических) и соматических нервов. При этом большинство афферентных волокон связывает орган со спинным и продолговатым мозгом по сегментарному принципу. Однако значительная часть афферентов является надсегментарной — от одного и того же органа импульсы идут в ЦНС по нескольким нервным стволам в различные отделы ЦНС, начиная от спинного и кончая продолговатым мозгом (одни и те же отделы ЦНС посылают импульсы ко многим внутренним органам). Такой принцип афферентной иннервации внутренних органов обеспечивает более надежную их регуляцию. При повреждении отдельных афферентных путей оставшиеся афференты могут компенсировать роль утраченных проводников. Имеется и некоторая избирательность, например, афферентация от желудка осуществляется в большей степени по чревным нервам, афферентация от тонкой кишки — в основном по блуждающим нервам.
Б. Периферические афферентные пути
В составе блуждающего нерва содержит ся до 90 % афферентных волокон (группы А, В и С), подавляющее большинство которых — тонкие безмиелиновые С-волокна; тела ней ронов этих волокон находятся в узловатом ганглии.
В тазовом нерве афферентные волокна составляют 50 %.
В составе симпатических стволов также имеются А, В и С — афферентные волокна, в
совокупности они составляют 50 % от всего количества волокон. Их рецепторы локализуются в серозных оболочках и в стенках органов. Большинство афферентных волокон симпатических нервов также являются С-аф-ферентами периферического происхождения. Тела нейронов, от которых идут эти волокна, локализуются в интра- и экстраорганных превертебральных ганглиях. Часть этих афферентных волокон образует синапсы на вставочных клетках экстраорганных ганглиев, формируя афферентное звено периферической экстраорганной вегетативной рефлекторной дуги. Вставочный нейрон контактирует с эффекторным симпатическим нейроном этого же ганглия. Часть афферентных волокон симпатических нервов идет в спинной мозг в составе задних корешков, т.е. вместе с чувствительными соматическими волокнами.
4. Рецептивные поля внутренних органов снабжены также чувствительными волокнами, являющимися отростками афферентных нейронов межпозвоночных спинальных ганглиев и гомологичных им черепных узлов [Ленгли Д., 1903; Лаврентьев Б.И., 1943].
В. Сенсорные рецепторы афферентов ВНС воспринимают различные изменения внутренней среды, все они являются первичными рецепторами.
Механорецепторы воспринимают давле ние (например, в артериях, в мочевом пузы ре), реагируют на растяжение стенок органов.
Хеморецепторы воспринимают измене ния рН, электролитного состава содержимо го внутренних органов, РО2 и РСО2, кон центрации глюкозы и аминокислот, осмоти ческого давления.
Терморецепторы реагируют на измене ния температуры, располагаются в основном в желудочно-кишечном тракте.
Болевые рецепторы реагируют на ноци- цептивные воздействия. Наличие специали зированных болевых рецепторов лишь допус кается; предполагается, что болевые ощуще ния возникают при чрезмерном раздражении любых висцерорецепторов.
Полимодальные рецепторы (желудочно- кишечного тракта) одновременно реагируют на несколько воздействий.
Г. Центральные афферентные пути ВНС. Часть афферентных волокон ВНС, вступив в спинной мозг, образует синапсы с промежуточными нейронами данного сегмента, соседних или расположенных выше сегментов. Промежуточные нейроны образуют синапсы с преганглионарными нейронами, образующими эфферентную часть дуги вегетативного
рефлекса. Часть афферентных путей достигает продолговатого мозга, где находится второй афферентный нейрон, посылающий свой аксон в зрительный бугор к третьему афферентному нейрону. Аксон третьего афферентного нейрона направляется в соматосенсор-ную зону коры большого мозга. Восходящие афферентные волокна дают большое количество коллатералей к нейронам ретикулярной формации ствола мозга. В стволе мозга промежуточные нейроны являются полимодальными — они реагируют на импульсацию от чувствительных соматических и вегетативных путей. В таламусе зоны представительства блуждающего и чревного нервов разделены, но в обеих зонах одни и те же нейроны возбуждаются от импульсов, поступающих по соматическим и вегетативным афферентным путям. Аналогичным образом реагируют И нейроны коры большого мозга. Это означает, что от таламуса в кору большого мозга поступает интегрированная информация. Специализированных зон в коре головного мозга, воспринимающих импульсацию от внутренних органов, не существует. Вызванные потенциалы регистрируются в одних и тех же участках I и II соматосенсорных областей при раздражении экстеро- и интерорецепто-ров. В соматосзнсорной зоне (постцентральная извилина) представительство чревного нерва совпадает с представительством нижних конечностей, блуждающего нерва — с представительством верхних конечностей. Корковые структуры вносят важный вклад в процесс интеграции соматической и висцеральной информации, что важно для согласования соматических и вегетативных компонентов при осуществлении поведенческих реакций организма.
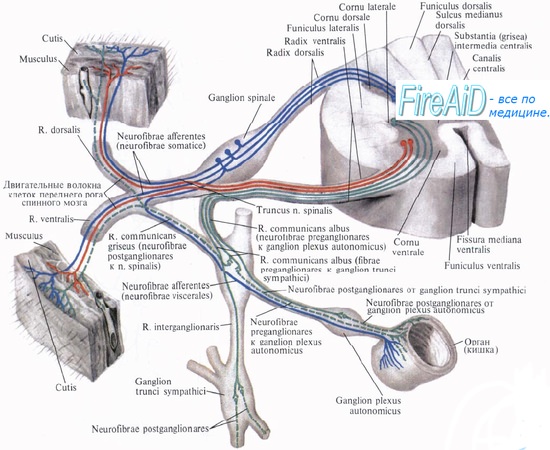
Рефлекторная дуга вегетативной нервной системы.
Клеточное тело воспринимающего нейрона как для анимальной, так и для вегетативной нервной системы помещается в спинномозговом узле, ganglion spinale, куда стекаются афферентные пути как от органов животной жизни, так и от органов растительной жизни и который, таким образом, является смешанным анимально-вегетативным узлом.
Клеточное тело вставочного нейрона вегетативной нервной системы в отличие от анимальной нервной системы помещается в боковых рогах спинного мозга. При этом аксон вставочного анимального нейрона, исходящий из клеток заднего рога, заканчивается в пределах спинного мозга среди клеток его передних рогов. Что же касается вставочного нейрона вегетативной нервной системы, то он в спинном мозге не заканчивается, а выходит за его пределы, к нервным узлам, расположенным на периферии.
Выйдя из спинного мозга, аксон вставочного нейрона подходит или к узлам симпатического ствола, ganglia trunci sympathici, относящимся к симпатическому отделу вегетативной нервной системы (они образуют симпатический ствол), или волокна не заканчиваются в этих узлах, а направляются к предпозвоночным узлам, расположенным более периферично, между симпатическим стволом и органом (ganglia coeliaca, ganglia mesenterica).
Кроме макроскопически видимых обособленных узлов, по ходу вегетативных нервов встречаются мигрировавшие сюда в ходе эмбрионального развития небольшие группы эффекторных нейронов — микроганглии. Все волокна, идущие до узлов первого, второго или третьего порядка и являющиеся аксонами промежуточного нейрона, называются предузловыми волокнами, rami preganglionares. Они покрыты миелином.

Третий, эффекторный, нейрон анимальной рефлекторной дуги помещается в передних рогах спинного мозга, а эффекторный нейрон вегетативной рефлекторной дуги вынесен в процессе развития из центральной нервной системы в периферическую, ближе к рабочему органу, и располагается в вегетативных нервных узлах. Из такого расположения эффекторных нейронов на периферии вытекает главный признак вегетативной нервной системы — двухнейронность эфферентного периферического пути: первый нейрон — вставочный; тело его лежит в вегетативных ядрах черепных нервов или боковых рогах спинного мозга, а нейрит идет к узлу; второй — эфферентный, тело которого лежит в узле, а нейрит достигает рабочего органа.
Эффекторные нейроны симпатических нервов начинаются в ganglia trunci sympathici (узлы первого порядка) или ganglia intermedia (узлы второго порядка), а для парасимпатических нервов — в около- или внутриорганных узлах, ganglia terminalia (третьего порядка); так как в названных узлах осуществляется связь вставочных и эфферентных нейронов, то отмеченная разница между симпатическим и парасимпатическим отделами вегетативной нервной системы связана именно с этими нейронами.
Аксоны эфферентных вегетативных нейронов почти лишены миелина — безмиелиновые (серые). Они составляют послеузловые волокна, rami postganglionics. Послеузловые волокна симпатической нервной системы, отходящие от узлов симпатического ствола, расходятся в двух направлениях. Одни волокна идут к внутренностям и составляют висцеральную часть симпатической системы. Другие волокна образуют rami communicantes grisei, соединяющие симпатический ствол с анимальными нервами.
В составе последних волокна достигают соматических органов (аппарата движения и кожи), в которых иннервируют непроизвольную мускулатуру сосудов и кожи, а также железы.
Совокупность описанных эфферентных вегетативных волокон, идущих от узлов симпатического ствола до органов сомы, составляет соматическую часть симпатического отдела. Такая структура обеспечивает функцию вегетативной нервной системы, которая регулирует обмен веществ всех частей организма применительно к непрерывно изменяющимся условиям среды и условиям функционирования (работы) тех или иных органов и тканей.
Соответственно этой наиболее универсальной своей функции, связанной не с какими-либо отдельными органами и системами, а со всеми частями, со всеми органами и тканями организма, вегетативная нервная система и морфологически характеризуется универсальным, повсеместным распространением в организме.
Следовательно, симпатический отдел иннервирует не только внутренности, но и сому, обеспечивая в ней обменные и трофические процессы.
В результате каждый орган, по И. П. Павлову, находится под тройным нервным контролем, в связи с чем он различает три вида нервов:
1) функциональные, осуществляющие функцию данного органа;
2) сосудодвигательные, обеспечивающие доставку крови к органу, и
3) трофические, регулирующие усвоение из доставленной крови питательных веществ.
Висцеральная часть симпатического отдела содержит все эти три вида нервов для внутренностей, а соматическая часть — только сосудодвигательные и трофические. Что же касается функциональных нервов для органов сомы (скелетная мускулатура и др.), то они идут в составе соматической, ани-мальной, нервной системы.
Таким образом, основное отличие эфферентной части вегетативной нервной системы от эфферентной части анимальной заключается в том, что анимальные, соматические, нервные волокна, выйдя из центральной нервной системы, идут до рабочего органа, нигде не прерываясь, тогда как вегетативные волокна на своем пути от мозга до рабочего органа прерываются в одном из узлов первого, второго или третьего порядка. Вследствие этого эфферентный путь вегетативной нервной системы разбивается на две части, из которых он и состоит: предузловые миелиновые волокна, rami preganglionares, и послеузловые, лишенные миелина (безмиелиновые) волокна, rami postganglionares.
Наличие узлов в эфферентной части рефлекторной дуги составляет характерный признак вегетативной нервной системы, отличающий ее от анимальной.

Лекция 11 . Физиология вегетативной нервной системы
Автономная (вегетативная) нервная система – это часть нервной системы, регулирующая работу внутренних органов, просвет сосудов, обмен веществ и энергии, обеспечивающих гомеостаз. Деятельность автономной нервной системы не подчиняется сознанию человека. Вегетативная нервная система делится на две части:
1. Симпатическую систему, задача которой состоит в мобилизации организма на решение жизненно важных задач;
2. Парасимпатическую систему, задача которой состоит в восстановлении нарушенного во время активности организма гомеостаза и восполнении потраченных ресурсов.
Эффекты этих двух систем часто противоположный друг другу. Большинство систем внутренних органов получают волокна от обеих систем ВНС. Обе системы работают согласованно. Кроме этого в ВНС выделяют интраорганную нервную систему, которая располагается непосредственно в органах человека и оказывает регулирующее воздействие на эти органы. Она регулирует моторику и сократительную активность.
Δ Общий план строения ВНС. Вегетативная нервная система состоит из четырех уровней организации: периферического, промежуточного, центрального и высшего.
Таблица 1. Уровни организации ВНС
Уровень организации
Локализация нервных центров
Основные функции
Интрамуральные сплетения во внутренних органах
Местные рефлекторные реакции на уровне отдельных органов или их участков
Паравертебральные и превертебральные ганглии
Периферические рефлекторные реакции; координация работы отдельных участков органа или органов одной системы
Краниобульбарный отдел продолговатого мозга, поясничный и крестцовый отделы спинного мозга
Запуск реакций мобилизации ресурсов в экстремальных состояниях (симпатическая нс) или восстановление потраченных ресурсов (парасимпатическая нс)
Гипоталамус, мозжечок, базальные ганглии, кора больших полушарий
Координация деятельности симпатической и парасимпатической нс; адаптация внутренних процессов, протекающих в организме, к требованиям окружающей среды
Δ Нейронный состав ВНС :
1. Чувствительные нейроны располагаются в спинномозговых узлах.
2. Преганглионарные нейроны располагаются в центральной нервной системе: в стволе мозга (парасимпатические) или в спинном мозге (в грудном отделе – симпатические, в крестцовом – парасимпатические). Их отростки – преганглионарные волокна идут до соответствующих вегетативных ганглиев (симпатические - до паравертебральных и превертебральных, парасимпатические – до интрамуральных), где они заканчиваются синапсами на постганлионарных нейронах.
3. Постганглионарные нейроны расположены в вегетативных ганглиях. Ганглии могут находиться или на некотором расстоянии от органов (симпатические ганглии), либо непосредственно в стенке иннервируемого органа (парасимпатические ганглии).
Δ Рефлекторный принцип работы ВНС. Дуга вегетативной нервной системы построена по общему принципу, но имеются особенности её организации:
1. Рефлекторная дуга ВНС может замыкаться вне ЦНС. Эффекторный нейрон для симпатического отдела ВНС расположен экстраорганно – в симпатических ганглиях, а для парасимпатической – внутри органа или непосредственно вблизи него.
2. Афферентное звено дуги вегетативного рефлекса может быть образовано как собственными – вегетативными, так и соматическими афферентными волокнами.
3. В дуге вегетативного рефлекса слабее выражена сегментарность, что повышает надежность вегетативной иннервации.
Вегетативные рефлексы подразделяются на следующие группы:
1. Периферические рефлексы , которые подразделяют на интраорганные, экстраорганные и центральные (разделение по уровню замыкания рефлекторной дуги).
2. Висцеро-соматические рефлексы – изменение соматической деятельности при возбуждении сенсорных рецепторов ВНС.
3. Висцеро-висцеральные рефлексы – рефлексы, рецептивные поля которых локализуются в одном органе или разных органах, при этом ответные реакции проявляются в изменениях активности других внутренних органов.
4. Сомато-висцеральные рефлексы – изменение деятельности внутренних органов при раздражении соматических рецепторов. Афферентное звено этих рефлексов относится к соматической нервной системе, а эфферентное – к ВНС.
5. Интероцептивные: механо-, хемо-, термо-, осмо-, ноцицепторы; проприоцептивные и экстероцептивные : термо - и ноцицепторы, рецепторы органов чувств (деление по рецепторам рефлексогенных зон).
ΔФункциональные отличия вегетативной нервной системы от соматической нервной системы :
1. Регуляция функций внутренних органов с помощью ВНС может осуществляться при полном нарушении связи с ЦНС, но менее совершенно, так как эффекторные нейроны ВНС находятся за пределами ЦНС.
2. Влияние ВНС на органы и ткани организма не контролируется непосредственно сознанием.
3. Нейроны ВНС имеют невысокую лабильность и небольшую скорость проведения импульсов.
4. Влияния симпатического отдела ВНС имеют генерализованный (диффузный) характер.
■ Физиология симпатической нервной системы . Центральные образования симпатической части ВНС расположены в ядрах боковых рогов серого вещества спинного мозга (с I по XII грудных сегментов и со II по IV поясничных сегментов). Симпатические нервные волокна выходят из спинного мозга в составе передних корешков, а затем отделяются от них. При этом выделяются две части этих волокон:
· Первая часть преганглионарных волокон прерывается в узлах симпатического ствола (паравертебральных ганглиях). Паравертебральные ганлии расположены в виде цепочек по обе стороны вдоль позвоночника от основания черепа до копчика. Здесь эта часть волокон прерывается и переключается на тела вторых нейронов. Симпатические ганглии входят в состав следующих вегетативных нервных сплетений:
1. Сердечное сплетение расположено у основания сердца; иннервирует сердце и легкие.
2. Чревное сплетение (солнечное) лежит позади желудка; иннервирует органы брюшной полости.
3. Подчревное сплетение находится впереди крестца; иннервирует органы малого таза.
· Вторая часть преганглионарных волокон идет через паравертебральные узлы не прерываясь. Эти волокна заканчиваются в превертебральных ганглиях.
Волокна, выходящие из паравертебральных узлов, называются постганглионарными. Постганглионарные волокна (после выхода из узла) разделяются: одни в составе нервов соматической нервной системы достигают скелетных мышц, другие идут к внутренним органам.
Δ Функции симпатической нервной системы . Симпатическая нервная система иннервирует почти все органы: сердце, сосуды, бронхи, желудочно-кишечный тракт, органы мочеполовой системы, потовые железы, печень, мышцы зрачка, матку, надпочечники и другие эндокринные железы. Её задача состоит в мобилизации организма на решение жизненно важных задач, для чего необходимо осуществить следующие физиологические процессы:
· Расширить кровеносные сосуды скелетной мускулатуры;
· Вовлечь в общий кровоток депонированную кровь, находящуюся в селезенке и подкожной клетчатке;
· Обеспечить более интенсивную работу сердца путем повышения частоты и силы сердечных сокращений, а также улучшения коронарного кровотока;
· Уменьшить доступ крови к органам, деятельность которых не нужна в данный момент;
· Извлечь резервные запасы питательных веществ и перевести их в доступную для организма форму;
· Усилить легочную вентиляцию для большего поступления кислорода в организм.
Комплекс данных реакций объединяют в понятие эрготропная функция симпатической нервной системы.
Δ Медиаторы симпатической нервной системы . В преганглионарных волокнах, которые контактируют с постганглионарным нейроном, выделяется ацетилхолин, то есть волокна и сам преганглионарный нейрон являются холинергическими. Ацетилхолин взаимодействует с Н-холинорецепторами (никотин-чувствительные рецепторы), в результате чего происходит передача возбуждения с преганглионарного волокна на постганглионарный нейрон. Никотин в малых концентрациях возбуждает Н-холинорецепторы, в больших – тормозит. Постганглионарные волокна симпатической нервной системы являются адренергическими (в их окончаниях выделяется норадреналин). Исключением являются постганглионарные симпатические волокна потовых желез, где выделяется ацетилхолин, поэтому эти волокна называются симпатическими холинергическими. Реакция органа на выделение медиатора зависит не столько от самого медиатора, сколь от типа рецептора, с которым данный медиатор взаимодействует. Конечный эффект возбуждения симпатических волокон зависит от того, какая популяция адренорецепторов (ά1,ά2,β1,β2) преобладает в органе на постсинаптической мембране. Механизм влияния норадреналина на клетку может быть различным: норадреналин после взаимодействия с адренорецептором меняет проницаемость мембраны для ионов, что приводит либо к деполяризации (увеличение потока входящих в клетку ионов натрия) - возбуждению, либо к гиперполяризации (повышение проницаемости для ионов калия) – торможению.
■ Физиология парасимпатической нервной системы . Центральные (преганглионарные) нейроны парасипатичекой нервной системы расположены в среднем, продолговатом мозге и в люмбально-сакральном отделе спинного мозга.
→ В среднем мозге расположены два парасимпатических ядра, относящихся к III паре черепно-мозговых нервов, которые обеспечивают иннервацию сфинктера зрачка и ресничную мышцу глаза.
→ В продолговатом мозге имеются парасимпатические ядра VII , IX , X пар черепно-мозговых нервов:
· Ядро VII пары чмн иннервирует слизистые железы носа, слезную железу, две слюнные железы (подъязычную и подчелюстную).
· Ядро IX пары чмн иннервирует околоушную железу.
· Ядро X пары чмн иннервирует сердце, легкие, желудочно-кишечный тракт.
→ В пояснично-сакральном отделе спинного мозга расположены нейроны, образующие центры мочеиспускания, дефекации и эрекции.
От преганглионарных нейронов аксоны образуют преганглионарные волокна, которые идут к органу. Парасимпатические ганглии расположены внутри органов, а в области головы и тазовых органов – в непосредственной близости от них. От нервных клеток ганглиев отходят постганглионарные волокна. По длине они короче, чем преганглионарные. Постганглионарные волокна заканчиваются либо на клетках органа, либо действуют на орган через интраорганную систему.
Функции парасимпатической нервной системы можно охарактеризовать следующим образом: возбуждение парасимпатической системы приводит к восстановлению гомеостаза, то есть к тропотрофному эффекту.
→ Медиаторы парасимпатической нервной системы . В преганглионарных волокнах парасимпатической нервной системы медиатором является ацетилхолин. Ацетилхолин взаимодействует на постсинаптической мембране постганглионарного нейрона с Н-холинорецепторами. В окончаниях постганглионарных волокон парасимпатической нервной системы также выделяется ацетилхолин, а рецепторы, расположенные на постсинаптической мембране органа – это М-холинорецепторы (мускаринчувствительные холинорецепторы).
■ Интраорганная нервная система . Многие внутренние органы, такие как сердце, мочеточники, желчный пузырь, кишечник, продолжают функционировать даже в изолированном от организма состоянии. Такую возможность им обеспечивает интраорганная нервная система – часть вегетативной нервной системы. Интраорганная нервная система имеет и другое название – метасимпатическая часть вегетативной нервной системы. Эта часть образована совокупностью микроганглионарных образований, находящихся в стенках внутренних органов (сердце, кишечник и т. д.). Данные образования обладают моторной активностью и имеют ряд особенностей:
1. Находятся только во внутренних органах, которые имеют свою собственную моторную активность;
2. Не вступают в антагонистические отношения с другими частями нервной системы;
3. Обладают независимостью от ЦНС;
4. Обладает холин-, адрен- и пуринергической видами передачи.
Рефлекторные дуги интраорганной нервной системы содержат в себе следующие нейроны: афферентные, вставочные и эффекторные. Чувствительные интрамуральные нейроны (афферентные) – клетки Догеля II типа образуют первое звено – рецептор, и второе – афферентный путь рефлекторной дуги. Двигательными являются клетки Догеля Iтипа (эфферентные), которые образуют эфферентный путь. Клетки интраорганной нервной системы могут быть механо-, хемо - и термочувствительными.
Интраорганная нервная система выполняет роль фактора надежности регуляторных механизмов организма.
Δ Медиаторы интраорганной нервной системы . Медиаторами интраорганной нервной системы являются более 20 видов химических веществ, к которым относят: серотонин, вазоактивный интестинальный пептид (ВИП), гистамин, дофамин, вещество Р и т. д.
■ Взаимодействие между отделами ВНС . Взаимодействие между отделами ВНС осуществляется следующими путями:
1. В основном синергизмом противоположных симпатических и парасимпатических влияний;
2. Взаимодействием в центральном отделе;
3. Взаимодействием в периферическом отделе.
4. Иногда однонаправленным синергичным влиянием на один и тот же орган;
■ Высшие центры ВНС . К высшим нервным центрам вегетативной нервной системы относят: гипоталамус, моноаминергическую систему, кору и мозжечок.
1. Гипоталамус является одним из важнейших образований мозга, участвующих в регуляции активности нейронов парасимпатической и симпатической нервной системы. Кроме того он является компонентом лимбической системы, в которую входят многие структуры старой и древней коры больших полушарий, в том числе гиппокамп и миндалевидный комплекс. В гипоталамусе имеются центры регуляции следующих функций:
· Центры регуляции водного и солевого обмена;
· Центры белкового, углеводного и липидного обмена;
· Центры регуляции сердечно-сосудистой системы, эндокринных желез;
· Центр голода и насыщения;
· Центры жажды и отказа от питья;
· Центр регуляции мочеотделения;
· Центр сна и бодрствования;
· Центр полового поведения;
· Центры, обеспечивающие эмоциональные переживания человека.
Гипоталамус представляет собой скопление более чем 32 пар ядер. Наиболее значимые из них – это
1. Супрооптическое и паравентрикулярные ядра – участвуют в процессах регуляции водно-солевого обмена, лактации, активности матки, способны продуцировать гормоны окситоцин, антидиуретический гормон;
2. Аркуатное и вентромедиальные ядра – вырабатывают пептидные гормоны либерины (высвобождающие другие гормоны) и статины (тормозящие выработку других гормонов);
2. Моноаминергическая система объединяет в себе скопление нейронов ствола, аксоны которых идут в составе медиального пучка переднего мозга и достигают практически всех структур мозга, в том числе гипоталамуса, таламуса, базальных ганглиев и некоторых участков коры. Моноаминергическая система работает с такими медиаторами как норадреналин, серотонин и дофамин.
→Тела норадренергических нейронов расположены в продолговатом мозге и в мосту. Особенно их много в голубом пятне. Эти нейроны оказывают тормозящее действие на структуры ЦНС.
→Тела дофаминергических нейронов лежат в среднем мозге, в основном черной субстанции. Аксоны их связаны с базальными ганглиями и другими структурами мозга.
→Серотонинергические нейроны локализованы в продолговатом мозге, в основном в ядрах шва, в мосту и в нижних отделах среднего мозга.
Норадреналин создает условия для появления у человека положительных эмоций – благодаря норадреналину в основном функционируют центры удовольствия.
Дофамин также способствует созданию положительных эмоций.
Серотонин играет роль в создании отрицательных эмоций – при этом активируются центры избегания.
Моноаминергическая система играет важную роль в создании эмоций и вегетативного обеспечения эмоционального состояния. Это осуществляется за счет воздействия нейронов моноаминергической системы на структуры лимбической системы и на ядра гипоталамуса, которые управляют вегетативными нейронами ствола и спинного мозга.
3. Мозжечок играет важную роль в регуляции вегетативных функций, так как влияет на возбудимость вегетативных центров и тем самым способствует адаптации организма к выполнению двигательных актов. То есть, мозжечок представляет собой посредника между вегетативной и соматической нервной системой. Мозжечок влияет на деятельность сердца, меняет величину артериального давления, регионарный кровоток, влияет на глубину и частоту дыхания, на моторную, секреторную, всасывательную функции желудочно-кишечного тракта, на тонус мышц мочевого пузыря. При повреждении мозжечка нарушается углеводный, белковый и минеральный обмен, нарушаются энергообразование и процессы терморегуляции.
4. Кора больших полушарий оказывает значительное влияние на вегетативную нервную систему. Особую роль играет гиппокамп: он задействован не только в процессах памяти и эмоциональных реакциях, но и в процессах регулирования вегетативных реакций. При раздражении гиппокампа происходит активация парасимпатического отдела автономной нервной системы, изменяется деятельность сердечно-сосудистой системы, меняются частота и глубина дыхания.
■ Тонус автономной нервной системы . Для оценки состояния тонуса ВНС применяется анализ вариабельности сердечного ритма, который не просто показывает общий тонус ВНС, а может избирательно вычленять, выделять вклад симпатического и парасимпатического отдела в общую картину. Интегральным показателем тонуса ВНС можно считать индекс напряжения (ИН) регуляторных систем по , отражающий соотношение активности симпатического и парасимпатического отделов ВНС. Тонус ВНС делят на пять типов:
1. Ваготонический – до 30 ед;
2. Нормотонический – 31-120 ед;
3. Симпатикотонический – 121-300ед;
4. Сверхсимпатикотонический – 301-600ед;
5. Запредельный – более 600ед.
При стрессе тонус автономной нервной системы сдвигается в область симпатикотонии не только за счет усиления активности симпатической нервной системы, но и за счет существенного снижения активности блуждающего нерва. При этом ИН увеличивается в 2-3 раза, достигая 150-180 единиц вместо обычных 65-80 единиц. Однако, если человеку не удается адаптироваться к острому стрессу, то со временем наступает обратная картина – сдвиг равновесия в сторону парасимпатической системы, сопровождающийся запредельным торможением коры больших полушарий.
Читайте также: