
Что такое конвергенция в нервной системе
Часто стимулы, поступающие к нервному пулу по одному волокну, возбуждают гораздо большее число нервных волокон, покидающих пул. Этот феномен называют дивергенцией. Существуют два основных типа дивергенции, функциональная роль которых различна.
На рисунке показан расходящийся тип дивергенции, при котором происходит распространение действия входящего сигнала на все большее количество нейронов по мере того, как сигнал проходит через последовательный ряд нейронов. Такой тип дивергенции характерен для кортикоспинального тракта, управляющего скелетными мышцами. При этом одна большая пирамидная клетка в двигательной области коры большого мозга при чрезвычайно облегченных условиях способна возбудить до 10000 мышечных волокон.
Второй тип дивергенции, приводит к расхождению нервных путей с образованием многочисленных трактов. В данном случае сигнал передается в двух направлениях от пула. Например, информация, передаваемая в восходящем направлении по задним столбам спинного мозга, в нижней части головного мозга направляется двумя разными путями: (1) в мозжечок; (2) через нижние области головного мозга к таламусу и коре большого мозга. Аналогично в таламусе почти вся сенсорная информация передается одновременно в еще более глубокие структуры самого таламуса и различные области коры большого мозга.
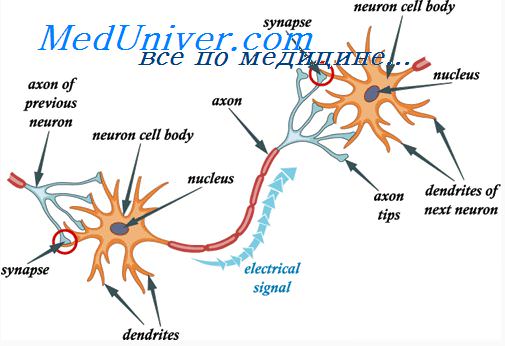
Конвергенция означает объединение сигналов множественных входов на одном нейроне. На рисунке схематически изображена конвергенция сигналов, исходящих из одного источника. Это значит, что на одном нейроне заканчиваются многочисленные терминали нервных волокон одиночного тракта. Этот тип конвергенции важен, поскольку нейроны почти никогда не возбуждаются потенциалом действия одной входящей терминали. Но потенциалы действия многих терминалей, конвергирующих на нейроне, обеспечивают достаточную пространственную суммацию, чтобы сдвинуть мембранный потенциал нейрона до порогового уровня, необходимого для его возбуждения.
Возможна также конвергенция сигналов (возбуждающих или тормозящих), исходящих из многих источников, как показано на рисунке. Например, на вставочных нейронах спинного мозга конвергируют сигналы от: (1) периферических нервных волокон, входящих в спинной мозг; (2) проприоспинальных волокон, идущих от одного сегмента спинного мозга к другому; (3) кортикоспинальных волокон из коры большого мозга; (4) нескольких других длинных нисходящих путей из головного в спинной мозг. Затем сигналы от вставочных нейронов сходятся на мотонейронах спинного мозга, непосредственно управляющих функцией скелетных мышц.
Такая конвергенция позволяет осуществлять суммацию информации из различных источников, а ответная реакция нейрона является результатом интеграции всей этой информации. Конвергенция — один из важных способов, с помощью которых центральная нервная система коррелирует, интегрирует и сортирует различные типы информации.
Иногда в ответ на сигнал, входящий в нервный пул, на выходе одновременно появляются возбуждающий сигнал, идущий в одном направлении, и тормозной сигнал, направляющийся по другому пути. Например, когда в спинном мозге одна группа нейронов посылает возбуждающий сигнал для движения ноги вперед, через другую группу нейронов передается сигнал, тормозящий мышцы, двигающие эту ногу назад, чтобы они не мешали движению вперед. Этот тип контура, называемый контуром с реципрокным торможением, характерен для всех нервных центров, управляющих мышцами-антагонистами.
На рисунке показан механизм развития такого торможения. Входящее волокно одновременно стимулирует возбуждающий выход пула (нейрон 1) и вставочный тормозной нейрон (нейрон 2), секретирующий медиатор, который тормозит второй выход пула. Этот тип контура важен также для предупреждения гиперактивности во многих частях мозга.
центре?
103.Что такое пролонгирование возбдужения в нервном центре?
Этим термином обозначается явление, при котором рефлекторный ответ сохраняется после прекращения действия раздражителя, т.е раздражение прекратилось, а, например, мышца продолжает сокращаться.
Существует 2 механизма этого феномена:
1. Следовая деполяризация мембраны нейрона, возникающая еще на фоне сохраняющегося ВПСП, создает условия для ритмической генерации потенциалов действия. Это кратковременное последствие.
2. Циркуляция возбуждения по замкнутым цепям нейронов в рефлекторном центре (принцип Лоренто-де-Но).
До прихода тормозного импульса или утомления одного из синапсов возбуждения может достаточно долго циркулировать по этим цепям. Бала выдвинута гипотеза о тем, что кратковременная память обусловлена реверберацией возбуждения в цепях положительной обратной связи.
104. Что такое феномен облегчения и каков его механизм?
Явление облегчения. Отдельные нейроны являются общими для каких-то двух рефлексов. При изолированной реализации одного из рефлексов эти нейроны не дают ответной реакции, так как раздражение для них оказывается подпороговым. При совместной реализации двух рефлексов их подпороговые эффекты суммируются и достигают пороговой величины. В результате суммации ответная реакция оказывается больше простой суммы изолированных эффектов.
Пул нейронов, расположенных в передних рогах спинного мозга состоит из высоковозбудимых (1,2) и низковозбудимых неронов (3). При раздражении только 1 афферентного нервного волокна в состояние возбуждения приходит нейрон 1, а в нейроне 3 возникает деполяризация, не достигающая критического уровня потенциала (т.е. его возбудимость увеличивается). При этом на выходе регистрируется сигнал равный 120 мВ.
При раздражении только II афферентного волокна возбуждается нейрон 2, а в нейроне 3 возникает увеличение возбудимости и на выходе регистрируется потенциал, например, равный 120 мВ. При одновременном раздражении I и II афферентных волокон в состояние возбуждения переходят нейроны 1,2,3. При данном воздействии регистрируется суммарный потенциал 320 мВ, т.е. не равный арифметической сумме отдельных потенциалов (120+120=240 мВ). Это обусловлено тем, что при одновременном раздражении волокон I и II дополнительно включается нейрон 3. Таким образом облегчение – это усиление поступающего сигнала.

временное
105. Что такое суммация возбуждения в нервном центре и ее механизм?
Особенностью ЦНС является то, что, как правило одни потенциал действия не вызывает возбуждение эффекторного нейрона. Только на ритмический раздражитель одного рецептора или одновременное раздражение нескольких нейронов возникает возбуждение эффекторного нейрона.
В зависимости от ситуации суммация бывает: 1) временная или 2) пространственная.
В случае временной суммации происходит суммирование квантов медиатора на постсинаптической мембране. Если в область постсинаптической мембраны ввести электрод и соединить его с усилителем постоянного тока и осциллографом, то можно наблюдать, что на одиночный раздражитель возбуждение нейрона не возникает, но на постсинаптической мембране возникает небольшой по амплитуде ВПСП.
С точки зрения возникновения возбуждения данный ВПСП является недостаточным, так как не может достигнуть критического уровня деполяризации. Если последующий импульс накладывается на первый, то есть суммируется и общий ВПСП достигает критического уровня. Это приводит к развитию потенциала действия. Таким образом 1) временная суммация – это суммация квантов медиатора, в первую очередь, в области аксонального холмика нейрона 2) временная суммация ВПСП обусловлена тем, что ВПСП продолжается дольше, чем рефрактерный период аксона, из окончания которого выделяется медиатор.
Пространственная суммация - это явление, когда раздельная стимуляция каждого из двух аксонов вызывает подпороговый ВПСП, однако при одновременной стимуляции обоих аксонов возникает потенциал действия, т.е. процесс, который не может быть обеспечен одиночным ВПСП.
Пространственная суммация может быть линейная и нелинейная. Если стимуляция будет пропорциональная каждому возбуждающему постсинаптическому потенциалу (ВПСП), то она называется линейной. Подобная ситуация возникает, когда возбуждающие синапсы на мембране одного нейрона удалены друг от друга на достаточном удалении.
Если синапсы находятся близко друг возле друга, силовые линии соседних ВПСП уменьшают их ВПСП и общий постсинаптический потенциал не будет пропорционален каждому ВПСП.
Пространственная суммация лежит в основе эффекта облегчения и окклюзии.
106. Что такое торможение? Классификация торможения.
Если на зрительный бугор поместить кристаллы соли (NaCl), то возникает торможение – удлинение времени рефлекса (замедление времени выдёргивания лапки из кислоты). Это наблюдение позволило И.М.Сеченову высказать мнение о явлении торможения в ЦНС. В последующем такой вид торможения получил название сеченовского торможения, или центральное торможение. Периферическое торможение было открыто ещё в 1845 году братьями Вебер (торможение деятельности сердца при раздражении блуждающих нервов).
Торможение – это активный процесс, связанный с возбуждением и ограничивающий его. Торможение это такой же врождённый процесс как и возбуждение, но общим для всех видов торможения является отсутствие способности к распространению по мембране нейрона и его отросткам. Различают два принципиально различных механизма торможения в ЦНС (рис. 20): первичное и вторичное. Первичное торможение развивается в клетках, примыкающих к тормозному нейрону. Оно инициировано возбуждением специальных тормозных нейронов, которые выделяют тормозные медиаторы. Вторичное торможение возникает в тех же нейронах, которые генерируют возбуждение.
107. Что такое первичное торможение?
Первичное торможение развивается в клетках, примыкающих к тормозному нейрону. Оно инициировано возбуждением специальных тормозных нейронов, которые выделяют тормозные медиаторы.
Первичное – возникает в специальных тормозных структурах под влиянием процесса возбуждения и проявляется подавлением другого возбуждения, развивающегося в соседних с этой структурой клетках. Следовательно, для тормозимой клетки этот процесс является первичным, т.е. без предварительного возбуждения.
| Постсинаптическое торможение (постторможение) | Пресинаптическое торможение |
| Этот вид торможения связан со специальными структурами (синапсами), которые выделяют тормозные медиаторы (ГАМК, Глицин) |
108. Что такое вторичное торможение и его виды?
Вторичное – это торможение возникает в той же самой клетке, в которой наблюдалось первичное возбуждение, т.е. торможение – это результат возбуждения.
Ко вторичному торможению относится пессимальное торможение и торможение вслед за возбуждением.
Пессимальное торможение – заключается в том, что в единицу времени к нейрону поступает очень большое количество потенциалов действия. В результате этого может происходить инактивация потенциал-зависимых натриевых каналов или десинситизация (потеря чувствительности) рецепторов, расположенных на постсинаптической структуре (десинситизация рецепторов обусловлена тем, что при большой частоте поступления медиатора он не успевает разрушаться и тем самым освобождать рецепторы для взаимодействия с новыми порциями медиатора).
Торможение вслед за возбуждением обусловлено тем, что в некоторых нейронах после потенциала действия развивается длительная следовая гиперполяризация, снижающая возбудимость клетки на долгое время.
Конвергенция нервных импульсов
Лат. converqere - сближать, сходиться - схождение к одному нейрону двух или нескольких возбуждений от сенсорных раздражителей (например звук, свет). Различают несколько видов конвергенции.
Конвергенция нервных импульсов сенсорно-биологическая - схождение к одному нейрону двух или нескольких возбуждений от сенсорных и биологических раздражителей одновременно (например, звук, голод, свет и жажда). Этот вид конвергенции является одним из механизмов обучения, образования условных рефлексов и афферентного синтеза функциональных систем.
Конвергенция нервных импульсов мультибиологическая - схождение к одному нейрону двух или нескольких возбуждений от биологических раздражителей например голод и боль, жажда и половое возбуждение).
Конвергенция нервных импульсов эфферентно-афферентная - схождение к одному нейрону двух или нескольких афферентных и эфферентных возбуждений одновременно. Эфферентное возбуждение отходит от нейрона, затем через несколько вставочных нейронов возвращается к нейрону и взаимодействует с афферентным возбуждением, приходящим к нейрону в этот момент. Этот вид конвергенции является одним из механизмов акцептора результата действия (предвидение будущего результата), когда афферентное возбуждение сличается с эфферентным.
Лат. diverqere - направляется в разные стороны - способность одиночного нейрона устанавливать многочисленные синаптические связи с различными нервными клетками. Благодаря процессу дивергенции одна и та же клетка может участвовать в организации различных реакций и контролировать большее число нейронов. В то же время каждый нейрон может обеспечивать широкое перераспределение импульсов, что приводит к иррадиации возбуждения.
Облегчение, проторение пути, банунг
Нем. bachnunq - проторение пути. После каждого, даже самого слабого раздражения, в нервном центре повышается возбудимость. При явлении суммации, когда в ЦНС идут два потока импульсов разделенных небольшим интервалом времени, то они вызывают значительно больший эффект, чем можно было ожидать в результате простого суммирования. Один поток импульсов как бы "проторяет путь” другому.
Лат. occlusum - закрывать, замыкать - взаимодействие двух потоков импульсов между собой. Впервые явление окклюзии было описано Ч. Шеррингтоном. Сущность его заключается во взаимном угнетении рефлекторных реакций, при котором суммарный результат оказывается значительно меньше, чем сумма взаимодействующих реакций. Согласно Ч.Шеррингтону, явление окклюзии объясняется перекрытием синаптических полей, образуемых афферентными звеньями взаимодействующих рефлексов. Поэтому при одновременном поступлении двух афферентных влияний возбуждающий постсинаптический потенциал вызывается каждым из них отчасти в одних и тех же мотонейронах спинного мозга.
Обмен веществ в нервных центрах
В нервных клетках, в противоположность нервному волокну, отмечается высокий уровень обмена веществ и чем больше дифференцирована нервная клетка, тем выше уровень обмена веществ. Если нервные клетки испытывают недостаток кислорода (например, при прекращении притока к ним крови), то через короткий срок они теряют способность возбуждаться и погибают. При деятельности нервных центров их обмен веществ возрастает. При рефлекторном возбуждении спинного мозга потребление кислорода увеличивается в 3-4 раза против уровня покоя. При этом также увеличивается потребление сахара, образование СО2. В нервных клетках или в окончаниях аксонов идет синтез медиаторов и ряда биологически активных нейропептидов, нейрогормонов и других веществ.
Утомляемость нервных центров - постепенное снижение и полное прекращение ответа при продолжительном раздражении афферентных нервных волокон. Утомление нервных центров вызывается прежде всего нарушением проведения возбуждения в межнейронных синапсах. Тот факт, что утомление сначала возникает в синапсе доказывается простым опытом. В то время, как раздражение афферентного нервного волокна спинальной лягушки не вызывает сокращения мышцы, стимуляция эфферентного волокна приводит к мышечной реакции.
В настоящее время считают, что утомление синапса обусловлено резким снижением запаса медиатора в пресинаптической мембране (истощение), уменьшением чувствительности постсинаптической мембраны (десенсетизация) и уменьшением энергетических ресурсов нейрона. Не все рефлекторные реакции одинаково быстро приводят к развитию утомления. Некоторые рефлексы могут в течение длительного времени протекать без развития утомления. К таким рефлексам относятся проприорецептивные тонические рефлексы.
Греч. tonos - натяжение, напряжение - состояние незначительного постоянного возбуждения, в котором обычно находятся все центры, имеющие рефлекторный характер. Тонус двигательных центров поддерживается непрерывным потоком импульсов от проприорецепторов, заложенных в мышцах. Слабое возбуждение от центров по центробежным волокнам передается мышцам, которые всегда находятся в несколько сокращенном состоянии (тонусе). Перерезка афферентных или эфферентных волокон приводит к потере мышечного тонуса.
Пластичность нервных центров - способность нервных элементов к перестройке функциональных свойств под влиянием длительных внешних воздействий или при очаговых повреждениях нервной ткани. Посттравматическая пластичность выполняет компенсаторную функцию. В экспериментах Флуранса (1827), П.К. Анохина (1935) доказано, что пластичностью обладают все нервные клетки, но наиболее сложные формы пластичности проявляются в корковых клетках. И.П.Павлов считал кору больших полушарий высшим регулятором пластических перестроек нервной деятельности. В настоящее время под пластичностью понимают изменение эффективности или направленности связей между нервными клетками.
Лат. dominantis - господствующий - временно господствующая рефлекторная система, обуславливающая интегральный характер функционирования нервных центров в какой-либо период времени и определяющая целесообразное поведение животного в конкретный, данный период времени. Доминантный нервный центр притягивает к себе возбуждение из других нервных центров и одновременно подавляет их деятельность, что приводит к блокаде реакций этих центров на те стимулы, которые ранее активировали их. Типичные черты доминанты проявляются в обнимательном рефлексе у самцов лягушек весной. Любое раздражение, например нанесение на лапку кислоты, приводит в таком состоянии к усилению обнимательного рефлекса.
Характерные черты доминанты: повышенная возбудимость, стойкость, способность к суммированию и инертность возбуждения, т.е. способность продолжать реакцию, когда первоначальный стимул уже миновал. Учение о доминанте разработано А.А.Ухтомским (1923). Доминанта является общим рабочим принципом центральной нервной системы..

1) вставочные нейроны, 2) эфферентный нейрон.
Конвергенция возбуждений лежит в основе облегчения рефлекторной реакции при одновременном раздражении нескольких афферентов.
Облегчение - это принцип координации рефлекторной деятельности, который характеризуется тем, что при одновременном раздражении двух афферентов возникает более сильная рефлекторная реакция, чем сумма рефлекторных ответов при раздельном возбуждении каждого из них.
Схема нейронной организации облегчения

1) афференты, 2) центральные зоны нервных центров, 3) периферические зоны нервных центров, 4) область перекрытия периферических зон.

1) афференты, 2) вставочные нейроны, 3) мотонейрон.
Окклюзия - это принцип координации рефлексов, который характеризуется тем, что при одновременном раздражении двух афферентов сила рефлекторной реакции меньше, чем сумма рефлекторных ответов при раздельном возбуждении каждого афферента.
Окклюзия возникает вследствие перекрытия центральных зон нервных центров. При одновременном раздражении двух афферентов активируется меньше центральных нейронов, чем при раздельной их стимуляции, так как часть нервных клеток локализуется в области перекрытия центральных зон и являются общими для обоих нервных центров.
Схема нейронной организации окклюзии
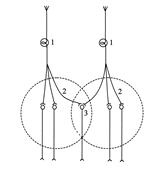
1) афференты, 2) центральные зоны нервных центров, 3) область перекрытия центральных зон.
Принцип реципрокного (сопряженного) торможенияхарактеризуется одновременным угнетением одного нервного центра при возбуждении другого.
Принцип реципрокности лежит в основе противоположных по функциональному назначению ответных реакций. Примером реципрокного торможения является взаимодействие нервных центров, обеспечивающих поочередное сокращение и расслабление мышц-сгибателей и разгибателей во время ходьбы, бега, плавания и т.п. Возбуждение, которое идет к мотонейронам, иннервирующим мышцы-сгибатели, по коллатерали афферента поступает к тормозным клеткам Реншоу. Эти клетки тормозят мотонейроны мышц-разгибателей. Если бы одновременно возбуждались нервные центры мышц сгибателей и разгибателей, движение конечности стало бы невозможным.
Схема нейронной организации реципрокного торможения жевательных мышц

1) собственно жевательная мышца, 2) мышца, опускающая нижнюю челюсть, 3) афферентные нейроны, 4) мотонейроны, 5) коллатераль, 6) тормозной нейрон.
Другим примером использования реципрокного торможения может быть стискивание зубов при сильной боли, чтобы подавить защитные движения, или прикусывание нижней губы, чтобы не смеяться при щекотании.
Нервные клетки обладают способностью регулировать частоту проходящих через них сигналов, а значит и силу рефлекторного ответа за счет обратной связи. Обратная связь базируется на способности коллатералей аксонов нервных клеток устанавливать синаптические контакты со вставочными нейронами, роль которых сводится к обратному воздействию на нейроны, образующие эти аксонные коллатерали.
Схема положительной обратной связи

1) эфферентный нейрон, 2) коллатераль, 3) возбуждающий вставочный нейрон.
Схема отрицательной обратной связи

1) эфферентный нейрон, 2) коллатераль, 3) тормозной вставочный нейрон.
Для деятельности нервной системы, как единого целого, в естественных условиях характерно наличие преобладающих (доминантных) очагов возбуждения, которые, по А.А. Ухтомскому (1923), являются проявлением одной из основных закономерностей работы нервных центров.
Доминанта - это господствующий очаг возбуждения в ЦНС, подчиняющий себе нервные центры в интересах выполнения наиболее важной в данный момент времени поведенческой деятельности организма.
Доминанту характеризуют пять основных свойств:
- повышенная возбудимость,
- способность к суммации,
- стойкость возбуждения,
- большая инертность,
- способность реципрокно тормозить деятельность других нервных центров (тех, которые не связаны с организацией наиболее важной в данный момент поведенческой деятельности).
Доминанта усиливается за счет конвергенции афферентных возбуждений от различных рефлексогенных зон. Так, очаг доминантного возбуждения, который возникает при острой зубной боли, поддерживается посторонними раздражителями (прикосновение, яркий свет, резкий звук), которые усиливают страдания больного.
На способности доминантного очага угнетать другие нервные центры основан метод звуковой аналгезии (аудиоаналгезия). Звуковой рардражитель – звук или даже простой шум, создает в ЦНС доминанту, которая реципрокно тормозит очаги возбуждения, возникающие при различных стоматологических манипуляциях. При определенных условиях обезболивающий эффект аудиоаналгезии во время препарировании твердых тканей зуба может достигать 90%.
Принцип доминанты лежит в основе организации целенаправленной поведенческой деятельности, но не определяет ее адекватности биологическим и социальным потребностям человека. Примером этого могут служить алкогольная и наркотическая доминанты, которые, подчиняя себе большинство нервных центров с целью удовлетворения потребности в алкоголе или наркотическом веществе, разрушительно отражаются на физическом, психическом и социальном благополучии человека.
Для осуществления сложных реакций необходима интеграция работы отдельных нервных центров. Большинство рефлексов являются сложными, последовательно и одновременно совершающимися реакциями. Рефлексы при нормальном состоянии организма строго упорядочены, так как имеются общие механизмы их координации. Возбуждения, возникающие в ЦНС, иррадиируют по ее центрам. Координация обеспечивается избирательным возбуждением одних центров и торможением других. Координация – это объединение рефлекторной деятельности ЦНС в единое целое, что обеспечивает реализацию всех функций организма.
Выделяют следующие основные принципы координации:
1. Принцип иррадиации возбуждений. Нейроны разных центров связаны между собой вставочными нейронами, поэтому импульсы, поступающие при сильном и длительном раздражении рецепторов, могут вызвать возбуждение не только нейронов центра данного рефлекса, но и других нейронов. Например, если раздражать у спинальной лягушки одну из задних лапок, слабо сдавливая ее пинцетом, то она сокращается (оборонительный рефлекс), если раздражение усилить, то происходит сокращение обеих задних лапок и даже передних. Иррадиация возбуждения обеспечивает при сильных и биологически значимых раздражениях включение в ответную реакцию большего количества мотонейронов.
3. Принцип доминанты. Был открыт А. А. Ухтомским, который обнаружил, что раздражение афферентного нерва (или коркового центра), обычно ведущего к сокращению мышц конечностей при переполнении у животного кишечника, вызывает акт дефекации. В данной ситуации рефлекторное возбуждение центра дефекации" подавляет, тормозит двигательные центры, а центр дефекации начинает реагировать на посторонние для него сигналы. А. А. Ухтомский считал, что в каждый данный момент жизни возникает определяющий (доминантный) очаг возбуждения, подчиняющий себе деятельность всей нервной системы и определяющий характер приспособительной реакции. К доминантному очагу конвергируют возбуждения из различных областей ЦНС, а способность других центров реагировать на сигналы, приходящие к ним, затормаживается. Благодаря этому создаются условия для формирования определенной реакции организма на раздражитель, имеющий наибольшее биологическое значение, т. е. удовлетворяющий жизненно важную потребность. В естественных условиях существования доминирующее возбуждение может охватывать целые системы рефлексов, в результате возникает пищевая, оборонительная, половая и другие формы деятельности.
Лат. converqere - сближать, сходиться - схождение к одному нейрону двух или нескольких возбуждений от сенсорных раздражителей (например звук, свет). Различают несколько видов конвергенции.
Конвергенция нервных импульсов сенсорно-биологическая - схождение к одному нейрону двух или нескольких возбуждений от сенсорных и биологических раздражителей одновременно (например, звук, голод, свет и жажда). Этот вид конвергенции является одним из механизмов обучения, образования условных рефлексов и афферентного синтеза функциональных систем.
Конвергенция нервных импульсов мультибиологическая - схождение к одному нейрону двух или нескольких возбуждений от биологических раздражителей например голод и боль, жажда и половое возбуждение).
Конвергенция нервных импульсов эфферентно-афферентная - схождение к одному нейрону двух или нескольких афферентных и эфферентных возбуждений одновременно. Эфферентное возбуждение отходит от нейрона, затем через несколько вставочных нейронов возвращается к нейрону и взаимодействует с афферентным возбуждением, приходящим к нейрону в этот момент. Этот вид конвергенции является одним из механизмов акцептора результата действия (предвидение будущего результата), когда афферентное возбуждение сличается с эфферентным.
Дивергенция возбуждения
Лат. diverqere - направляется в разные стороны - способность одиночного нейрона устанавливать многочисленные синаптические связи с различными нервными клетками. Благодаря процессу дивергенции одна и та же клетка может участвовать в организации различных реакций и контролировать большее число нейронов. В то же время каждый нейрон может обеспечивать широкое перераспределение импульсов, что приводит к иррадиации возбуждения.
Иррадиация(от лат. irradio — сияю, испускаю лучи) в физиологии, распространение процесса возбуждения или торможения в центральной нервной системе. Важную роль И. играет в деятельности коры больших полушарий головного мозга. И. возбуждения особенно отчётливо проявляется при сильном раздражении, когда в рефлекторный ответ вовлекаются нервные центры, обычно в нём не участвующие. Так, на умеренное болевое раздражение кожи стопы животное отвечает сгибанием лапы в голеностопном суставе; увеличение силы раздражения приводит к сгибанию ноги в коленном и тазобедренном суставах. При изучении действия тормозного условного раздражителя И. П. Павловым было показано, что торможение также может распространяться (иррадиировать) в клетках коры больших полушарий
Принцип иррадиации, или дивергенции, возбуждения ЦНС. Иррадиация возбуждения (от лат.irradio, озарять, освещать) - это распространение процесса возбуждения из одного участка ЦНС в другой. Каждый нейрон за счет многочисленных ответвлений (дивергенции), заканчивающихся синапсами, и большого числа вставочных нейронов связан со многими другими нейронами. Поэтому нервные импульсы от одного нейрона могут быть направлены к тысяче других нейронов. Процесс иррадиации возбуждения регулируется различными механизмами. Он может быть усилен, например, за счет активации ретикулярной формации ствола мозга. С другой стороны, процесс иррадиации ограничивается, и это важное действие осуществляется с участием многочисленных тормозных нейронов. Например, в спинном мозге ограничение иррадиации осуществляется благодаря механизму возвратного торможения с помощью специальных тормозных интернейронов - клеток Реншоу. Иррадиация возбуждения играет исключительно важную роль, так как позволяет обмениваться многочисленными потоками информации различным структурам мозга. Именно за счет иррадиации происходит обмен информацией между первой и второй сигнальными системами (элективная иррадиация), что существенно увеличивает возможности высшей нервной деятельности человека. Благодаря иррадиации возбуждения осуществляется интеграция деятельности правого и левого полушарий.
Принцип концентрации возбуждения
Возбуждение и торможение могут либо иррадиировать, либо концентрироваться. Концентрация - это явление, противоположное иррадиации. Оно возникает в тот момент, когда иррадиация достигает определенной границы, после чего распространение возбуждения или торможения идет в обратном направлении, т. е. в исходный пункт. Если нейроны находятся в заторможенном состоянии, это препятствует распространению на них процесса возбуждения, и наоборот, если они находятся в состоянии возбуждения, торможением они охватываются труднее. На скорость иррадиации и концентрации нервных процессов оказывает влияние и тип нервной системы, ее индивидуальные особенности.
Принцип индукции нервных процессов.Индукция отражает проявление процессов иррадиации возбуждения и торможения в коре больших полушарий. Принято считать, что индукция - (от лат.Inductio- введение, наведение) обозначает возникновение нервного процесса, противоположного по знаку процессу, вызванному условным раздражителем (положительным или отрицательным, т. е. тормозным). Одновременная индукция заключается в том, что формирование в каком-либо центре коры больших полушарий концентрированного возбуждения вызывает в прилежащих к этому центру зонах торможение (отрицательная одновременная индукция), а такое же концентрированное торможение вызывает в этих зонах возбуждение (положительная одновременная индукция). Таким образом, при одновременной индукции нервный процесс вызывает в другом участке коры процесс, противоположный по знаку (процесс возбуждения вызывает процесс торможения и наоборот), а при последовательной индукции происходит смена противоположных нервных процессов в одном и том же участке.
Принцип конвергенции возбуждения(или принцип общего конечного пути, воронка Шеррингтона). Конвергенция нервных импульсов (от лат.convergo,convergere- сближать, сходиться) означает схождение к одному нейрону двух или нескольких различных возбуждений одновременно. Это явление было открыто Ч. Шеррингтоном. Он показал, что одно и то же движение, например рефлекторное сгибание конечности в коленном суставе, можно вызвать путем раздражения различных рефлексогенных зон. В связи с этим им было введено понятие "общего конечного пути", или "принципа воронки", согласно которому потоки импульсов от различных нейронов могут сходиться на одном и том же нейроне (в данном случае - на альфа-мотонейронах спинного мозга). В частности, Ч. Шеррингтон обнаружил схождение к одним и тем же промежуточным или эфферентным нейронам различных афферентов от разных участков общего рецептивного поля (в спинном и продолговатом мозге) или даже от разных рецептивных полей (в высших отделах головного мозга). В настоящее время показано, что конвергенция возбуждения, так же как и дивергенция возбуждения, - очень распространенное явление в ЦНС. Основой для конвергенции (как и для иррадиации) является определенная морфологическая и функциональная структура различных отделов мозга. Очевидно, что часть конвергентных путей является врожденными, а другая часть (главным образом в коре большого мозга) - приобретенная в результате обучения в процессе онтогенеза. Формирование новых конвергентных отношений для нейронов коры большого мозга в процессе онтогенеза во многом связаны с формированием в коре доминантных очагов возбуждения, которые способны "притягивать" к себе возбуждение от других нейронов.
Принцип доминанты,или господствующего очага возбуждения. Доминанта (от лат.dominans,dominantis- господствующий) - это временно господствующая рефлекторная система, обусловливающая интегральный характер функционирования нервных центров в какой-либо период времени и определяющая целесообразное поведение животного и человека. В целом принцип доминанты означает, что текущая деятельность мозга определяется наличием господствующего (доминантного) очага возбуждения, или господствующего нейронного объединения, которое в данный момент времени подавляет и подчиняет себе деятельность остальных нейронных образований. Таким образом, благодаря формированию доминантного очага (доминантного нейронного объединения) деятельность мозга организуется таким образом, чтобы удовлетворить потребность организма, наличие которой и сформировало доминантный очаг возбуждения. Детальное изучение свойств доминантного очага показало, что для совокупности нейронов, входящих в его состав, характерны повышенная возбудимость, повышенная стойкость возбуждения, или инертность (заключающаяся в том, что у таких нейронов сложно вызвать торможение), повышенная способность к суммированию возбуждения, высокая способность "притягивать" возбуждение от других нервных центров (и тем самым повышать свою активность). Доминанта, как один из основных принципов координационной деятельности ЦНС, имеет важное значение в жизни человека. Например, именно благодаря доминанте возможно сосредоточение психической деятельности (внимание) и выполнение умственной или физической трудовой деятельности (в данном случае - это трудовая доминанта). В период поиска пищи и ее поедания реализуется пищевая доминанта. В настоящее время в отношении человека выделяют различные виды доминант (пищевую, оборонительную, половую, игровую, трудовую и др.).
Принцип субординации, или соподчинения, также относится к категории важнейших принципов организации работы мозга. Согласно этому принципу, деятельность нижележащих отделов мозга контролируется и управляется вышележащими отделами ЦНС. Например, в двигательных системах мозга и в вегетативной нервной системе имеются нейронные объединения (нервные центры), расположенные в спинном мозге или в стволе мозга, которые подчиняются деятельности нейронных объединений (нервных центров), находящихся в гипоталамусе, таламусе, мозжечке, базальных ядрах и коре больших полушарий.
Принцип обратной связи(обратной афферентации) и копий эфферентаций. Согласно этому принципу, для точной координации деятельности различных нейронных объединений (нервных центров, рефлекторных дуг) необходима оптимальная по объему информация о результатах действия. Она поступает в мозг по сенсорным каналам. Отсутствие такой информации приводит к дезинтеграции деятельности мозга. Особенно наглядна роль обратной афферентации при реализации двигательной активности - нарушение проприоцептивной чувствительности, как правило, препятствует выполнению точных движений, а также нарушает возможность формирования и сохранения адекватной для данного движения позы.
Принцип реципрокности(сопряжения) возбуждения и торможения на уровне спинного мозга реализуется с участием реципрокного торможения, благодаря которому возникают безусловные двигательные реципрокные рефлексы. Реципрокное торможение осуществляется по механизму постсинаптического торможения, которое возникает с участием специальных вставочных тормозных нейронов.
Принципы кодирования информациив нервной системе. В целом вся информация или значительная ее часть, передаваемая в ЦНС от одного отдела к другому, заключена в пространственном и временном распределении импульсных потоков, при этом используются различные нейронные коды. Выделяют три основные группы кодов.Неимпульсные сигналы, для которых характерны внутри- и внеклеточные факторы. К внутриклеточным факторам относятся амплитудные характеристики рецепторных и синаптических потенциалов, амплитудные и пространственные характеристики изменений синаптической проводимости, пространственное и временное распределение характеристик мембранного потенциала и градуальные потенциалы в аксонных терминалях. Внеклеточные факторы - это высвобождение медиаторов и ионов калия, нейросекреция, электротонические взаимодействия. Импульсные сигналы в одиночных нейронах. Дляимпульсных кодовглавными кандидатами являются кодыпространственные ("меченые линии", т. е. представление информации номером канала) ивременные - различные виды частотных или интервальных кодов (взвешенное среднее значение частоты, мгновенное значение частоты, частота разряда, форма интервальных гистограмм и т. д.). Выделяют такжемикроструктурное кодирование(временный узор импульсов), латентный код (момент появления или фазовые изменения разряда), числовой код (количество импульсов в пачке), код длинной пачки (длительность импульсации), наличие отдельного импульса или его отсутствие) изменение скорости распространения возбуждения в аксоне и пространственную последовательность явлений в аксоне. Ансамблевая активность (кодирование по ансамблю). В большинстве случаев в ЦНС используется пространственно-временное кодирование, когда информация о признаках сигнала передается канально и уточняется различными модификациями временных кодов.
2) Надпочечники (glandula suprarenalis, ед. ч.)-парные железы внутренней секреции, расположенные над верхними полюсами почек. У человека они находятся на уровне XI грудного - I поясничного позвонков , забрюшинно. Правый надпочечник имеет треугольную форму , левый - полулунную; вогнутые основания надпочечников примыкают к выпуклым полюсам почек. Вместе с почками надпочечники заключены в жировую капсулу ( сapsula adiposa ) и покрыты почечной фасцией ( fascia renalis ). Длина надпочечника взрослого человека варьирует от 30 до 70 мм, ширина - от 20 до 35 мм , толщина - от 3 до 10 мм, масса обоих надпочечников составляет 10 -14 г. Снаружи надпочечник покрыт соединительнотканной капсулой, от которой в паренхиму отходят перегородки, заключающие в себе сосуды и нервы и делящие паренхиму надпочечников на группы клеток и клеточные тяжи. В надпочечниках различают наружное корковое вещество, составляющее примерно 2/3 всей массы надпочечника , и внутреннее мозговое вещество. У новорожденных масса надпочечника в среднем 3,5 г. С возрастом она увеличивается, постепенно разграничиваются корковое и мозговое вещество, отдельные зоны коркового вещества.
Кровоснабжение надпочечников осуществляют три группы надпочечниковых артерий: верхняя, средняя, нижняя, проникающие в паренхиму в виде многочисленных капилляров, которые широко анастамозируют между собой и образуют в мозговом веществе синусоиды - расширения. Отток крови от надпочечников происходит через центральную и многочисленные поверхностные вены, впадающие в венозную сеть окружающих органов и тканей. Параллельно кровеносным расположены лимфатические капилляры, отводящие лимфу. Иннервируются надпочечники симпатическими (преимущественно) и парасимпатическими волокнами чревного, блуждающего и диафрагмального нервов . Корковое вещество надпочечника делится на клубочковую, пучковую и сетчатую зоны. Клубочковая зона, прилегающая тонким слоем к соединительнотканной капсуле, состоит из клеток неправильной формы. В пучковой, средней, наиболее широкой зоне группируются железистые клетки, расположенные радиальными колонками, а в сетчатой, внутренней, зоне группы клеток имеют вид неправильной сетки. Корковое вещество надпочечника богато липидами, окрашивающими его в желтый цвет. Гормоны коркового вещества кортикостероиды (глюко- и минералокортикоиды) синтезируются в митохондриях секреторных клеток из холестерина. Многообразное влияние кортикостероидов на все виды обмена веществ, сосудистый тонус, иммунитет и др. делает корковое вещество надпочечников важнейшим участком жизнеобеспечения человека в обычных условиях и в условиях адаптации к различным стрессам. В клубочковой зоне коркового вещества синтезируется альдостерон - основной минералокортикоид, участвующий в регуляции водно-солевого обмена. В пучковой зоне синтезируется преимущественно кортизол - глюкокортикоид, влияющий на белковый, жировой и углеводный обмен и на обмен нуклеиновых кислот, и кортикостерон, обладающий свойствами глюко- и минералокортикоида. В сетчатой зоне образуются половые гормоны, главным образом андрогены. Синтез кортикостероидов, прежде всего глюкокортикостероидов, регулируются адренокортикотропным гормоном. В глубине надпочечника находится мозговое вещество. Железистые клетки мозгового вещества получили название хромаффинных, или феохромных, т.к. избирательно окрашиваются солями хрома в желто-бурый цвет. Помимо железистых клеток в мозговом веществе надпочечника много нервных волокон и нервных клеток. Скопления хромаффинных нервных клеток, так называемых параганглиев, обнаруживают также по ходу легочного ствола и восходящей части аорты, в средостении находится поясничный аортальный параганглий и другие. Хромаффинные клетки секретируют три гормона (адреналин, норадреналин и дофамин), объединенных под общим названием катехоламины. Биосинтетическими предшественниками этих гормонов является аминокислота тирозин. Адреналин синтезируется только в надпочечниках; норадреналин и дофамин образуются также в параганглиях и многочисленных нейронах симпатической нервной системы. Все ткани, продуцирующие катехоламины, составляют адреналиновую систему. Активность хромаффинных клеток стимулируют различные воздействия окружающей и внутренней среды: эмоции, артериальная гипотензия, гипогликемия, физическая нагрузка, охлаждение и др.
Читайте также: