
Цитоплазма нервных клеток это
Цитоплазманейронов содержит обычные для всех клеток органеллы. В ней представлены эндоплазматическая сеть, рибосомы, митохондрии, комплекс Гольджи, клеточный центр, лизосомы, нейротубулы и нейрофиламенты (рис. 5).

Рис. 5. Схема ультраструктурной организации нервной клетки коры
головного мозга позвоночных (по И.Г. Павловой):
1 – плазмолемма; 2 – ядро; 3 – гранулярная эндоплазматическая сеть
(хроматофильная субстанция); 4 – комплекс Гольджи; 5 – лизосомы; 6 – митохондрии;
7 – нейрофиламенты; 8 – микротрубочки; 9 – дендрит; 10 – аксодендритические синапсы;
11 – аксосоматические синапсы
Наличие центросомы в настоящее время установлено в нейронах почти всех отделов нервной системы. Центросома лежит чаще всего около ядра нейрона, занимая всегда определенное положение в клетке. В нейробластах в период формирования нейрона центросома находится со стороны растущего отростка (аксона); в дифференцированных нейронах центросома – между дендритами и ядром.
Цитоплазма всех нервных клеток богата рибосомами, которые, как и в клетках других тканей, представлены гранулами диаметром 150–350 А. В нейробластах рибосомы распределяются в матриксе свободно поодиночке или образуют небольшие группы – полирибосомы. В дифференци-рованных нейронах значительная часть рибосом связана с поверхностью мембран цитоплазматической сети, которая соответствует эргастоплазме железистых или других клеток, продуцирующих белок.
В телах и крупных дендритах нервных клеток при световой микроскопии или окрашивании анилиновыми красителями хорошо видны глыбки базофильного вещества (хроматофильное вещество или субстанция Ниссля). При электронной микроскопии выявлено, что базофильное вещество представляет собой часть цитоплазмы, насыщенную уплощенными цистернами гранулярной эндоплазматического сети, содержащей многочисленные свободные и прикрепленные к мембранам рибосомы и полирибосомы. Обилие рРНК в рибосомах обусловливает базофильную окраску этой части цитоплазмы, видимую при световой микроскопии. Поэтому базофильное вещество отождествляют с гранулярной эндоплазматической сетью (рибосомами, содержащими рРНК). Оно никогда не содержится в нейрите и его конусовидном основании – аксонном холмике.
Размер глыбок базофильной зернистости и их распределение в нейронах разных типов различны. Так, в моторных клетках спинного мозга глыбки базофильного вещества крупные, неправильной угловатой формы; расположены они наиболее плотно вокруг ядра. Ближе к периферии тела клетки и в дендритах они обычно мельче, несколько вытянуты в длину и лежат реже. В чувствительных нейронах спинальных ганглиев глыбки имеют вид мелкой пылевидной зернистости. Базофильное вещество в клетках большинства узлов вегетативной нервной системы представлено мелкими зернами, расположенными в цитоплазме неравномерно, и образует нежную сеточку (узлы пограничного симпатического ствола, верхний шейный узел). В других ганглиях оно состоит из грубых глыбок, заполняющих все тело клетки (узлы солнечного сплетения, звездчатый узел) и ее дендриты.
В гранулярном эндоплазматическом ретикулуме в рибосомах, содержащих рРНК, непрерывно синтезируются новые белки цитоплазмы. К ним относятся белки, участвующие в построении и восстановлении клеточных мембран, метаболические ферменты, специфические белки, функционирующие в синаптическом проведении, и ферменты, инактивирующие этот процесс. Вновь синтезированные в цитоплазме нейрона белки поступают в аксон (а также в дендриты) для замещения израсходованных белков.
Морфология базофильного вещества изменяется в зависимости от функционального состояния клетки. При увеличении интенсивности специфической деятельности нейрона базофилия глыбок возрастает. В условиях перенапряжения или каких-либо травм (перерезка отростков, отравление, кислородное голодание, неадекватное раздражение) глыбки распадаются и исчезают. Этот процесс получил название хроматолиза (тигролиза),т.е. растворения базофильного вещества. Хроматолиз в разных случаях имеет свои специфические особенности, соответствующие характеру травмы. Это позволяет по морфологическим изменениям базофиль-ного вещества судить о состоянии нервных клеток в условиях патологии и эксперимента. Возвращение нейронов в нормальное состояние сопровождается восстановлением типичной для этих клеток картины базофильного вещества.
При регенерации аксона в теле нейрона наблюдается перемещение базофильного вещества по направлению к аксону, увеличивается количество гранулярного эндоплазматического ретикулума и числа митохондрий, усиливается белковый синтез и на проксимальном конце перерезанного аксона возможно появление отростков.
Пластинчатый комплекс(аппарат Гольджи) – система внутриклеточных мембран, каждая из которых представляет собой ряды уплощенных цистерн и секреторных пузырьков. Эту систему цитоплазматических мембран называют агранулярным ретикулумом ввиду отсутствия прикрепленных к ее цистернам и пузырькам рибосом. Пластинчатый комплекс принимает участие в транспорте из клетки определенных веществ, в частности белков и полисахаридов. Значительная часть белков, синтезированных в рибосомах на мембранах гранулярного эндоплазматического ретикулума, поступив в пластинчатый комплекс, превращается в гликопротеины, которые упаковываются в секреторные пузырьки, а затем выделяются во внеклеточную среду, это указывает на тесную связь между пластинчатым комплексом и мембранами гранулярного эндоплазматического ретикулума.
В цитоплазме фиксированных и обработанных солями серебра нервных клеток выявляется сеть тонких нитей – нейрофибриллы. В отростках нейронов нейрофибриллы располагаются параллельно друг другу. В теле нервной клетки они ориентированы неодинаково и в совокупности образуют густую связь. Нейрофибриллярный аппарат представляет собой морфологическое выражение правильной, линейной ориентации белковых молекул нейроплазмы. Изучение живых нефиксированных нервных клеток в культурах тканей, а также клеток, фиксированных при различных экспериментальных условиях, показало, что нейрофибриллярный аппарат – структура весьма подвижная и при невозможных функциональных состояниях выражена по-разному.
При электронной микроскопии в цитоплазме нервных клеток структуры, соответствующие микроскопически видимым нейрофибриллам, не обнаружены, но выявляются тонкие нити диаметром 60–100 А – нейрофиламенты (neurofilamenti) и трубочки – нейротубулы (neurotubuli) диаметром 200–300 А. Очевидно, они и представляют собой те комплексы белковых молекул, которые при агрегации и импрегнации азотнокислым серебром приобретают вид нейрофибрилл.
Нейрофиламенты можно выявить в большинстве крупных нейронов, где они располагаются в базофильном веществе, а также в миелинизированных аксонах и дендритах. Функции нейрофила-ментов еще до конца не выяснены. Роль нейротубул заключается в поддержании формы нейрона, особенно его отростков, и участии в аксоплазматическом транспорте веществ вдоль аксона.
Наряду с описанными выше нейронами имеются группы нервных клеток, например нейроны некоторых ядер гипоталамической области головного мозга, обладающие секреторной деятельностью. Нейросекреторные клетки имеют ряд специфических морфологических признаков. Это крупные нейроны. Цитоплазма их бедна базофильным веществом, оно преимущественно располагается по периферии тела клеток. В цитоплазме нейронов и в аксонах находятся различной величины гранулы и капли секрета, содержащие белок, а в некоторых случаях – липоиды и полисахариды. Гранулы нейросекрета нерастворимы в воде и спирте. Многие нейросекреторные клетки имеют ядра неправильной формы, что свидетельствует об их высокой функциональной активности.
Лизосомыпредставляют собой пузырьки, ограниченные простой мембраной и обеспечивающие фагоцитоз клетки. Они содержат набор гидролитических ферментов, способных гидролизовать вещества, попавшие в клетку. В случае гибели клетки лизосомальная мембрана разрывается и начинается процесс аутолиза: вышедшие в цитоплазму гидролазы расщепляют белки, нуклеиновые кислоты и полисахариды. Нормально функционирующая клетка надежно защищена лизосомальной мембраной от действия гидролаз, содержащихся в лизосомах.
Митохондрии –структуры, в которых локализованы ферменты окислительного фосфори-лирования. Они имеют внешнюю и внутреннюю мембраны, располагаются по всей цитоплазме нейрона, образуя скопления в концевых синаптических расширениях, они являются своеобразными энергетическими станциями клеток, в которых синтезируется аденозинтрифосфат (АТФ) – основной источник энергии в живом организме. Благодаря митохондриям в организме осуществляется процесс клеточного дыхания. Компоненты тканевой дыхательной цепи так же, как и система синтеза АТФ, локализованы во внутренней мембране митохондрий.
Митохондрии расположены как в теле нейрона, так и во всех его отростках. Особенно богата митохондриями цитоплазма нервных клеток в месте отхождения аксона и в концевых аппаратах отростков, в частности, цитоплазма структур межнейрональных синапсов. Митохондрии в нервных клетках при рассмотрении в световом микроскопе имеют форму палочек, нитей и зерен. По субмикроскопическому строению они существенно не отличаются от митохондрий других клеток.
В нервной клетке часто обнаруживаются пигментные гранулы. Темно-коричневые или черные гранулы меланина постоянно встречаются в нервной клетке некоторых отделов центральной нервной системы (средний мозг – черная субстанция, продолговатый мозг, дорсальное ядро блуждающего нерва, а также межпозвоночные и симпатические узлы). Значение этих пигментных образований пока неясно. Однако известно, что уменьшение числа пигментированных клеток в черной субстанции связано со снижением содержания дофамина в ее клетках и хвостатом ядре, что приводит к синдрому паркинсонизма. В нервной клетке часто встречаются также желтые гранулы липофусцина, являющегося продуктом нормальной жизнедеятельности нервной клетки. Количество липофусцина увеличивается с возрастом. Накопление жира в нервной клетке может являться результатом как нормального, так и патологического обмена. Гликоген обнаруживается во многих нервных клетках. Показано, что в цитоплазме ряда крупных нейронов, в особенности двигательных, содержится гликоген, количество которого меняется в зависимости от функционального состояния нервной клетки. По некоторым данным, гликоген связан с тигроидом; обнаружен также в нервных окончаниях в области синапсов.
Железосодержащие гранулы выявлены в нервной клетке черной субстанции, бледного шара и некоторых других образований центральной нервной системы. С возрастом количество железосодержащих гранул увеличивается. При прижизненном воздействии на нервные клетки основных или кислых красителей происходит, как и в других клетках организма, их отложение в виде гранул в области аппарата Гольджи, что можно рассматривать как защитное приспособление нервной клетки.
В цитоплазме аксона (аксоплазме) имеется много нитевидных митохондрий, аксоплазма-тических пузырьков, нейрофиламентов и нейротрубочек. Рибосомы в аксоплазме встречаются очень редко. Гранулярный эндоплазматический ретикулум отсутствует. Это приводит к тому, что тело нейрона снабжает аксон белками; поэтому гликопротеиды и ряд макромолекулярных веществ, а также некоторые органеллы, такие как митохондрии и различные пузырьки, должны перемещаться по аксону из тела клетки. Этот процесс называется аксонным, или аксоплазма-тическим транспортом. Определенные цитоплазматические белки и органоиды движутся вдоль аксона двумя потоками с различной скоростью. Один – медленный поток – со скоростью 1–3 мм/сут перемещает лизосомы и некоторые ферменты, необходимые для синтеза нейромедиаторов в окончаниях аксонов. Другой – быстрый, также направляется от тела клетки, но его скорость составляет 5–10 мм/ч (примерно в 100 раз выше скорости медленного потока). Этот поток транспортирует компоненты, необходимые для синаптической функции (гликопротеиды, фосфолипиды, митохондрии, дофамингидроксилаза для синтеза адреналина). Механизм аксоплазматического тока до конца не изучен.
Крупные дендриты отличаются от аксона также тем, что содержат рибосомы и цистерны гранулярного эндоплазматического ретикулума (базофильное вещество); здесь также много нейротрубочек, нейрофиламентов и митохондрий. Таким образом, дендриты имеют тот же набор органоидов, что и тело нервной клетки.
Цитоплазма нейронов содержит обычные для всех клеток органеллы. Пластинчатый комплекс в нервных клетках был впервые описан Гольджи в 1898 г. Наличие центросомы в настоящее время установлено в нейронах почти всех отделов нервной системы. Центросома лежит чаще всего около ядра нейрона, занимая всегда определенное положение в клетке. В нейробластах в период формирования нейрона центросома находится со стороны растущего отростка (аксона). В дифференцированных нейронах центросома лежит между дендритами и ядром. Митохондрии расположены как в теле нейрона, так и во всех его отростках. Особенно богата митохондриями цитоплазма нервных клеток в месте отхождения аксона и в концевых аппаратах отростков, в частности цитоплазма структур межнейрональных синапсов. Митохондрии в нервных клетках при рассмотрении в световом микроскопе имеют форму палочек, нитей и зерен. По субмикроскопическому строению они существенно не отличаются от митохондрий других клеток.
Цитоплазматическая сеть в дифференцированных нейронах представлена системой связанных между собой цистерн, пузырьков и канальцев. Их диаметр колеблется от 300 до 400 ?, а в отдельных случаях достигает 800-2000 ?. В совокупности они представляют трехмерную сеть двухконтурных мембран (альфа-цитомембран), ориентированных параллельно друг другу. Степень ориентации мембран в нейронах различных типов неодинакова. Максимально упорядоченно располагаются мембраны в нейронах спинного мозга. В целом цитоплазматическая сеть цитоплазмы нейронов - структура очень подвижная, изменяющаяся в соответствии с функциональным состоянием клетки.
Цитоплазма всех нервных клеток богата рибосомами, которые, как и в клетках других тканей, представлены гранулами диаметром 150-350 ?. В нейробластах рибосомы распределяются в матриксе свободно по одиночке или образуют небольшие группы - полирибосомы. В дифференцированных нейронах значительная часть рибосом связана с поверхностью мембран цитоплазматической сети, которая соответствует эргастоплазме железистых или других клеток, продуцирующих белок.
Рис. 3. Тигроидное вещество в корешковом нейроне спинного мозга (схема): 1 - аксон; 2 - дендрит
Базофильное вещество (substantia basophila), или хроматофильное вещество, тигроидное вещество, глыбки Ниссля, - участки цитоплазмы с большим содержанием рибосом, а, следовательно, и РНК, интенсивно окрашивается основными красителями. В соответствии с этим на препаратах, обработанных основными красками, или специфически на РНК, в перикарионе нейронов и их дендритах выявляется зернистость. Она образует в совокупности нерезко отграниченные базофильные глыбки, впервые описанные Нисслем (рис.3).
Базофильное вещество никогда не содержится в аксоне и в его конусовидном основании (аксонном холмике). Морфологии базофильного вещества различных типов нейронов присущ ряд особенностей.
Так, в моторных клетках спинного мозга глыбки базофильного вещества крупные, неправильной угловатой формы; расположены они наиболее плотно вокруг ядра. Ближе к периферии тела клетки и в дендритах они обычно мельче, несколько вытянуты в длину и лежат реже. В чувствительных нейронах спинальных ганглиев глыбки имеют вид мелкой пылевидной зернистости. Базофильное вещество в клетках большинства узлов вегетативной нервной системы представлено мелкими зернами, расположенными в цитоплазме неравномерно, и образует нежную сеточку (узлы пограничного симпатического ствола, верхний шейный узел). В других ганглиях базофильное вещество состоит из грубых глыбок заполняющих все тело клетки (узлы солнечного сплетения, звездчатый узел) и ее дендриты.
Морфология базофильного вещества изменяется в зависимости от функционального состояния клетки. При увеличении интенсивности специфической деятельности нейрона базофилия глыбок возрастает. В условиях перенапряжения или каких-либо травм (перерезка отростков, отравление, кислородное голодание, неадекватное раздражение) глыбки распадаются и исчезают. Этот процесс получил название хроматолиза (тигролиза), т.е. растворения базофильного вещества. Хроматолиз в разных случаях имеет свои специфические особенности, соответствующие характеру травмы. Это позволяет по морфологическим изменениям базофильного вещества судить о состоянии нервных клеток в условиях патологии и эксперимента. Возвращение нейронов в нормальное состояние сопровождается восстановлением типичной для этих клеток картины базофильного вещества.
Глыбки базофильного вещества нейронов представляют собой участки цитоплазмы, соответствующие гранулярной цитоплазматической сети других клеток. Так как РНК принимает активное участие в синтезе белковых веществ, можно считать, что глыбки базофильного вещества являются частью цитоплазмы, активно синтезирующей белок, необходимый для осуществления специфической функции нейрона.
При дифференцировке нейронов в период эмбрионального развития по мере роста отростков объем цитоплазмы резко увеличивается (в 2000 раз и более), при этом в соответствии с интенсивностью синтеза белка содержание РНК в них постепенно увеличивается и оформляется базофильное вещество. Наиболее заметные сдвиги в синтезе белка, накоплении РНК и формировании базофильного вещества наблюдаются в определенные периоды развития зародыша, совпадающие с усилением деятельности нервной системы. Например, с 7-х суток развития зародыша курицы обнаруживаются его рефлекторные движения, так как к этому времени оформляются рефлекторные дуги. Появление движений совпадает с увеличением концентрации РНК в моторных клетках спинного мозга и в чувствительных клетках спинальных ганглиев. В последующие дни моторная активность зародыша ослабевает, что сопровождается снижением количества РНК в нервных клетках. Затем двигательная активность зародыша усиливается с 19-20-х суток. В это время соответственно резко увеличивается и концентрация РНК, а также связанного с ней основного белка в нервных клетках. Базофильное вещество приобретает характерные для зрелой нервной клетки форму и химический состав.
Кроме гранулярного вида цитоплазматической сети, для цитоплазмы нервных клеток характерно наличие гладкой цитоплазматической сети в виде узких трубочек и пузырьков. В тесной связи с базофильным веществом в ряде нервных клеток, например в двигательных клетках, находятся включения гликогена, который образует с ними временные связи (симплексы). Помимо того, в цитоплазме нервных клеток всегда имеются различные ферменты: оксидаза, пероксидаза, фосфатаза, холинэстераза и др.
Пигментные включения нервных клеток представлены двумя видами пигмента. Меланин в виде черных, грубых, различной величины зерен находится только в определенных отделах нервной системы, а именно - в нейронах черного вещества и голубого места, а также дорсального ядра блуждающего нерва. Желтый пигмент липофусцин, содержащий липоиды, в виде мелкой зернистости встречается в нервных клетках всех отделов нервной системы. Появляется он у человека преимущественно после 7 лет и количество его увеличивается к 30 годам жизни.
Цитоплазма нейронов содержит обычные для всех клеток органеллы. Пластинчатый комплекс в нервных клетках был впервые описан Гольджи в 1898 г. Наличие центросомы в настоящее время установлено в нейронах почти всех отделов нервной системы. Центросома лежит чаще всего около ядра нейрона, занимая всегда определенное положение в клетке. В нейробластах в период формирования нейрона центросома находится со стороны растущего отростка (аксона). В дифференцированных нейронах центросома лежит между дендритами и ядром. Митохондрии расположены как в теле нейрона, так и во всех его отростках. Особенно богата митохондриями цитоплазма нервных клеток в месте отхождения аксона и в концевых аппаратах отростков, в частности цитоплазма структур межнейрональных синапсов. Митохондрии в нервных клетках при рассмотрении в световом микроскопе имеют форму палочек, нитей и зерен. По субмикроскопическому строению они существенно не отличаются от митохондрий других клеток.
Цитоплазматическая сеть в дифференцированных нейронах представлена системой связанных между собой цистерн, пузырьков и канальцев. Их диаметр колеблется от 300 до 400 Ǻ, а в отдельных случаях достигает 800-2000 Ǻ. В совокупности они представляют трехмерную сеть двухконтурных мембран (альфа-цитомембран), ориентированных параллельно друг другу. Степень ориентации мембран в нейронах различных типов неодинакова. Максимально упорядоченно располагаются мембраны в нейронах спинного мозга. В целом цитоплазматическая сеть цитоплазмы нейронов - структура очень подвижная, изменяющаяся в соответствии с функциональным состоянием клетки.
Цитоплазма всех нервных клеток богата рибосомами, которые, как и в клетках других тканей, представлены гранулами диаметром 150-350 Ǻ. В нейробластах рибосомы распределяются в матриксе свободно по одиночке или образуют небольшие группы - полирибосомы. В дифференцированных нейронах значительная часть рибосом связана с поверхностью мембран цитоплазматической сети, которая соответствует эргастоплазме железистых или других клеток, продуцирующих белок.
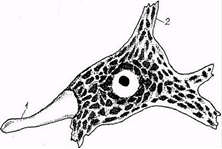
Рис. 3. Тигроидное вещество в корешковом нейроне спинного мозга (схема): 1 – аксон; 2 – дендрит
Базофильное вещество (substantia basophila), или хроматофильное вещество, тигроидное вещество, глыбки Ниссля, - участки цитоплазмы с большим содержанием рибосом, а, следовательно, и РНК, интенсивно окрашивается основными красителями. В соответствии с этим на препаратах, обработанных основными красками, или специфически на РНК, в перикарионе нейронов и их дендритах выявляется зернистость. Она образует в совокупности нерезко отграниченные базофильные глыбки, впервые описанные Нисслем (рис.3).
Базофильное вещество никогда не содержится в аксоне и в его конусовидном основании (аксонном холмике). Морфологии базофильного вещества различных типов нейронов присущ ряд особенностей.
Так, в моторных клетках спинного мозга глыбки базофильного вещества крупные, неправильной угловатой формы; расположены они наиболее плотно вокруг ядра. Ближе к периферии тела клетки и в дендритах они обычно мельче, несколько вытянуты в длину и лежат реже. В чувствительных нейронах спинальных ганглиев глыбки имеют вид мелкой пылевидной зернистости. Базофильное вещество в клетках большинства узлов вегетативной нервной системы представлено мелкими зернами, расположенными в цитоплазме неравномерно, и образует нежную сеточку (узлы пограничного симпатического ствола, верхний шейный узел). В других ганглиях базофильное вещество состоит из грубых глыбок заполняющих все тело клетки (узлы солнечного сплетения, звездчатый узел) и ее дендриты.
Морфология базофильного вещества изменяется в зависимости от функционального состояния клетки. При увеличении интенсивности специфической деятельности нейрона базофилия глыбок возрастает. В условиях перенапряжения или каких-либо травм (перерезка отростков, отравление, кислородное голодание, неадекватное раздражение) глыбки распадаются и исчезают. Этот процесс получил название хроматолиза (тигролиза), т.е. растворения базофильного вещества. Хроматолиз в разных случаях имеет свои специфические особенности, соответствующие характеру травмы. Это позволяет по морфологическим изменениям базофильного вещества судить о состоянии нервных клеток в условиях патологии и эксперимента. Возвращение нейронов в нормальное состояние сопровождается восстановлением типичной для этих клеток картины базофильного вещества.
Гигиеническое значение воды
"Вода, у тебя нет ни цвета, ни вкуса, ни запаха, тебя невозможно описать, тобой наслаждаются, не ведая, что ты такое. Нельзя сказать, что ты необходима для жизни: ты сама жизнь. Ты исполняешь нас радостью, которую не объяснишь н .
Клубочковая фильтрация
Фильтрация воды и низкомолекулярных компонентов из плазмы крови в полость капсулы происходит через клубочковый, или гломерулярный, фильтр. Гломерулярный фильтр имеет 3 слоя: эндотелиальные клетки капилляров, базальную мембрану и эпителий .
Субстраты
Основное правило — должны быть воздухо- и влагопроницаемые. Некоторые "рецепты": 1. все компоненты берутся в равных частях § легкая компостная земля § корни папоротника (или фибра древовидного папоротника) § мох сфагнум § .
Нервные клетки человека в подавляющем большинстве содержат одно ядро. Двухъядерные нейроны и тем более многоядерные встречаются крайне редко. Исключение составляют нервные клетки некоторых ганглиев вегетативной нервной системы, а именно - сплетения предстательной железы и узлов шейки матки. В этих нервных образованиях можно иногда наблюдать нейроны, содержащие до 15 ядер.
Форма ядра нервных клеток округлая. В ядрах содержится мало хроматина, что часто придает им на окрашенных препаратах пузырькообразный вид. Располагаются ядра обычно в центре тела нейрона, реже эксцентрично. Изучение ядер нервных клеток под электронным микроскопом показало, что они отграничены от цитоплазмы клетки двумя мембранами, расположенными друг от друга на расстоянии 200 ? и имеющими поры. В ядре нервных клеток имеется одно, а иногда 2 - 3 крупных ядрышка. Усиление функциональной активности нейронов обычно сопровождается увеличением объема и количества ядрышек. Ядра нервных клеток, особенно ядрышки, богаты РНК. Ряд авторов высказывает предположения, что в некоторых нейронах, характеризующихся высоким показателем ядерно-плазменного отношения (клетки-зерна мозжечка, ганглионарные клетки сетчатки и др.), значительная часть белков образуется в ядре, откуда поступает в цитоплазму и в отростки. ДНК ядра обычно мелко распылена, поэтому ядра крупных нейронов выглядят светлыми.
Цитоплазма нервной клетки
Цитоплазма нейронов содержит обычные для всех клеток органеллы. Пластинчатый комплекс в нервных клетках был впервые описан Гольджи в 1898 г. Наличие центросомы в настоящее время установлено в нейронах почти всех отделов нервной системы. Центросома лежит чаще всего около ядра нейрона, занимая всегда определенное положение в клетке. В нейробластах в период формирования нейрона центросома находится со стороны растущего отростка (аксона). В дифференцированных нейронах центросома лежит между дендритами и ядром. Митохондрии расположены как в теле нейрона, так и во всех его отростках. Особенно богата митохондриями цитоплазма нервных клеток в месте отхождения аксона и в концевых аппаратах отростков, в частности цитоплазма структур межнейрональных синапсов. Митохондрии в нервных клетках при рассмотрении в световом микроскопе имеют форму палочек, нитей и зерен. По субмикроскопическому строению они существенно не отличаются от митохондрий других клеток.
Цитоплазматическая сеть в дифференцированных нейронах представлена системой связанных между собой цистерн, пузырьков и канальцев. Их диаметр колеблется от 300 до 400 ?, а в отдельных случаях достигает 800-2000 ?. В совокупности они представляют трехмерную сеть двухконтурных мембран (альфа-цитомембран), ориентированных параллельно друг другу. Степень ориентации мембран в нейронах различных типов неодинакова. Максимально упорядоченно располагаются мембраны в нейронах спинного мозга. В целом цитоплазматическая сеть цитоплазмы нейронов - структура очень подвижная, изменяющаяся в соответствии с функциональным состоянием клетки.
Цитоплазма всех нервных клеток богата рибосомами, которые, как и в клетках других тканей, представлены гранулами диаметром 150-350 ?. В нейробластах рибосомы распределяются в матриксе свободно по одиночке или образуют небольшие группы - полирибосомы. В дифференцированных нейронах значительная часть рибосом связана с поверхностью мембран цитоплазматической сети, которая соответствует эргастоплазме железистых или других клеток, продуцирующих белок.

Рис. 3. Тигроидное вещество в корешковом нейроне спинного мозга (схема): 1 - аксон; 2 - дендрит
Базофильное вещество (substantia basophila), или хроматофильное вещество, тигроидное вещество, глыбки Ниссля, - участки цитоплазмы с большим содержанием рибосом, а, следовательно, и РНК, интенсивно окрашивается основными красителями. В соответствии с этим на препаратах, обработанных основными красками, или специфически на РНК, в перикарионе нейронов и их дендритах выявляется зернистость. Она образует в совокупности нерезко отграниченные базофильные глыбки, впервые описанные Нисслем (рис.3).
Базофильное вещество никогда не содержится в аксоне и в его конусовидном основании (аксонном холмике). Морфологии базофильного вещества различных типов нейронов присущ ряд особенностей.
Так, в моторных клетках спинного мозга глыбки базофильного вещества крупные, неправильной угловатой формы; расположены они наиболее плотно вокруг ядра. Ближе к периферии тела клетки и в дендритах они обычно мельче, несколько вытянуты в длину и лежат реже. В чувствительных нейронах спинальных ганглиев глыбки имеют вид мелкой пылевидной зернистости. Базофильное вещество в клетках большинства узлов вегетативной нервной системы представлено мелкими зернами, расположенными в цитоплазме неравномерно, и образует нежную сеточку (узлы пограничного симпатического ствола, верхний шейный узел). В других ганглиях базофильное вещество состоит из грубых глыбок заполняющих все тело клетки (узлы солнечного сплетения, звездчатый узел) и ее дендриты.
Морфология базофильного вещества изменяется в зависимости от функционального состояния клетки. При увеличении интенсивности специфической деятельности нейрона базофилия глыбок возрастает. В условиях перенапряжения или каких-либо травм (перерезка отростков, отравление, кислородное голодание, неадекватное раздражение) глыбки распадаются и исчезают. Этот процесс получил название хроматолиза (тигролиза), т.е. растворения базофильного вещества. Хроматолиз в разных случаях имеет свои специфические особенности, соответствующие характеру травмы. Это позволяет по морфологическим изменениям базофильного вещества судить о состоянии нервных клеток в условиях патологии и эксперимента. Возвращение нейронов в нормальное состояние сопровождается восстановлением типичной для этих клеток картины базофильного вещества.
Глыбки базофильного вещества нейронов представляют собой участки цитоплазмы, соответствующие гранулярной цитоплазматической сети других клеток. Так как РНК принимает активное участие в синтезе белковых веществ, можно считать, что глыбки базофильного вещества являются частью цитоплазмы, активно синтезирующей белок, необходимый для осуществления специфической функции нейрона.
При дифференцировке нейронов в период эмбрионального развития по мере роста отростков объем цитоплазмы резко увеличивается (в 2000 раз и более), при этом в соответствии с интенсивностью синтеза белка содержание РНК в них постепенно увеличивается и оформляется базофильное вещество. Наиболее заметные сдвиги в синтезе белка, накоплении РНК и формировании базофильного вещества наблюдаются в определенные периоды развития зародыша, совпадающие с усилением деятельности нервной системы. Например, с 7-х суток развития зародыша курицы обнаруживаются его рефлекторные движения, так как к этому времени оформляются рефлекторные дуги. Появление движений совпадает с увеличением концентрации РНК в моторных клетках спинного мозга и в чувствительных клетках спинальных ганглиев. В последующие дни моторная активность зародыша ослабевает, что сопровождается снижением количества РНК в нервных клетках. Затем двигательная активность зародыша усиливается с 19-20-х суток. В это время соответственно резко увеличивается и концентрация РНК, а также связанного с ней основного белка в нервных клетках. Базофильное вещество приобретает характерные для зрелой нервной клетки форму и химический состав.
Кроме гранулярного вида цитоплазматической сети, для цитоплазмы нервных клеток характерно наличие гладкой цитоплазматической сети в виде узких трубочек и пузырьков. В тесной связи с базофильным веществом в ряде нервных клеток, например в двигательных клетках, находятся включения гликогена, который образует с ними временные связи (симплексы). Помимо того, в цитоплазме нервных клеток всегда имеются различные ферменты: оксидаза, пероксидаза, фосфатаза, холинэстераза и др.
Пигментные включения нервных клеток представлены двумя видами пигмента. Меланин в виде черных, грубых, различной величины зерен находится только в определенных отделах нервной системы, а именно - в нейронах черного вещества и голубого места, а также дорсального ядра блуждающего нерва. Желтый пигмент липофусцин, содержащий липоиды, в виде мелкой зернистости встречается в нервных клетках всех отделов нервной системы. Появляется он у человека преимущественно после 7 лет и количество его увеличивается к 30 годам жизни.
Нейрофибриллы
В цитоплазме фиксированных и обработанных солями серебра нервных клеток выявляется сеть тонких нитей - нейрофибриллы (рис.4). В отростках нейронов нейрофибриллы располагаются параллельно друг другу. В теле нервной клетки они ориентированы различно и в совокупности образуют густую связь. Нейрофибриллярный аппарат представляет собой морфологическое выражение правильной, линейной ориентации белковых молекул нейроплазмы. Изучение живых нефиксированных нервных клеток в культурах тканей, а также клеток, фиксированных при различных эксперементальных условиях, показало, что Нейрофибриллярный аппарат - структура весьма подвижная и при различных функциональных состояниях выражена не одинаково.

Рис. 4. Нейрофибриллярный аппарат нейрона (схема)
При электронной микроскопии в цитоплазме нервных клеток структуры, соответствующие микроскопически видимым нейрофибриллам, не обнаружено, но выявляются тонкие нити диаметром 60-100 ? - нейрофиламенты и трубочки - нейротубулы диаметром 200-300 ?. Очевидно они и представляют собой те комплексы белковых молекул, которые при агрегации и импрегнации азотнокислым серебром приобретают вид нейрофибрилл.
Нейросекреторные клетки
Наряду с описанными выше нейронами имеются группы нервных клеток, например нейроны некоторых ядер гипоталамической области головного мозга, обладающие секреторной деятельностью. Нейросекреторные клетки имеют ряд специфических морфологических признаков. Это крупные нейроны. Цитоплазма их бедна базофильным веществом; оно преимущественно располагается по периферии тела клеток. В цитоплазме нейронов и в аксонах находятся различной величины гранулы и капли секрета, содержащие белок, а в некоторых случаях - липоиды и полисахариды. Гранулы нейросекрета нерастворимы в воде и спирте. Многие нейросекреторные клетки имеют ядра неправильной формы, что свидетельствует об их высокой функциональной активности.
Зеркальные нейроны
В настоящее время некоторые ученые выделяют зеркальные нейроны. Они обнаружены недавно и еще не признаны другими научными сотрудниками. Зеркальные нейроны на стадии изучения. Конкретных функций и свойств этих нейронов неизвестно, но ученые предполагают, что одна из их задач - это “сканирование" информации с этих нейронов (например: другого человека), вследствие чего мы понимаем его настроение, о чем он думает и т.д., глядя на него (это простейший пример). Факт гистогенеза и регенерации зеркальных нейронов еще не известен.
Читайте также: