
Физиология нервных и нервно-мышечных синапсов
Синапс – это специальная структура для передачи импульсов от клетки к клетке. Каждый синапс имеет пресинаптическую и постсинаптическую мембраны и синаптическую щель между ними. Пресинаптическая мембрана покрывает окончание аксона мотонейрона. Через нее в синаптическую щель поступает медиатор, который затем взаимодействует с рецепторами (их 20-30 млн в одном синапсе) постсинаптической мембраны – это мембрана подлежащей мышечной клетки. Механизм передачи возбуждения в синапсе состоит из 3-х этапов:
1 – ПД, возникший в пресинаптической мембране, повышает ее проницаемость для ионов кальция, который входит в нервное окончание и освобождает медиатор ацетилхолин из пузырьков.
2 – Ацетилхолин диффундирует в синаптическую щель, избыток медиатора разрушает фермент ацетилхолинэстераза.
3 – Ацетилхолин взаимодействует с холинорецепторами постсинаптической мембраны, в результате чего открываются натриевые каналы для входа натрия внутрь клетки – развивается потенциал действия, который распространяется вдоль мышечного волокна, вызывая его сокращение.
Нервно-мышечный синапс имеет следующие свойства: 1.одностороннее проведение возбуждения, 2.синаптическую задержку (на выход медиатора и его диффузию), 3.низкую лабильность (до 100 имп/сек), 4.быструю утомляемость, 5.высокую чувствительность к химическим веществам, лекарственным препаратам, токсинам (что может ускорять или тормозить передачу импульсов от нерва к мышце).
Движение как основное свойство живого.
Смеется ли ребенок при виде игрушки, улыбается ли Гарибальди, когда его гонят за излишнюю любовь к родине, создает ли Ньютон мировые законы и пишет их на бумаге – везде окончательным фактом является движение – так писал отец русской физиологии И.М.Сеченов.
14.Виды мышц, функции и свойства. Все мышцы организма делятся на: поперечно-полосатые (скелетные и сердечная) и гладкие, которые выстилают стенки внутренних органов, сосудов.
Поперечно-полосатые скелетные мышцы являются активной частью опорно-двигательного аппарата. В результате их сокращений происходит перемещение тела в пространстве, перемещение частей тела относительно друг друга, поддержание позы, произносятся слова, осуществляется дыхание, а также вырабатывается тепло.
Каждое мышечное волокно имеет следующие свойства:
1. Возбудимость – способность генерировать ПД в ответ на действие раздражителей.
2. Проводимость – способность проводить возбуждение в обе стороны от места нанесения раздражения.
3. Сократимость – способность сокращаться, изменять свою длину и напряжение.
4. Утомляемость. 5. Пластичность. 6. Тонус. 7.Рефрактерность.
Типы мышечных сокращений.
В зависимости от характера сокращений мышцы различают три их вида:
1.Ауксотоническое сокращение –когда одновременно изменяется длина и напряжение мышцы. Оно может быть эксцентрическим – когда напряжение мышцы сопровождается ее удлинением (например, в процессе приседания) и концентрическим – когда напряжением мышцы сопровождается ее укорочением (например, при разгибании нижних конечностей после приседания).
2.Изометрическое сокращение – когда напряжение мышцы возрастает, а длина ее не меняется. Этот вид сокращения можно наблюдать в эксперименте, когда оба конца мышцы зафиксированы (при фиксации определенного положения).
3.Изотоническое сокращение – укорочение мышцы при ее постоянном напряжении – сокращение ненагруженной мышцы.
В зависимости от длительности сокращений мышцы выделяют 2 их вида: одиночное и тетаническое.
1.Одиночное сокращение мышцывозникает при однократном раздражении нерва или самой мышцы.
2.Тетанические сокращения –это результат суммации двух и более одиночных сокращений. При полной суммации возникает гладкий тетанус, а при неполной суммации – зубчатый тетанус. При прекращении тетанического сокращения мышца расслабляется не полностью, а некоторое время находится в состоянии посттетанической контрактуры (напряжения).
Скелетные мышцы состоят из быстрых и медленных волокон, и соответственно – моторных единиц. Это – группа мышечных волокон, которые иннервируются одним нервным волокном, отходящим от мотонейрона. Быстрая мышца (мышцы глазного яблока) содержит больше моторных единиц, чем медленная (мышцы спины).
Вне действия раздражителя, т.е. в покое, скелетные мышцы находятся в состоянии некоторого напряжения, которое называют тонусом. Внешним выражением тонуса является упругость мышц.
Работа и сила мышц.
Степень укорочения мышцы зависит как от строения, так и от функционального состояния мышцы. Чем больше длина мышцы и ее поперечное сечение, тем больше она сокращается: это мышцы, имеющие перистое строение, - массетер. Сила мышцы – это максимальный груз, который она может поднять. Например, мышцы челюсти собаки поднимают груз, превышающий вес ее тела более 8 раз, а у человека – в 4 раза.
Работа мышц – произведение поднятого груза на величину укорочения мышцы. Если мышца сокращается без нагрузки, то ее работа равна 0. По мере увеличения нагрузки работа сначала увеличивается, а затем постепенно падает. Наибольшую работу мышца совершает при средних нагрузках, и это важно знать врачу при подборе тренировочных нагрузок для спортсмена. Работа мышц, связанная с перемещением груза, называется динамической. Статическая работа совершается при удержании груза в определенном положении, когда нарастает напряжение мышц. Статическая работа более утомительна, чем динамическая.
17.Утомление –это временное понижение работоспособности клетки, органа, организма, наступающее после работы и исчезающее после отдыха. Утомление мышц наступает по разным причинам.
1.Шиффсчитал, что утомление – это результат истощения запасов энергии в мышце.
2.Пфлюгер полагал, что накопление в мышце недоокисленных продуктов обмена во время работы приводит к развитию утомления.
3.Ферворн доказал, что утомление связано с нехваткой кислорода в мышцах.
Эти теории утомления мышц носят узколокалистический характер, так как в целостном организме (как доказал Сеченов в опыте с пильщиками дров) утомление первично возникает не в мышцах, а в нервных центрах. Заканчиваются запасы медиатора в синапсах ЦНС, требуется время на его синтез. Работоспособность мышцы восстанавливается.
Механизм сокращения мышц.
Мышечные волокна состоят из миофибрилл. Которые содержат до 2500 протофибрилл, представленные молекулами сократительных белков – актина и миозина. Изотропные участки состоят из тонких длинных нитей актина, а анизотропные – из толстых и коротких нитей миозина. Белок миозин имеет поперечные мостики с головками, где хранится АТФ. Сам белок обладает свойствами фермента АТФ-азы. На нитях актина имеются активные центры, содержащие белки тропонин и тропомиозин. Американские ученые- братья Хаксли считают, что при сокращении мышцы белковые нити не укорачиваются, а скользят друг по другу (теория скольжения нитей). Началом мышечного сокращения является выход ионов кальция из цистерн саркоплазматического ретикулума в межфибриллярное пространство. Кальций взаимодействует с белком тропонином, что приводит к смещению тропомиозина. В результате обнажаются активные центры актина, куда с помощью энергии АТФ прикрепляются головки миозиновых нитей. Происходит скольжение миозина и актина. Мышца укорачивается. Затем с помощью кальциевого насоса ионы кальция возвращаются в цистерны. Мышца расслабляется, так как актиновые и миозиновые нити принимают прежнее положение. Энергия АТФ необходима как для сокращения мышцы, так и для ее расслабления.
Синапс (греч. synapsis — соединение) — это специализированная структура, обеспечивающая передачу сигнала от клетки к клетке. Посредством синапса реализуется действие многих фармакологических препаратов.
Структурно-функциональная организация. Каждый синапс имеет пре- и постсинаптическую мембраны и синаптическую щель (рис. 17).
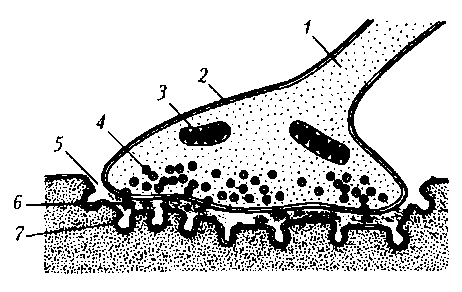
Рис. 17. Нервно-мышечный синапс скелетной мышцы: 1 – ветвь аксона; 2 – пресинаптическое окончание аксона; 3 – митохондрия; 4 – синаптические пузырьки, содержащие ацетилхолин; 5 – синаптическая щель; 6 – молекулы медиатора в синаптической щели; 7 – постсинаптическая мембрана мышечного волокна с N-холинорецепторами
Пресинаптическая мембрана нервно-мышечного синапса представляет собой часть мембраны пресинаптического окончания аксона мотонейрона. Через нее осуществляется выброс (экзоцитоз) медиатора (лат. mediator — посредник) в синаптическую щель. В нервно-мышечном синапсе медиатором является ацетилхолин. Медиатор пресинаптического окончания содержится в синаптических пузырьках (везикулах), диаметр которых составляет около 40 нм. Они образуются в комплексе Гольджи, с помощью быстрого аксонного транспорта доставляются в пресинаптическое окончание, где заполняются медиатором и АТФ. В пресинаптическом окончании содержится несколько тысяч везикул, в каждой из которых имеется от 1 тыс. до 10 тыс. молекул химического вещества.
Постсинаптическая мембрана (концевая пластинка в нервно-мышечном синапсе) — это часть клеточной мембраны иннервируемой мышечной клетки, содержащая рецепторы, способные связывать молекулы ацетилхолина. Особенность этой мембраны: множества мелких складок, увеличивающих ее площадь и количество рецепторов на ней до 10—20 млн в одном синапсе.
Синаптическая щель в нервно-мышечном синапсе имеет ширину в среднем 50 нм. Она содержит межклеточную жидкость, ацетилхолинэстеразу и мукополисахаридное плотное вещество в виде полосок, мостиков, в совокупности образующих базальную мембрану, соединяющую пре- и постсинаптическую мембраны.
Механизмы синаптической передачи включают три основных этапа (рис. 18).
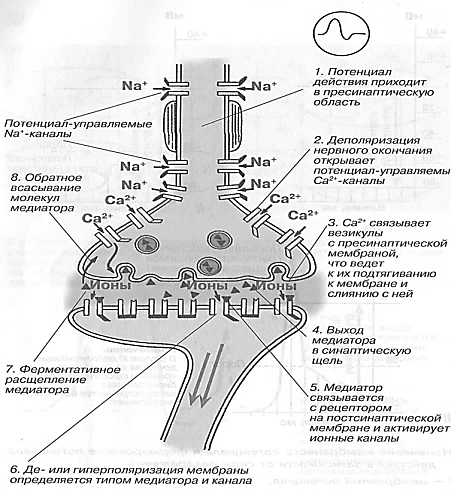
Рис. 18. Механизм проведения импульса через химический синапс: 1-8 – этапы процесса (Чеснокова, 2007)
Первый этап — процесс выброса медиатора в синаптическую щель, который запускается посредством ПД пресинаптического окончания. Деполяризация его мембраны ведет к открытию потенциалуправляемых Са-каналов. Са 2+ входит в нервное окончание согласно электрохимическому градиенту. Часть медиатора в пресинаптическом окончании локализуется на пресинаптической мембране изнутри. Са 2+ активирует экзоцитозный аппарат пресинапса, представляющий собой совокупность белков (синапсин, спектрин и др.), пресинаптического окончания, активация которых обеспечивает выброс ацетилхолина посредством экзоцитоза в синаптическую щель. Количество высвобождаемого ацетилхолина из пресинаптического окончания пропорционально в четвертой степени количеству поступившего туда Са 2+ . На один ПД из пресинаптического окончания нервно-мышечного синапса выбрасывается 200—300 квантов (везикул) медиатора.
Второй этап — диффузия ацетилхолина в течение 0,1—0,2 мс к постсинаптической мембране и действие его на N-холинорецепторы (стимулируются также никотином, вследствие чего и получили свое название). Удаление ацетилхолина из синаптической щели осуществляется путем разрушения его под действием ацетилхолинэстеразы, расположенной в базальной мембране синаптической щели, в течение нескольких десятых долей миллисекунды. Около 60% холина захватывается обратно пресинаптическим окончанием, что делает синтез медиатора более экономичным, часть ацетилхолина рассеивается. В промежутках между ПД из пресинаптического окончания происходит спонтанное выделение 1— 2 квантов медиатора в синаптическую щель в течение 1 с, формируя так называемые миниатюрные потенциалы (0,4—0,8 мВ). Они поддерживают высокую возбудимость иннервируемой клетки в условиях функционального покоя и выполняют трофическую роль, а в ЦНС — способствуют поддержанию тонуса ее центров.
Третий этап — взаимодействие ацетилхолина с N-холинорецепторами постсинаптической мембраны, в результате чего открываются ионные каналы на 1 мс и, вследствие преобладания входа N + в клетку, происходит деполяризация постсинаптичедкой мембраны (концевой пластинки). Эту деполяризацию в нервно-мышечном синапсе называют потенциалом концевой пластинки (ПКП) (рис. 19).
Особенностью нервно-мышечного синапса скелетного мышечного волокна является то, что при одиночной его активации формируется ПКП большой амплитуды (30—40 мВ), электрическое поле которого вызывает генерацию ПД на мембране мышечного волокна вблизи синапса. Большая амплитуда ПКП обусловлена тем, что нервные окончания делятся на многочисленные веточки, каждая из которых выбрасывает медиатор.
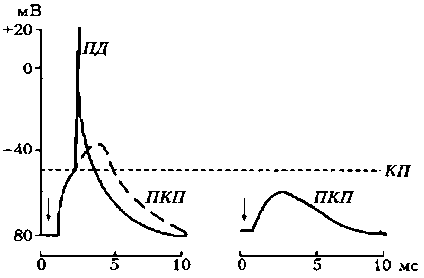
Рис. 19. Потенциал концевой пластинки (Шмидт, 1985): КП – критический потенциал; ПД – потенциал действия; А – ПКП в нормальной мышце; Б – ослабленный ПКП в курарезированной мышце; стрелками указан момент нанесения стимула
Характеристика проведения возбуждения в химических синапсах. Одностороннее проведение возбуждения от нервного волокна к нервной или эффекторной клетке, так как пресинаптическое окончание чувствительно только к нервному импульсу, а постсинаптическая мембрана — к медиатору.
Неизолированное — возбуждение рядом расположенных постсинаптических мембран суммируется.
Синаптическая задержка в передаче сигнала к другой клетке (в нервно-мышечном синапсе 0,5-1,0 мс), что связано с высвобождением медиатора из нервного окончания диффузией его к постсинаптической мембране и возникновением постсинаптических потенциалов, способных вызвать ПД.
Декрементность (затухание) возбуждения в химических синапсах при недостаточном выделении медиатора из пресинаптических окончаний в синаптические щели.
Низкая лабильность (в нервно-мышечном синапсе составляет 100 Гц), которая в 4 - 8 раз ниже лабильности нервного волокна. Это объясняется синаптической задержкой.
Проводимость нервно-мышечного синапса (как и химических синапсов ЦНС) угнетается или, наоборот, стимулируется различными веществами.
Например, кураре и курареподобные вещества (диплацин, тубокурарин) обратимо связываются с N-холинорецепторами постсинаптической мембраны, блокируют действие на нее ацетилхолина и передачу в синапсе. Напротив, некоторые фармакологические препараты, например прозерин, подавляют активность ацетилхолинэстеразы, способствуя умеренному накоплению ацетилхолина и облегчению синаптической передачи, что используется в лечебной практике.
Утомляемость (синаптическая депрессия) — ухудшение проводимости вплоть до полной блокады проведения возбуждения при длительном функционировании синапса (главная причина — истощение медиатора в пресинаптическом окончании).
Вопросы для самоконтроля
1.Каков механизм распространения возбуждения по нервному волокну? Какова роль перехватов Ранвье в проведении возбуждения по миелинизированному нервному волокну?
2.В чем преимущество скачкообразного (сальтаторного) распространения возбуждения над непрерывным его проведением вдоль мембраны волокна?
3.В чем физиологическое значение изолированного проведения возбуждения по нервному волокну?
4.Какие нервные волокна (афферентные или эфферентные, вегетативные или соматические) относятся к группе А? Какова скорость проведения возбуждения по ним?
5.Какие нервные волокна (афферентные или эфферентные, вегетативные или соматические) относятся к группе В? Какова скорость проведения по ним?
6.Какие нервные волокна (афферентные или эфферентные, вегетативные или соматические) относятся к группе С? Какова скорость проведения возбуждения по ним?
7.Перечислите структуры нервно-мышечного синапса (скелетная мышца). Что называют концевой пластинкой?
8.Перечислите последовательность процессов, ведущих к освобождению медиатора из пресинаптической мембраны в синаптическую щель при передаче возбуждения в синапсе.
9. Локальным потенциалом или распространяющимся возбуждением является потенциал концевой пластинки?
10.Что такое миниатюрные потенциалы концевой пластинки, каков механизм их возникновения?
11.В чем заключается трофическое влияние нерва на мышцу, осуществляемое через нервно-мышечный синапс?
12.Какие вещества являются медиаторами в нервно-мышечных синапсах гладкой и поперечнополосатой мышц?
13.Что такое сенсорный рецептор?
14.На какие две группы делятся сенсорные рецепторы по скорости адаптации? Назовите рецепторы, относящиеся к каждой из них.
15.Что понимают под первичными и вторичными рецепторами?
16.Перечислите основные свойства рецепторов.
17.Что называют адаптацией рецепторов? Как изменяется частота импульсов в афферентном нервном волокне при адаптации рецептора?
18.Назовите локальные потенциалы, возникающие при возбуждении первичных и вторичных рецепторов.
19.Рецепторный потенциал, где он возникает, каково его значение?
20.Генераторный потенциал, где он возникает, каково его значение?
21.Где возникает потенциал действия при возбуждении первичного сенсорного рецептора?
22. Где возникает потенциал действия при возбуждении вторичного сенсорного рецептора?
Физиология мышц
1.3.1. Структурнофункциональная характеристика скелетной мышцы
Мышцы подразделяют на поперечнополосатые (скелетная и сердечная) и гладкие (сосуды и внутренние органы, кроме сердца).
Скелетная мышца состоит из мышечных волокон, изолированных в структурном и функциональном отношении друг от Друга, которые представляют собой вытянутые многоядерные клетки. Толщина волокна составляет 10—100 мкм, а его длина варьирует в пределах от нескольких миллиметров до нескольких сантиметров. Количество мышечных волокон, установившись постоянным на 4—5-м месяце постнатального онтогенеза, в последующем не изменяется; с возрастом изменяются (увеличиваются) лишь их длина и диаметр.
Назначение основных структурных элементов. Характеристика основных элементов мышечного волокна. От клеточной мембраны мышечного волокна (сарколеммы) вглубь отходят многочисленные поперечные инвагинации (Т-трубочки), которые обеспечивают ее взаимодействие с саркоплазматическим ретикулулом (СПР) (рис. 20).
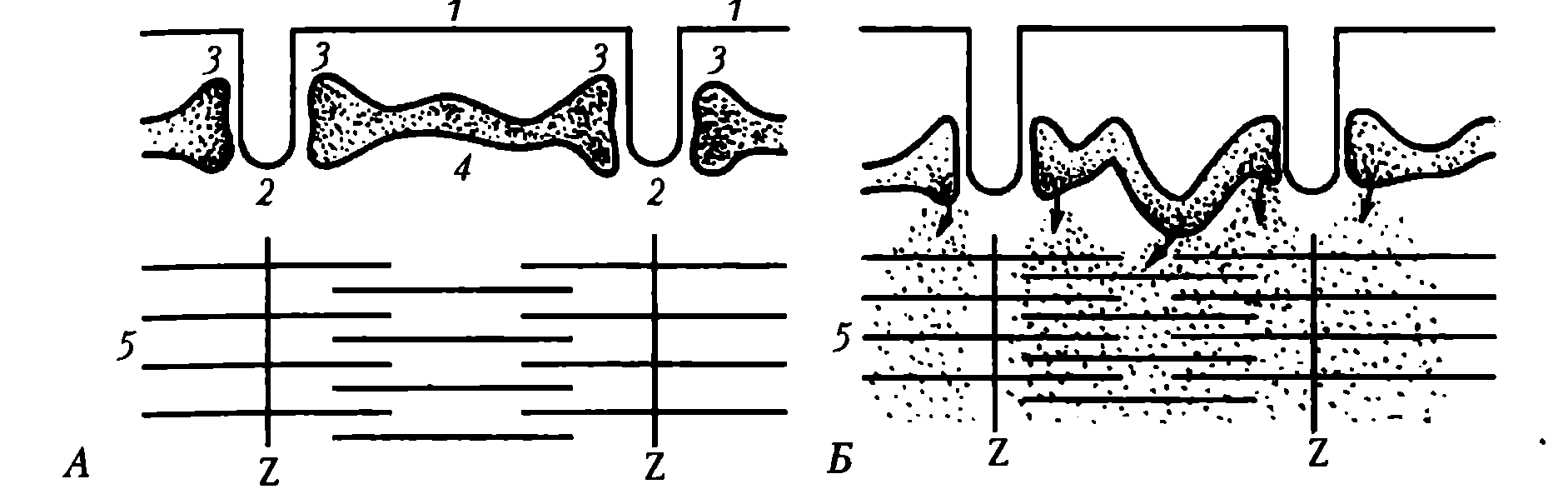
Рис. 20. Взаимоотношение клеточной мембраны (1), поперечных трубочек (2), боковых цистерн (3) и продольных трубочек (4) саркоплпзматическаого ретикулума, сократительных белков (5): А – в состоянии покоя; Б – при сокращении мышечного волокна; точками обозначены ионы Ca 2+
СПР представляет собой систему связанных друг с другом цистерн и отходящих от них в продольном направлении канальцев, расположенных между миофибриллами. Терминальные (концевые) цистерны СПР примыкают к Т-трубочкам, формируя так называемые триады. В цистернах содержится Са 2+ , играющий важную роль в мышечном сокращении. В саркоплазме имеются внутриклеточные элементы: ядра, митохондрии, белки (в том числе миоглобин), капельки жира, гранулы гликогена, фосфатсодержащие вещества, различные малые молекулы и электролиты.
Миоибриллы — субъединицы мышечного волокна. В одном мышечном волокне может насчитываться более 2 тыс. миофибрилл, их диаметр 1—2 мкм. В одиночной миофибрилле содержится 2—2,5 тыс. протофибрилл — параллельно расположенных нитей белка (тонкие — актин, толстые — миозин). Актиновые нити состоят из двух субъединиц, скрученных в виде спирали. В состав тонких нитей входят также регуляторные белки — тропомиозин и тропонин (рис. 21).
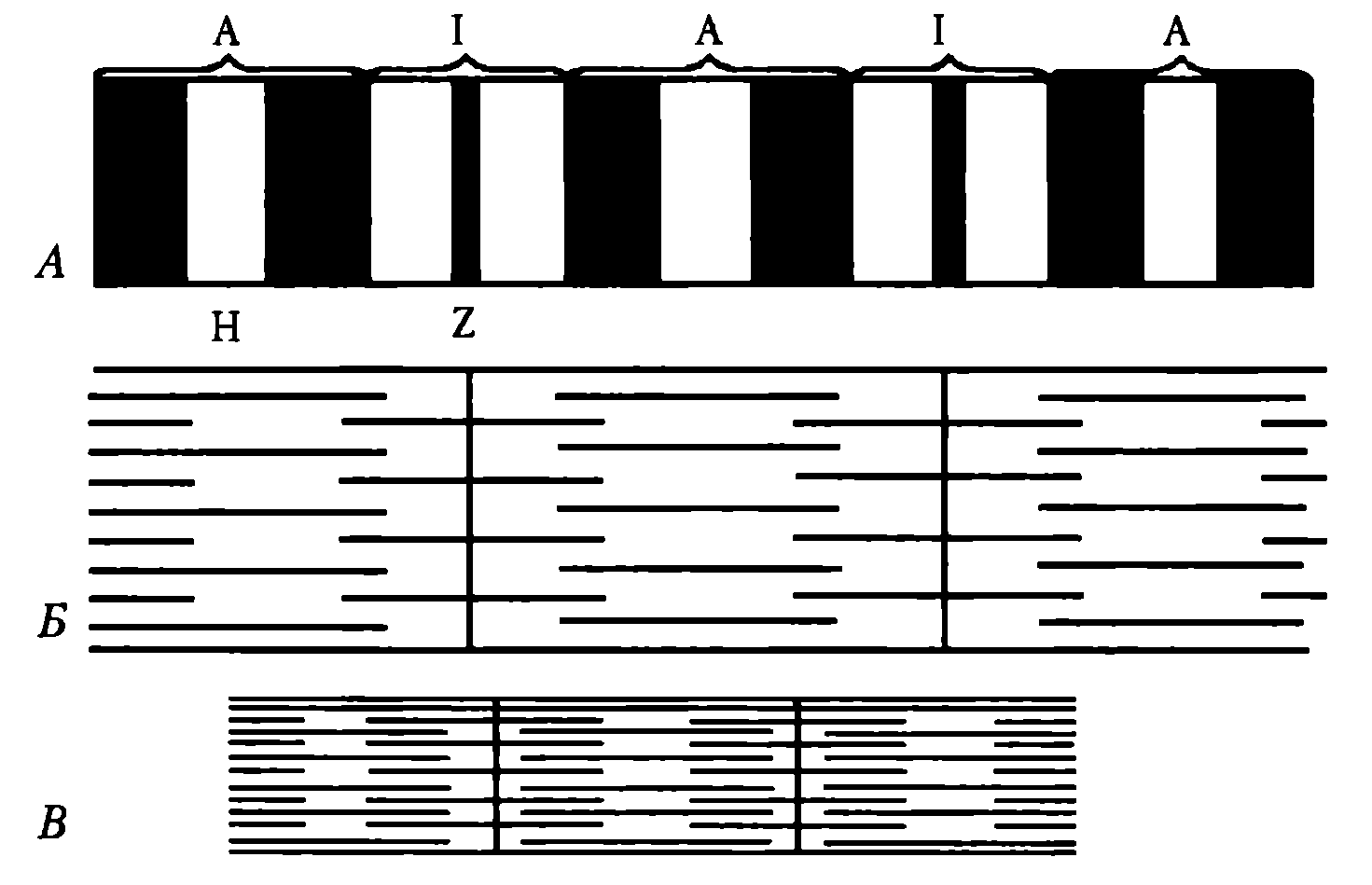
Рис. 21. Взаимное расположение структурных элементов миофибрилл при их расслаблении (А,Б) и сокращении (В)
Эти белки в невозбужденной мышце препятствуют взаимосвязи актина и миозина, поэтому мышца в покое находится в расслабленном состоянии. Миофибриллы включают в себя последовательно соединенные блоки — саркомеры (Б), отделенные друг от друга Z-полосками. Саркомер (длина 2—Змкм) является сократительной единицей мышечного волокна; при длине 5см оно включает в себя около 20 тыс. последовательно соединенных саркомеров. Миофибриллы отдельного мышечного волокна связаны таким образом, что расположение саркомеров совпадает, и это создает картину поперечной исчерченности волокна при наблюдении в световом микроскопе (рис. 22).
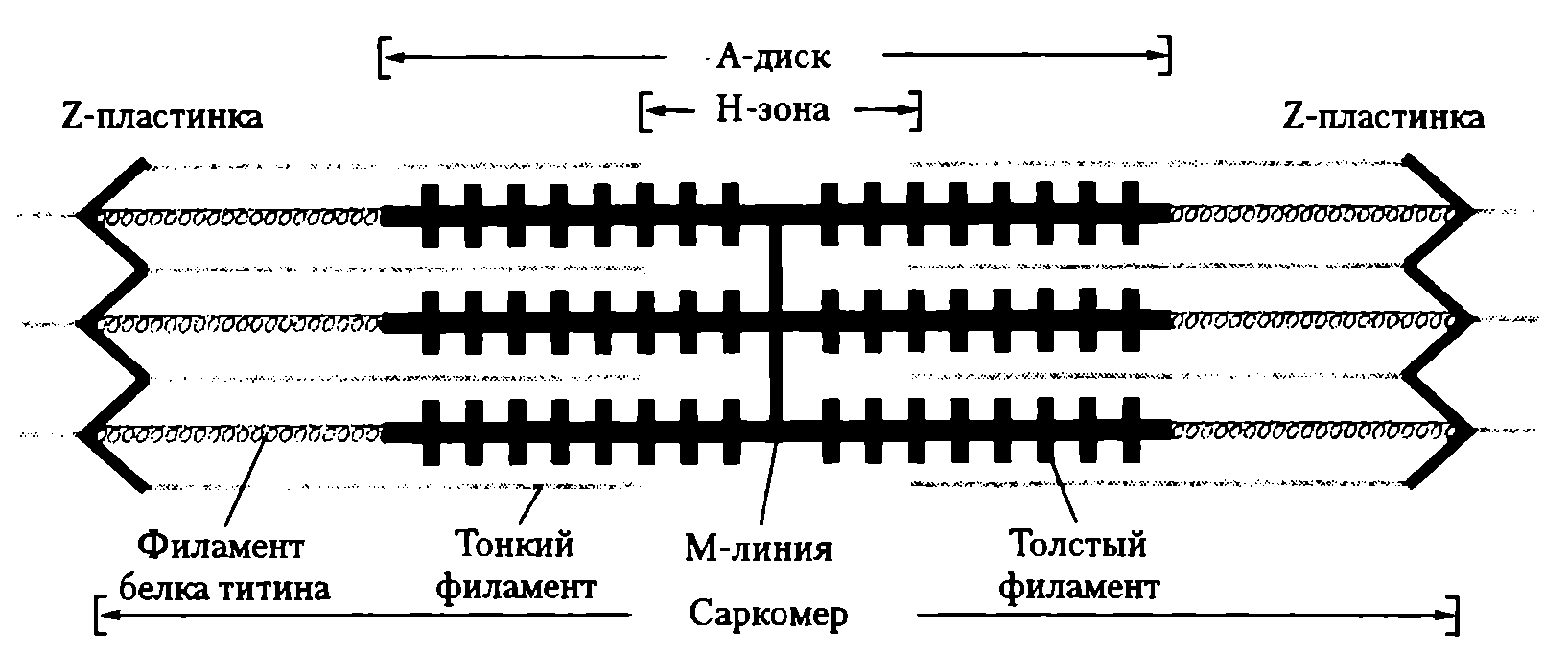
Рис. 22. Саркомер миоцита скелетной мышцы (A. Vander, J. Sherman, D. Luciano, 2004)
Элементы саркoмера (см. рис. 21). Миозиновые протофибриллы образуют наиболее темную часть саркомера — А-диск (анизотропный, он сильно поляризует белый свет). Более светлый участок в центре А-диска называют Н-зоной. Светлый участок саркомера между двумя А-дисками называют 1-диском (изотропный, почти не поляризует свет). Он образован актиновыми протофибриллами, идущими в обе стороны от Z-полосок. Каждый саркомер имеет два набора тонких нитей, прикрепленных к Z-полоскам, и один комплект толстых нитей, сосредоточенных в А-диске. В расслабленной мышце концы толстых и тонких филаментов в разной степени перекрывают друг друга на границе между А- и 1-дисками.
Классификация мышечных волокон:
По структурно-функциональным свойствам и цвету выделяют две основные группы мышечных волокон: быстрые и медленные.
Белые (быстрые) мышечные волокна содержат больше миофибрилл и меньше — митохондрий, миоглобина и жиров, но больше гликогена и гликолитических ферментов; эти волокна называют гликолитическими. Капиллярная сеть, окружающая эти волокна, относительно редкая. Скорость рабочего цикла у данных волокон примерно в 4 раза больше, чем у медленных, что объясняется более высокой АТФазной активностью быстрых волокон, но они обладают малой выносливостью. У белых мышечных волокон число нитей актина и миозина больше, чем у красных, поэтому они толще и сила их сокращения больше, чем у красных волокон.
Красные мышечные волокна содержат много митохондрий, миоглобина, жирных кислот. Эти волокна окружены густой сетью кровеносных капилляров, они имеют меньший диаметр. Митохондрии обеспечивают высокий уровень окислительного фосфорилирования, поэтому данные волокна называют оксидативными. Красные мышечные волокна подразделяются на две подгруппы: быстрые и медленные. Медленные волокна могут выполнять работу в течение относительно продолжительного периода времени; утомление в них развивается медленнее. Они более приспособлены к тоническим сокращениям. Красные быстрые волокна по скорости утомления занимают промежуточное положение между белыми и красными медленными. Скорость их сокращения близка к скорости сокращения белых волокон, что также объясняется высокой АТФазной активностью миозина красных быстрых волокон.
Также имеется незначительное число истинных тонических мышечных волокон; на них локализуется по 7-10 синапсов, принадлежащих, как правило, нескольким мотонейронам, например, в глазодвигательных мышцах, мышцах среднего уха. ПКП этих мышечных волокон не вызывают генерации ПД в них, а непосредственно запускают мышечное сокращение.
Группа мышечных волокон, двигательную (нейромоторную) единицу. В мышцах, совершающих быстрые и точные движения, например в глазодвигательных, нейромоторные единицы состоят из 3—5 мышечных волокон. В мышцах, осуществляющих менее точные движения (например, мышцы туловища и конечностей), двигательные единицы включают сотни и тысячи мышечных волокон. Большая двигательная единица, по сравнению с малой, включает крупный мотонейрон с относительно толстым аксоном, который образует большое число концевых веточек в мышце и, следовательно, иннервирует большое число мышечных волокон. Все мышечные волокна одной двигательной единицы, независимо от их количества, относятся к одному типу. Все скелетные мышцы по своему составу являются смешанными, т.е. образованы красными и белыми мышечными волокнами.
Специфическим свойством всех мышц является сократимость — способность сокращаться, т.е. укорачиваться или развивать напряжение. Реализация этой способности осуществляется с помощью возбуждения и его проведения по мышечному волокну (свойства соответственно возбудимости и проводимости).
Скелетные мышцы не обладают автоматией, управляются организмом произвольно импульсацией из ЦНС, поэтому их называют также произвольными. Гладкие мышцы по собственному желанию не сокращаются, поэтому их называют также непроизвольными, но они обладают автоматией.
Функции скелетной мышцы:
Обеспечение двигательной активности организма — поиск и добывание воды и пищи, ее захват, жевание, глотание, оборонительные реакции, трудовая деятельность — физическая и творческая работа художника, писателя, ученого, композитора в конечном итоге выражается в движении: рисование, письмо, игра на музыкальном инструменте и т.п.
Обеспечение дыхания (движений грудной клетки и диафрагмы).
Коммуникативная функция (устная и письменная речь, мимика и жесты).
Участие в процессах терморегуляции организма с помощью изменения интенсивности сократительного термогенеза.
Теория по нормальной физиологии на тему: Синапс. Физиология мышечных волокон. В данной статье рассматривается механизм синаптической передачи, НМС…
При создании данной страницы использовалась лекция по соответствующей теме, составленная Кафедрой Нормальной физиологии БашГМУ
Синапс — это специфическое место контакта двух возбудимых систем (клеток) для передачи возбуждения.
По способу передачи сигналов:
- механические,
- химические,
- электрические.
По виду медиатора: холинэргические и др.
Нервно-мышечный синапс (НМС) — химический, передача с помощью медиатора ацетилхолина.
Синонимы к слову НМС:
- Нервно-мышечное соединение;
- Моторная концевая пластинка.

Аксоны нервных клеток на своих окончаниях теряют миелиновую оболочку , ветвятся, и концевые веточки аксона утолщаются. Это пресинаптическая терминаль или бляшка или пуговка , которая погружается в углубление на поверхности мышечного волокна.
Покрывающая концевую веточку аксона поверхностная мембрана называется пресинаптической мембраной , т.е. это мембрана, покрывающая поверхность синаптической бляшки (терминали аксона).
Мембрана, покрывающая мышечное волокно в области синапса, называется постсинаптической мембраной , или концевой пластинкой. Она имеет извитую структуру, образуя многочисленные складки, уходящие вглубь мышечного волокна, за счет чего увеличивается площадь контакта.

На постсинаптической мембране находятся белковые структуры — рецепторы , способные связывать медиатор . В одном синапсе количество рецепторов достигает 10-20 млн.
Между пре- и постсинаптическими мембранами находится синаптическая щель , размеры ее в среднем 50 нм, она открывается в межклеточное пространство и заполнена межклеточной жидкостью .
В синаптической щели находится мукополисахаридное плотное вещество в виде полосок, мостиков и содержится фермент ацетилхолинэстераза .
В пресинаптической терминали находится большое количество пузырьков или везикул , заполненных медиатором — химическим веществом посредником, осуществляющим передачу возбуждения.
В нервно-мышечном синапсе медиатор — ацетилхолин (АХ).

АХ синтезируется из холина и уксусной кислоты (ацетил-коэнзима А) с помощью фермента холинэстеразы. Эти вещества перемещаются из тела нейрона по аксону к пресинаптической мембране. Здесь в пузырьках происходит окончательное образование АХ.
3 фракции медиатора:
- Первая фракция — доступная — располагается рядом с пресинаптической мембраной.
- Вторая фракция — депонированная — располагается над первой фракцией.
- Третья фракция — диффузно рассеянная — наиболее удаленная от пресинаптической мембраны.
Механизм синаптической передачи



Ионы Ca вызывают образование специального белкового комплекса , который включает в себя везикулу и структуры, расположенные непосредственно около пресинаптической мембраны.
Они связаны между собой так называемыми белками экзоцитоза.
Часть белков расположена на везикулах (синапсин, синаптотагмин, синаптобревин), а часть — на пресинаптической мембране (синтаксин, синапсо-ассоциированный белок). Данный комплекс получил название секретосома .



Излитию содержимого пузырька в щель способствует белок синаптопорин , формирующий канал, по которому идет выброс медиатора.
Квант медиатора — количество молекул, содержащихся в одной везикуле.
На 1 ПД выбрасывается 100 квантов АХ.





На постсинаптической мембране возникает потенциал концевой пластинки (ПКП). Он является аналогом локального ответа (ЛО).
Потенциал действия на постсинаптической мембране не возникает ! Он формируется на соседней мембране мышечного волокна.
- связывание с рецептором,
- разрушение ферментов (ацетилхолинэстеразой),
- обратное поглощение в пресинаптическую мембрану,
- вымывание из щели и фагоцитоз.
События в синапсе :
- ПД приходит к терминали аксона;
- Он деполяризует пресинаптическую мембрану;
- Ca2+ входит в терминаль, что приводит к выделению АХ;
- В синаптическую щель выделяется медиатор АХ;
- Он диффундирует в щель и связывается с рецепторами постсинаптической мембраны;
- Меняется проницаемость постсинаптической мембраны для ионов Na+;
- Ионы Na+ проникают в постсинаптическую мембрану и уменьшают ее заряд — возникает потенциал концевой пластинки (ПКП) .
На самой постсинаптической мембране ПД возникнуть не может, так как здесь отсутствуют потенциалзависимые каналы, они являются хемозависимыми!
- ПКП суммируются и достигают КУД на соседнем участке мышечного волокна, что приводит к возникновению ПД и его распространению по мышечному волокну (около 5 м/с).
Достигнув пороговой величины, то есть КУД, ПКП возбуждает соседнюю (внесинаптическую) мембрану мышечного волокна за счет местных круговых токов.

Особенности проведения возбуждения в нервно-мышечном синапсе
Одностороннее проведение возбуждения — только в направлении от пресинаптического окончания к постсинаптической мембране.
Суммация возбуждения соседних постсинаптических мембран.
Синаптическая задержка — замедление в проведении импульса от нейрона к мышце составляет 0,5-1 мс. Это время затрачивается на секрецию медиатора, его диффузию к постсинаптической мембране, взаимодействие с рецептором, формирование ПКП, их суммацию.
Низкая лабильность — она составляет 100-150 имп/с для сигнала, что в 5-6 раз ниже лабильности нервного волокна.
Чувствительность к действию лекарственных веществ, ядов, БАВ, выполняющих роль медиатора.
Утомляемость химических синапсов — выражается в ухудшении проводимости вплоть до блокады в синапсе при длительном функционировании синапса. Главная причина утомляемости — исчерпание запасов медиатора в пресинаптическом окончании.
Законы проведения возбуждения по нервам:
- Закон функциональной целостности нерва.
- Закон изолированного проведения возбуждения.
- Закон двустороннего проведения возбуждения.
В зависимости от скорости проведения возбуждения нервные волокна подразделяются на 3 группы: A, B, C. В группе A выделяют 4 подгруппы: альфа, бетта, гамма и сигма.
Физиология мышечных волокон
- скелетная (40-50% массы тела),
- сердечная (менее 1%),
- гладкая (8-9%).
Физиологические свойства скелетных мышц:
- Возбудимость — способность отвечать на действие раздражителя возбуждением.
- Проводимость — способность проводить возбуждение из места его возникновения к другим участкам мышцы.
- Лабильность — способность мышцы сокращаться в соответствии с частотой действия раздражителя (200-300 Гц для скелетной мышцы).
- Сократимость — для мышцы является специфическим свойством — это способность мышцы изменять длину или напряжение в ответ на действие раздражителя.
Физические свойства скелетных мышц:
- Растяжимость — способность мышцы изменять длину под действием растягивающей силы.
- Эластичность — способность мышцы восстанавливать первоначальную длину или форму после прекращения действия растягивающей силы.
- Силы мышц — способность мышцы поднять максимальный груз.
- Способность мышцы совершать работу.
Режимы сокращения:
- Изотонический,
- Изометрический,
- Ауксотонический.
Изотонический режим — сокращение мышцы происходит с изменением ее длины без изменения напряжения (тонуса) (напр.: сокращение мышц языка).
Изометрический режим — длина постоянная, увеличивается степень мышечного напряжения (тонуса) (напр.: при поднятии непосильного груза).
Ауксотонический режим — одновременно изменяется длина и напряжение мышцы (характерен для обычных двигательных актов).
Механизм сокращения поперечно-полосатых мышц
Любая скелетная мышца состоит из мышечных волокон, которые, в свою очередь, состоят из множества тонких нитей — миофибрилл , расположенных продольно. Каждая миофибрилла состоит из протофибрилл — нитей сократительных белков: миозина (миозиновая протофибрилла), актина (актиновая протофибрилла).
Кроме сократительных белков в миофибрилле имеются два регуляторных белка: тропомиозин и тропонин .
Миозиновые волокна соединены в толстый пучок, от которого в торону актиновых нитей отходят поперечные мостики. У каждого мостика выделяют шейку и головку.
Нить актина располагается в виде 2 скрученных ниток бус. На ней имеются актиновые центры.

Тропомиозин в виде спиралей оплетает поверхность актина, закрывая в покое ее центры. Одна молекула тропомиозина контактирует с 7 молекулами актина.
Тропонин образует утолщение на конце каждой нити тропомиозина.
Под влиянием возникшего в мышечном волокне ПД из саркоплазматического ретикулума (СПР — депо Ca2+) высвобождаются ионы Ca. Кальций связывается с тропонином, который смещает тропомиозиновый стержень, что приводит к открытию актиновых центров.

В результате, к актиновым центрам присоединяются головки поперечных миозиновых мостиков.
Процесс расслабления происходит в обратной последовательности с использованием энергии АТФ за счет функционирования кальциевого насоса.
При отсутствии повторного импульса ионы Ca не поступают из СПР. В результате отсутствия Ca-тропонинового комплекса, тропомиозин возвращается на свое прежнее место, блокируя актиновые центры актина. Актиновые протофибриллы легко скользят в обратном направлении благодаря эластичности мышцы, и мышца удлиняется (расслабляется).
Гладкие мышцы
Гладкие мышцы — это мышцы, формирующие слой стенок полых внутренних органов. Они построены из веретенообразных одноядерных мышечных клеток без поперечной исчерченности за счет хаотичного расположения миофибрилл.
Особенности гладких мышц:
- Иннервируются волокнами вегетативной нервной системы (ВНС);
- Обладают низкой возбудимостью:
- Обладают низкой величиной МП (мембранного потенциала) — -50 — -60 мВ из-за более высокой проницаемости для ионов Na+
- ПД (потенциал действия) отличается меньшей амплитудой и большей длительностью. Он формируется в основном за счет ионов Ca2+
- Медленная проводимость:
Клетки в гладких мышцах функционально связаны между собой посредством щелевидных контактов — нексусов, которые имеют низкое электрическое сопротивление. За счет этих контактов ПД распространяется с одного мышечного волокна на другое, охватывая большие мышечные пласты, и в реакцию вовлекается вся мышца.
Гладкие мышцы способны осуществлять относительно медленные ритмические и длительные тонические сокращения.
Медленные ритмические сокращения обеспечивают перемещение содержимого органа из одной области в другую.
Длительные тонические сокращения, особенно сфинктеров полых органов, препятствуют выходу из них содержимого.
Это способность сохранять приданную им при растяжении или деформации форму. Благодаря пластичности гладкая мышца может быть полностью расслаблена как в укороченном, так и в растянутом состоянии.
Особенность гладких мышц, отличающая их от скелетных. Благодаря автоматии гладкие мышцы могут сокращаться в условиях отсутствия иннервации . Важную роль в этом играет растяжение.
Растяжение является адекватным раздражителем для гладкой мускулатуры. Сильное и резкое растяжение гладких мышц вызывает их сокращение.
Сравнительная характеристика скелетных и гладких мышц:
Читайте также: