
Где расположен спиральный ганглий
Гистологический препарат. №19
Орган слуха. Улитка (спиральный орган) Аксиальный срез пирамиды височной кости.
Окраска гематоксилином и эозином. Увеличение малое и большое. Найти:
- костный лабиринт;
- преддверную (вестибулярную) лестницу,
- барабанную (тимпанальную) лестницу;
- перепончатый канал улитки;
- вестибулярную (рейснерову) мембрану;
- спиральную костную пластинку;
- лимб спиральной костной пластинки;
- связку;
- сосудистую полоску;
- базилярную пластинку;
- спиральный ганглий улитки,
- спиральный ганглий улитки;
- спиральный орган и в нем:
- наружные волосковые клетки;
- внутренние волосковые клетки;
- наружные столбовые клетки;
- покровную пластинку;
- внутренние фаланговые опорные клетки;
- наружные фаланговые опорные клетки;
- туннель;
- внутренние столбовые клетки.
Дополнительный материал
Орган слуха и равновесия (преддверно-улитковый орган)
В состав преддверно-улиткового органа входят: наружное, среднее и внутреннее ухо, воспринимающее звуковые, гравитационные, вибрационные стимулы линейных и угловых ускорений. В наружном ухе различают ушную раковину, наружный слуховой проход, барабанную перепонку. Среднее ухо представленно барабанной полостью, слуховыми косточками, слуховой трубой. У млекопитающих и человека рецепторные клетки органа слуха и равновесия располагаются во внутреннем ухе в перепончатом лабиринте, ограниченном костным лабиринтом. При этом волосковые сенсорные эпителиоциты органа слуха находятся в улитковом лабиринте, в спиральном органе улитки, а рецепторы органа равновесия — в вестибулярном лабиринте — в пятнах мешочков и гребешках полукружных каналов.
В процессе эмбриогенеза перепончатый лабиринт внутреннего уха закладывается из парных утолщений эктодермы (слуховые и лабиринтные плакоды). Они погружаются в подлежащую мезенхиму и превращаются в слуховые пузырьки. Дифференцировка слуховых пузырьков приводит к разделению на два зачатка — органа равновесия и органа слуха. Одновременно слуховой пузырек контактирует с эмбриональным слуховым нервным ганглием, который также делится на две части — ганглий преддверия и ганглий улитки.
Спиральный орган. Орган слуха.
Спиральный орган, воспринимающий звуки, состоит из сенсорных и поддерживающих, опорных клеток. Среди обеих групп различают внутренние и наружные клетки. Внутренние сенсорные клетки — эпителиоциты, имеющие кувшинообразную форму с расширенным основанием, лежат в один ряд. Апикальная часть этих клеток покрыта кутикулой с короткими подвижными и уплотненными микроворсинками (волосками) — стереоцилиями. Наружные волосковые клетки имеют цилиндрическую форму с округлым основанием. Они также несут на своей апикальной поверхности кутикулярную пластинку со слуховыми волосками-стереоцилиями. Наружные волосковые клетки лежат в три параллельных ряда, а в верхних завитках улитки их может быть 4 — 5 рядов. Наружные волосковые клетки значительно чувствительнее к звукам большой интенсивности, чем внутренние. Цитоплазма сенсорных клеток богата окислительными ферментами, монофосфоэстеразой, содержит РНК. Наружные сенсорные эпителиоциты содержат много гликогена, в их стереоцилиях обнаружена ацетилхолинэстераза. Своими основаниями волосковые клетки расположены во вдавлениях, образованных телами подлежащих опорных поддерживающих, фаланговых клеток. Если наружные и внутренние волосковые эпителиоциты располагаются на соответствующих опорных, фаланговых клетках, то опорные клетки лежат на базальной мембране. В их цитоплазме имеются тонофибриллы. В спиральном органе на базальной мембране расположена еще одна разновидность опорных клеток — столбовые эпителиоциты внутренние (обращенные от туннеля к лимбу) и наружные обращенные от туннеля к сосудистой полоске), которые, соприкасаясь своими вершинами, образуют внутренней туннель, заполненный эндолимфой. Через туннель проходят безмякотные нервные волокна, идущие от нейронов спирального ганглия к сенсорным клеткам. Терминала дендритов биполярных нейронов спирального ганглия подходят к основаниям сенсорных клеток и образуют синапсы. На базилярной мембране этого органа рядом с наружными фаланговыми располагаются наружные пограничные эпителиоциты, богатые гликогеном. Эти клетки выполняют трофическую функцию. Латеральнее их, также на базальной мем-бране, находятся наружные поддерживающие эпителиоциты — клетки кубической формы, которые постепенно переходят в эпителий, выстилающий сосудистую полоску.
Звуковые воздействия с барабанной перепонки передаются на молоточек, наковальню и стремечко, а затем через овальное окно на перилимфу, базилярную и покровную мембраны. При этом происходит отклонение стереоцилий и возбуждение рецепторных клеток. Ацетилхолин, содержащийся в эндолимфе, взаимодействует с холинрецепторным белком, вмонтирвоанным в мембраны стереоцилий, а также с ацетилхолинэстеразой. Последняя разрушает ацетилхолин. Это приводит к возникновению микрофонного эффекта. Далее афферентные нервные окончания биполярных нейронов спирального ганглия несут импульсы в вышележащие отделы слухового анализатора.
Орган равновесия.
Вестибулярная часть перепончатого лабиринта. В вестибулярной части перепончатого лабиринта расположены рецепторы органа равновесия. Он состоит из двух мешочков — сферического и эллиптического, со-общающихся при помощи узкого канала и связанных с тремя полукружными каналами. В местах соединения каналов с эллиптическим мешочком (маточка) имеются расширения — ампулы. В ампулах и мешочках располагаются чув-ствительные (сенсорные) клетки. В мешочках эти участки называются пятна-ми (макулами), а в ампулах — гребешками (кристами). Пятна мешочков представлены эпителием, расположенным на базальной мембране и состоящим из сенсорных и опорных клеток. Поверхность эпителия покрыта особой студенистой отолитовой мембраной со статокониями. Статоконии или отолиты состоят из кристаллов карбоната кальция.

Волосковые сенсорные клетки по строению подразделяются на два типа. Один из них — грушевидные эпителиоциты с широким основанием, к кото-рым примыкают нервные окончания в виде чаши. Клетки второго типа — столбчатые, имеют призматическую форму, у их основания находятся точечные нервные окончания. На наружной поверхности этих клеток имеется кутикула, от которой отходят 60 — 80 неподвижных волосков-стереоцилий и одна подвижная ресничка-киноцилия, контактирующие с отолитовой мембраной. При смещении киноцилии в сторону стереоцилий клетка возбуждается, а если движение направлено в противоположную сторону, то происходит торможение клетки. Полученные через афферентные синапсы импульсы передаются через вестибулярный нерв в соответствующие части вестибулярного анализатора. Поддерживающие клетки располагаются между сенсорными. Они отличаются овальными темными ядрами, имеют микроворсинки и большое количество ми-тохондрий. Пятна воспринимают гравитацию (сила тяжести), линейные ускорения, а макула сферического мешочка к тому же воспринимает и вибрационные колебания.

Ампулярные гребешки (кристы). Они находятся в ампулярных расширениях полукружных каналов. Ампулярные гребешки состоят из таких же сенсорных и поддерживающих эпителиоцитов, как и пятна. Апикальная часть этих клеток окружена желатинообразным прозрачным куполом, лишенным полости. В этот купол входят волоски сенсорных клеток. Ампулярные гребешки воспринимают угловые ускорения (повороты тела и головы). Нервные волокна, разветвляющиеся вокруг волосковых клеток гребешков ампул и пятен, принадлежат биполярным чувствительным нейронам, находящимся в вестибулярном ганглии. Центральные отростки клеток ганглия направляются в продолговатый мозг и заканчиваются на клетках вестибулярных ядер. От этих клеток импульс поступает далее в вышележащие отделы анализатора.
Методичка МГМСУ в формате PDF — скачать и читать со страницы 25 (Орган слуха и равновесия.)
Методичка МГМСУ. Частная гистология.
Спиральный ганглий располагается в канале Розенталя, который находится в модиолусе улитки. Он содержит тела афферентных нейронов, дендриты, которых возбуждаются под действием нейромедиаторов, секретируемых клетками кортиева органа, и аксоны, которые направляются в улитковые ядра расположенные в стволе головного мозга. Большинство (приблизительно 95%) афферентных волокон являются толстыми, покрыты миелиновой оболочкой и берут начало от нейронов I типа.
Эти волокна иннервируют исключительно внутренние волосковые клетки. Оставшиеся афферентные волокна тонкие, не имеют миелиновой оболочки и исходят из нейронов II типа, эти волокна контактируют с наружными волосковыми клетками. Около десятка нейронов I типа иннервируют каждую внутреннюю волосковую клетку (сходящаяся иннервационная модель), в то время как афферентные нервные волокна II типа разделяются на несколько ветвей и контактируют с несколькими наружными волосковыми клетками (расходящаяся иннервационная модель).
Вся слуховая информация переносится к стволу мозга по афферентной системе. Слуховой и вестибулярный нервы соединяются друг с другом в преддверно-улитковом нерве (VIII черепной нерв).
Эфферентные волокна берут начало в стволе головного мозга от нейронов, расположенных в комплексе верхней оливы и посылают информацию к улитке посредством синаптических соединений с наружными волосковыми клетками, а также с афферентными волокнами внутренних волосковых клеток. Эфферентная система позволяет центральной нервной системе регулировать работу улитки.
Иннервационная структура Кортиева органа отражает функциональное различие двух типов улитковых волосковых клеток.
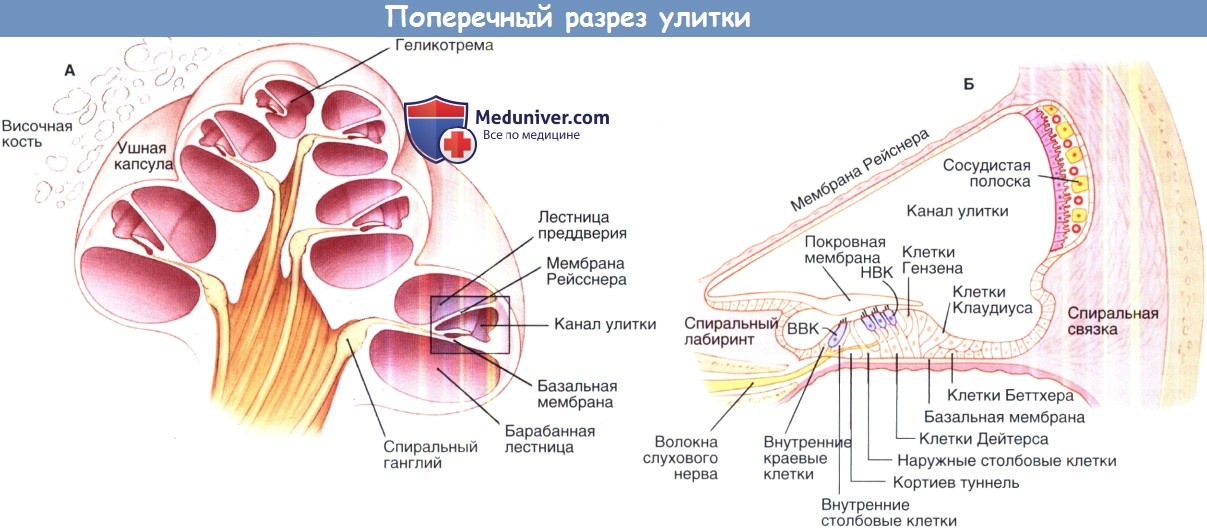
Поперечный разрез улитки. А, схематичное изображение улитки в разрезе.
Выделенная область, показанная на рисунке Б, изображает улитковый проток и окружающие структуры, такие как кортиев орган и сосудистую полоску в разрезе.
ВВК — внутренние волосковые клетки; НВК — наружные волосковые клетки.
а) Нейропроцессинг слуховой информации и синапсы внутренних волосковых клеток. Афферентная нейропередача, осуществляемая внутренними волосковыми клетками, инициируется синапсами ленточного типа количеством от 5 до 30, где поток ионов кальция через потенциал-зависимые кальциевые каналы приводит к четко дозированному высвобождению синаптических пузырьков в пресинаптическую область. В результате экзоцитоз нейротрансмиттеров прямо пропорционален пресинаптическому току кальция, который в свою очередь зависит от изменения потенциала, обусловленного механотрансдукцией.
Информация, кодируемая на афферентных синапсах, поразительно точна, что позволяет получить высокую временную точность, так же как и значительный динамический диапазон с пятикратной амплитудой от 0 до более 100 дБ.
Каждый ленточный синапс состоит из пресинаптического плотного тела (схожего с ленточным синапсом фоторецепторных клеток) в окружении пузырьков с нейротрансмиттерами, плотной полоски пресинаптической мембраны, синаптической щели и постсинаптической области, содержащей АМРА-глютаминэргические рецепторы афферентных нейронов. Считается что глутамат или вещество, близкое по составу, является нейротрансмиттером афферентных связей внутренних волосковых клеток, хотя в этом процессе могут также быть задействованы другие, еще нераспознанные трансмиттеры.
Тонотопическая организация кортиева органа происходит благодаря афферентной системе, где деполяризация внутренних волосковых клеток в определенных участках ведет к возбуждению афферентных нейронов спирального ганглия. Каждое афферентное волокно характеризуется определенной настроечной кривой, описывающей необходимый для получения ответа на заданной частоте уровень звукового давления. Особенность настроечных кривых заключается в отражении частоты наивысшей чувствительности нервного волокна, т.е. частотной характеристики.
Стимулы выше или ниже характерной частоты могут также вызывать ответ, но только при наличии более высокой интенсивности. Форма и пороги чувствительности настроечных кривых афферентных нервных волокон зависят от множества факторов, включая морфологию кортиева органа и активные процессы, связанные с улитковой амплификацией. Активность эфферентной системы играет важную роль в модуляции характеристик афферентных нервов. Потеря улитковой амплификации, в частности, в результате утраты наружных волосковых клеток, ведет к расширению настроечных кривых и увеличению порогов ответа нервного волокна.
Тонотопическая организация улитки является основой частотной кодировки в волокнах слухового нерва (кодирование позиции). Кроме того, выходные характеристики частот кодируются волокнами слухового нерва, это явление известно, как фазовая синхронизация. Волокна слухового нерва посылают сигнал в определенной фазе с частотным стимулом, приводящим к регулярному ответу с характерными интервалами между пиками, равными или кратными длине волны стимула. Фазовая синхронизация возникает только на низких частотах.
Свыше 5 кГц возбуждение, идущее от волокон слухового нерва, возникает с хаотическими интервалами. Тонотопическая организация и фазовая синхронизация крайне важны для выделения сигнала нужной частоты. Выходная частота волокон слухового нерва определяется не только частотными характеристиками, но также и интенсивностью стимулов. По мере увеличения интенсивности стимула увеличивается импульсная частота слухового нерва. Аналогичным образом количество волокон слуховых нервов, активированных на заданных частотах, увеличивается по мере усиления стимула.
Это является результатом вовлечения в процесс большего количества нервных волокон в результате того, что волокна слухового нерва при одинаковых частотных характеристиках имеют разные пороги частотного ответа. Кроме того, при росте интенсивности стимула активируются и другие афферентные нервные волокна с похожими частотными характеристиками. Эти важные физиологические характеристики определяют ряд правил, обязательных при изготовлении и применении имплантатов улитки.
Таким образом, частотно-зависимая стимуляция волосковых клеток ведет к увеличению синаптической передачи в ленточных синапсах внутренних волосковых клеток. Последующее возбуждение постсинаптических токов в волокнах афферентного нерва ведет к временной разрядке потенциала действия в слуховом нерве. В действительности, временное разрешение и динамический диапазон слуховой системы не имеют равных среди сенсорных систем.
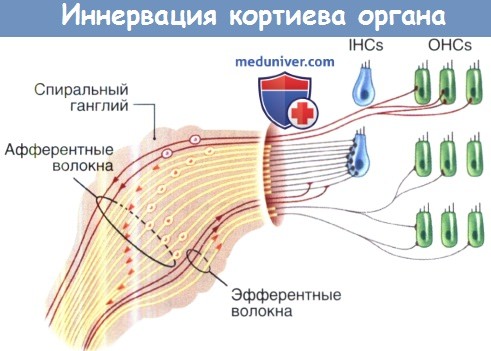
Иннервация кортиева органа.
Схема отображает афферентную и эфферентную иннервацию внутренних и наружных волосковых клеток.
Сверху вниз показаны немиелиновые и миелиновые афферентные волокна II типа,
немиелоновые эфферентные волокна латеральных оливокохлеарных путей (ЛОК) и миелиновые эфферентные волокна медиальных оливокохлеарных путей (МОК).
б) Эфферентная иннервация улитки. В головном мозге четко различают две группы эфферентных нервных волокон. Первые, миелиновые медиальные оливокохлеарные (МОК) эфферентные волокна, берут начало от нейронов, расположенных около верхнего медиального оливного ядра. МОК волокна направляются к контраи ипсилатеральной улитке, где они образуют холинергические синапсы с наружными волосковыми клетками. Вторая группа волокон представлена безмиелиновыми латеральными оливокохлеарными (ЛОК) эфферентными волокнами, которые берут начало от нейронов с маленькими телами, локализующимися около латерального верхнего ядра оливы.
ЛОК волокна направляются преимущественно к ипсилатеральной улитке, где они оканчиваются на дендритах афферентных нейронов I типа у основания внутренних волосковых клеток. Синапсы ЛОК волокон являются сложным химическим комплексом и используют холинэргическую, ГАМК-эргическую и дофаминэргическую передачу, а также различные нейропептиды.
Эффект от стимуляции МОК волокон был изучен подробнее, чем стимуляция ЛОК волокон. В целом стимуляция МОК системы ведет к увеличению порогов чувствительности из-за снижения степени улитковой амплификации наружными волосковыми клетками. Это обусловленная звуком обратная связь, следовательно, чувствительность слухового аппарата снижается в ситуациях, когда нет нужды в энергозатратных механизмах метаболической амплификации.
Функция ЛОК нервных волокон представляется более сложной. Их непосредственный вход в афферентные нейроны позволяет предположить, что они регулируют афферентную активность, тем самым оказывая влияние на динамический диапазон. Исследование различных патологических состояний подтверждает эту точку зрения тем, что снижение специфических нейротрансмиттеров или разрушение тел нейронов в стволе головного мозга ведет либо к усилению, либо к супрессии ответа слухового нерва.
Эффекты обратной связи ЛОК волокон медленны и обычно проявляются в течение минут. Еще одна функция ЛОК системы заключается в обеспечении медленной интеграции и настройки бинауральной функции, необходимой для точного различения локализации звука.
В итоге, активация МОК и ЛОК эфферентной системы, по-видимому, имеет защитный эффект, направленный против акустического повреждения и такая обратная связь может быть важна в шумной окружающей среде.
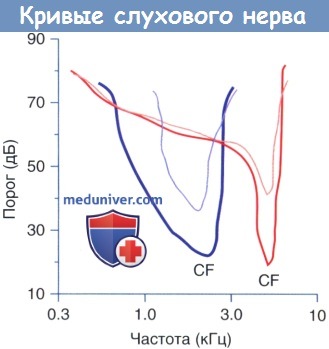
Характерные настроечные кривые слухового нерва (толстая линия).
Настроечная кривая слухового нерва с частотной характеристикой около 2 кГц (синяя) и 5 кГц (красная) (тонкие линии).
Значительное снижение порогов слуховой чувствительности в ответ на стимуляцию МОК системы.
Изменения специфической формы настроечных кривых зависят от частотной характеристики (CF) отдельно взятого волокна.
- Вернуться в оглавление раздела "отоларингология"
ГАНГЛИИ (ganglia нервные узлы)— скопления нервных клеток, окруженные соединительной тканью и клетками глии, расположенные по ходу периферических нервов.
Различают Г. вегетативной и соматической нервной системы. Г. вегетативной нервной системы делятся на симпатические и парасимпатические и содержат тела постганглионарных нейронов. Г. соматической нервной системы представлены спинномозговыми узлами и Г. чувствительных и смешаных черепных нервов, содержащих тела чувствительных нейронов и дающих начало чувствительным порциям спинномозговых и черепных нервов.
Содержание
- 1 Эмбриология
- 2 Анатомия
- 3 Гистология
- 4 Физиология
- 5 Патология
- 6 Краткая анатомическая характеристика нервных ганглиев (узлов)
Эмбриология
Зачатком спинномозговых и вегетативных узлов является ганглиозная пластинка. Она образуется у эмбриона в тех отделах нервной трубки, которые граничат с эктодермой. У зародыша человека на 14—16-й день развития ганглиозная пластинка располагается по дорсальной поверхности замкнувшейся нервной трубки. Затем она расщепляется по всей длине, обе ее половины перемещаются вентрально и в виде нервных валиков лежат между нервной трубкой и поверхностной эктодермой. В дальнейшем соответственно сегментам дорсальной стороны зародыша в нервных валиках возникают очаги пролиферации клеточных элементов; эти участки утолщаются, обособляются и превращаются в спинномозговые узлы. Из ганглиозной пластинки развиваются также чувствительные ганглии У, VII—X пар черепных нервов, аналогичные спинномозговым ганглиям. Зародышевые нервные клетки, нейробласты, формирующие спинномозговые ганглии, представляют собой биполярные клетки, т. е. имеют два отростка, отходящих от противоположных полюсов клетки. Биполярная форма чувствительных нейронов у взрослых млекопитающих и человека сохраняется только в чувствительных клетках преддверно-улит кового нерва, преддверного и спирального ганглиев. В остальных, как спинномозговых, так и черепных чувствительных узлах, отростки биполярных нервных клеток в процессе их роста и развития сближаются и сливаются в большинстве случаев в один общий отросток (processus communis). По этому признаку чувствительные нейроциты (нейроны) называют псевдоуниполярными (neurocytus pseudounipolaris), реже протонейронами, подчеркивая древность их происхождения. Спинномозговые узлы и узлы в. н. с. отличаются характером развития и строения нейронов. Развитие и морфология вегетативных ганглиев — см. Вегетативная нервная система.
Анатомия
Основные сведения об анатомии Г. приведены в таблице.
Гистология


Возраст и перенесенные заболевания сказываются на структуре спинномозговых ганглиев — в них появляется значительно большее, чем у здоровых, количество различных атипичных нейронов, в особенности с дополнительными отростками, снабженными колбовидными утолщениями, как, напр., при ревматическом пороке сердца (рис. 2), стенокардии и др. Клинические наблюдения, а также экспериментальные исследования на животных показали, что чувствительные нейроны спинномозговых узлов реагируют значительно быстрее интенсивным ростом дополнительных отростков на различные эндогенные и экзогенные вредности, нежели моторные соматические или вегетативные нейроны. Эта способность чувствительных нейронов иногда выражена значительно. В случаях хрон, раздражений вновь образованные отростки могут обвиваться (в виде намотки) вокруг тела собственного или соседнего нейрона, напоминая кокон. Чувствительные нейроны спинномозговых узлов, как и другие типы нервных клеток, имеют ядро, различные органеллы и включения в цитоплазме (см. Нервная клетка). Т. о., отличительным свойством чувствительных нейронов спинномозговых и узлов черепных нервов является их яркая морфол, реактивность, выражающаяся в изменчивости их структурных компонентов. Это обеспечено высоким уровнем синтеза белков и различных активных веществ и свидетельствует об их функциональной подвижности.
Физиология
У беспозвоночных Г. играют ту же роль, что и ц. н. с. у позвоночных, являясь высшими центрами координации соматических и вегетативных функций. В эволюционном ряду от червей до головоногих моллюсков и членистоногих Г., перерабатывающие всю информацию о состоянии окружающей и внутренней среды, достигают высокой степени организации. Это обстоятельство, а также простота анатомической препаровки, относительно большие размеры тел нервных клеток, возможность введения в сому нейронов под прямым визуальным контролем одновременно нескольких микроэлектродов сделали Г. беспозвоночных общераспространенным объектом нейрофизиол, экспериментов. На нейронах круглых червей, октапод, декапод, брюхоногих и головоногих моллюсков методами электрофореза, прямого измерения активности ионов и фиксации напряжения проводят исследования механизмов генерации потенциалов и процесса синаптической передачи возбуждения и торможения, зачастую невыполнимые на большинстве нейронов млекопитающих. Несмотря на эволюционные различия, основные электрофизиол, константы и нейрофизиол, механизмы работы нейронов во многом одинаковы у беспозвоночных и высших позвоночных животных. Поэтому исследования Г., беспозвоночных имеют общефизиол. значение.
У позвоночных соматосенсорные черепные и спинномозговые Г. функционально однотипны. Они содержат тела и проксимальные части отростков афферентных нейронов, передающих импульсацию от периферических рецепторов в ц. н. с. В сомато-сенсорных Г. нет синаптических переключений, эфферентных нейронов и волокон. Так, нейроны спинномозговых Г. у жабы характеризуются следующими основными электрофизиол, параметрами: специфическим сопротивлением — 2,25 кОм/см 2 для деполяризующего и 4,03 кОм/см 2 для гиперполяризующего тока и удельной емкостью 1,07 мкФ/см 2 . Полное входное сопротивление нейронов сомато-сенсорных Г. существенно ниже, чем соответствующий параметр аксонов, поэтому при высокочастотной афферентной импульсации (до 100 импульсов в 1 сек.) проведение возбуждения может блокироваться на уровне тела клетки. В этом случае потенциалы действия, хотя и не регистрируются от тела клетки, продолжают проводиться с периферического нерва в задний корешок и сохраняются даже после экстирпации тел нервных клеток при условии интактности Т-образных ветвлений аксонов. Следовательно, возбуждение сомы нейронов сомато-сенсорных Г. для передачи импульсов от периферических рецепторов к спинному мозгу не обязательно. Эта особенность впервые появляется в эволюционном ряду у бесхвостых амфибий.
Вегетативные Г. позвоночных в функциональном плане принято разделять на симпатические и парасимпатические. Во всех вегетативных Г. происходит синаптическое переключение с преганглионарных волокон на постганглионарные нейроны. В подавляющем большинстве случаев синаптическая передача осуществляется хим. путем с помощью ацетилхолина (см. Медиаторы). В парасимпатическом цилиарном Г. птиц обнаружена электрическая передача импульсов при помощи так наз. потенциалов соединения, или потенциалов связи. Электрическая передача возбуждения через один и тот же синапс возможна в двух направлениях; в процессе онтогенеза она формируется позже химической. Функциональное значение электрической передачи пока не ясно. В симпатических Г. амфибий выявлено небольшое количество синапсов с хим. передачей нехолинергической природы. В ответ на сильное одиночное раздражение преганглионарных волокон симпатического Г. в постганглионарном нерве прежде всего возникает ранняя отрицательная волна (O-волна), обусловленная возбуждающими постсинаптическими потенциалами (ВПСП) при активации н-холинорецепторов постганглионарных нейронов. Тормозной постсинаптический потенциал (ТПСП), возникающий в постганглионарных нейронах под действием катехоламинов, выделяемых хромаффинными клетками в ответ на активацию их м-холинорецепторов, формирует следующую за 0-волной положительную волну (П-волна). Поздняя отрицательная волна (ПО-волна) отражает ВПСП постганглионарных нейронов при активации их м-холинорецепторов. Завершает процесс длительная поздняя отрицательная волна (ДПО-волна), возникающая вследствие суммации ВПСП нехолинергической природы в постганглионарных нейронах. В нормальных условиях на высоте О-волны при достижении ВПСП величины 8—25 мв возникает распространяющийся потенциал возбуждения с амплитудой 55—96 мв, длительностью 1,5—3,0 мсек, сопровождаемый волной следовой гиперполяризации. Последняя существенно маскирует волны П и ПО. На высоте следовой гиперполяризации возбудимость снижается (период рефрактерности), поэтому обычно частота разрядов постганглионарных нейронов не превышает 20—30 импульсов в 1 сек. По основным электрофизиол. характеристикам нейроны вегетативных Г. идентичны большинству нейронов ц. н. с. Нейрофизиол. особенностью нейронов вегетативных Г. является отсутствие истинной спонтанной активности при деафферентации. Среди пре- и постганглионарных нейронов преобладают нейроны групп В и С по классификации Гассера — Эрлангера, основанной на электрофизиол, характеристиках нервных волокон (см.). Преганглионарные волокна обширно ветвятся, поэтому раздражение одной преганглионарной ветви приводит к возникновению ВПСП во многих нейронах нескольких Г. (феномен мультипликации). В свою очередь на каждом постганглионарном нейроне оканчиваются терминали многих преганглионарных нейронов, отличающихся по порогу раздражения и скорости проведения (феномен конвергенции). Условно мерой конвергенции можно считать отношение количества постганглионарных нейронов к количеству преганглионарных нервных волокон. Во всех вегетативных Г. оно больше единицы (за исключением цилиарного ганглия птиц). В эволюционном ряду это отношение возрастает, достигая в симпатических Г. человека величины 100:1. Мультипликация и конвергенция, обеспечивающие пространственную суммации) нервных импульсов, в сочетании с временной суммацией являются основой интегрирующей функции Г. при обработке центробежной и периферической импульсации. Через все вегетативные Г. проходят афферентные пути, тела нейронов которых лежат в спинномозговых Г. Для нижнего брыжеечного Г., чревного сплетения и некоторых интрамуральных парасимпатических Г. доказано существование истинных периферических рефлексов. Афферентные волокна, проводящие возбуждение с малой скоростью (ок. 0,3 м/сек), входят в Г. в составе постганглионарных нервов и оканчиваются на постганглионарных нейронах. В вегетативных Г. обнаружены окончания афферентных волокон. Последние информируют ц. н. с. о происходящих в Г. функционально-хим. изменениях.
Патология
В клин, практике наиболее часто встречается ганглионит (см.), называемый также симпато-ганглионитом, — заболевание, связанное с поражением ганглиев симпатического ствола. Поражение нескольких узлов определяют как полиганглионит, или трунцит (см.).
Спинальные ганглии нередко вовлекаются в патологический процесс при радикулитах (см.).
2. В этих органах рецепцию раздражений осуществляют не нейросенсорные, а
- специализированные производные эпителия (п. 16.1.2).
2. Но, как и в первично чувствующих органах, соответствующее раздражение вызывает
17.1. Орган слуха и равновесия
17.1.1. Общие сведения
17.1.1.1. Составные части
В составе органа - 3 части: наружное, среднее и внутреннее ухо.
I. Наружное ухо
2. а) Ушная раковина и первая треть слу х ового прохода имеют хрящевую основу;
II. Среднее ухо
III,а. Внутреннее ухо: костный лабиринт
2. Средняя часть - преддверие (vestibulum) (8) - овальная полость, сообщающаяся с соседними отделами лабиринта .
III,б. Внутреннее ухо: перепончатый лабиринт
2. а) Во внутреннем ухе имеется жидкость:
3. а) Итак, в костной улитке находится перепончатая улитка (15) , занимающая примерно треть сечения спирального костного канала.
б ) В костном преддверии перепончатый лабиринт образует два мешочка -
17.1.1.2. Распределение рецепторных функций между частями лабиринта
б) Причём, в каждой части лабиринта (улитке, мешочках преддвери я , полукружных каналах) рецепторные образования имеют строго определённую функцию .
II. Четыре рецепторных образования
в) При этом колебания перилимфы улитки
В отсутствие круглого окна стремечко, из-за несжимаемости жидкости, не могло бы приводить перилимфу в движение.
г) Одновременно колебания перилимфы (и эндолимфы) улитки вызывают
б) При этом н ад чувствительными клетками пятна имеется студенистая мембрана ,
которая, в зависимости от гравитационного притяжения,
б) П ри вращении головы в определённой плоскости
III. Резюме
Вышеизложенное распределение функций можно суммировать в таблице. –
| Спиральный (кортиев) орган (в перепончатой улитке) | Восприятие звука |
| Пятно эллиптического мешочка | Восприятие гравитации |
| Пятно сферического мешочка | Восприятие гравитации и вибрации |
| Ампулы полукружных каналов | Восприятие угловых ускорений при вращении головы и тела |
17.1.1.3. Развитие органа слуха и равновесия
2. При их впячивании последовательно появляются слуховые ямки и слуховые пузырьки , выстланные
(Следовательно, все эпителиоциты рецепторных образований органа слуха и равновесия развиваются из данного типа эмбрионального эпителия ).
3. Затем слуховые пузырьки разделяются перетяжкой на два отдела -
4 . а) Одновременно у станавливается связь со слуховыми нервными ганглиями,
которые делятся на две части - преддверную и улитковую .
Теперь обратимся к гистологическому строению органа слуха и равновесия.
17.1.2. Наружное и среднее ухо
1. В основе ушной раковины (1) -
Его строение и препарат см. в п. 10.2.5.
1. В коже наружного слухового прохода (2) - не только сальные, но и церуминозные железы, которые выделяют ушную серу .
1. Барабанная перепонка (3) покрыта:
В ней преобладают коллагеновые волокна, но имеются и эластические.
Барабанная полость (4) , к ак и внутренняя поверхность барабанной перепонки, выстлана
1. Стенка слуховой трубы (6) выстлана слизистой оболочкой.
Последняя похожа на таковую в дыхательных путях:
2. Под слизистой оболочкой -
17.1.3. Улитковый отдел лабиринта
17.1.3.1. Внутреннее строение костной улитки
I. Срединная часть костной улитки
2. От стержня отходит костная пластинка - спиральный гребешок (2) . Последний

б) Край лимба делится на две части -
II. Подразделение канала костной улитки на 3 части
2. В итоге, канал костной улитки на всём своём протяжении подразделяется на 3 части:
б) Обе лестницы содержат перилимфу, а перепончатая улитка - эндолимфу.
3. а) При этом вестибулярная лестница

б) Барабанная же лестница заканчивается у круглого окна преддверия.
4. Поэтому колебания перилимфы проходят по следующей цепочке:
17.1.3.2. Стенки перепончатой улитки
а) П ерепончат ый лабиринт улитки имеет на разрезе почти треугольную форму .
б) Охарактеризуем три его стороны.
I. Верхнемедиальная стенка
б) Она п рикрепляется к лимбу в области его верхней (вестибулярной) губы (11).
2. Вестибулярная мембрана включает три слоя:

II. Наружная стенка
2. Она имеет два слоя :
3. а) Сосудистая полоска представляет собой

в) Функция сосудистой полоски - трофическая и секреторная ( продукция эндолимфы).
4. Спиральная связка
III. Нижняя стенка
б) Она натянута между
б) А. Основу пластинки составляют коллагеновые волокна, расположенные в матриксе.
Б. Возможно, волокна играют роль струн , которые откликаются на колебания определённой частоты.
3. Со стороны эндолимфы на базилярной пластинке располагается спиральный, или кортиев, орган (9), который
IV. Внутренний угол канала перепончатой улитки
2. В перепончатом канале улитки лимб образует внутренний угол - между
17.1.3.3. Спиральный (кортиев) орган
Наконец, обратимся к ключевому элементу органа слуха - кортиеву органу.
I. Клеточный состав
в (II) (Среднее увеличение)


II. Поддерживающие клетки
б) На каждой такой клетке, как на ложе, располага е тся сенсорн ая клетк а .
III. Сенсорные волосковые клетки
Сенсорные клетки находятся на фаланговых клетках (1); поэтому, в соответствии с числом последних,
2. Апикальная поверхность
На апикальной поверхности сенсорных клеток присутствуют два образования:
а) кутикула (4) - плёнка гликопротеидной природы,
б) особые микроворсинки - стереоцилии (5), которые
17.1.3.4. Восприятие звуковых раздражений
в) Видимо, эфферентные волокна оказывают
г) Медиаторами в образумых ими синапсах являются:
17.1.3.5. Спиральный ганглий

17.1.4. Вестибулярный отдел лабиринта
17.1.4.1. Эллиптический и сферический мешочки преддверия
I. Рецепторные пятна
2. Строение пятен того и другого мешочка практически одинаково. –
В эпителии пятна - три элемента:
II. Сенсорные клетки рецепторных пятен
1. Сенсорные клетки
2. Контакт с нервными окончаниями
а) Грушевидные клетки:
(хотя собственно синаптические контакты образуются только в некоторых участках этой чаши).
б) Цилиндрические клетки:
а) В апикальных отделах тех и других клеток име ю тся
III. Восприятие гравитационных воздействий
1. Цепочка событий такова:
2. В результате, мозг
17.1.4.2. Рецепторные участки полукружных каналов

б) (Среднее увеличение)

в) (Большое увеличение)

б) При вращении тела
17.2. Орган вкуса
17.2.1. Вкусовые почки
а) (Малое увеличение)

б) (Среднее увеличение)

2. а) В частности, на снимк ах - листовидные сосочки (1), покрытые многослойным плоским неороговевающим эпителием.
б) Между сосочками - узкие просветы (2).
в) С этими просветами контактируют вкусовые почки (3) округлой или овальной формы.
3. В эмбриогенезе эти почки развиваются
17.2.2. Клеточный состав
17.2.2.1. Три вида клеток вкусовой почки
( Большое увеличение) 
17.2.2.2. Характеристика клеток
I. Сенсорные, или "светлые", клетки
б) При этом, как правило, во вкусовой почке представлены рецепторы лишь какого-то одного вкуса.
б) Возбуждение вкусовых рецепторов на микроворсинках
II. Прочие клетки вкусовой почки
Б. Благодаря им, происходит постоянное обновление сенсорных и поддерживающих клеток вкусовых почек
Читайте также: