
Химический синапс в цнс
Физиология синаптической передачи
Классификация синапсов
Синапс – это морфофункциональное образование ЦНС, которое обеспечивает передачу сигнала с нейрона на другой нейрон или с нейрона на эффекторную клетку (мышечное волокно, секреторную клетку). Все синапсы ЦНС можно классифицировать следующим образом.
1. По локализации: центральные (головной и спинной мозг) и периферические (нервно-мышечный, нейросекреторный синапс вегетативной нервной системы). Центральные синапсы можно в свою очередь разделить на аксо-аксональные, аксо-дендритические (дендритные), аксо-соматические, дендро-дендритические, дендро-соматические и т.п. Согласно Г. Шенсрду, различают реципрокные синапсы, последовательные синапсы и синаптические гломерулы (различным способом соединенные через синапсы клетки).
2. По развитию в онтогенезе: стабильные (например, синапсы дуг безусловного рефлекса) и динамичные, появляющиеся в процессе индивидуального развития.
3. По конечному эффекту: тормозные и возбуждающие.
4. По механизму передачи сигнала: электрические, химические, смешанные.
5. Химические синапсы можно классифицировать:
а) по форме контакта – терминальные (колбообразное соединение) и преходящие (варикозное расширение аксона);
б) по природе медиатора – холинергические (медиатор – ацетилхолин, АХ), адренергическис (медиатор – норадреналин, НА), дофаминергические (дофамин), ГАМК-ергические (медиатор – гаммааминомасляная кислота), глицинергические, глутаматергические, аспартатсргические, пептидергические (медиатор – пептиды, например, вещество Р), пуринергические (медиатор – АТФ).
Электрические синапсы. В настоящее время признают, что в ЦНС имеются электрические синапсы. С точки зрения морфологии электрический синапс представляет собой щелевидное образование (размеры щели до 2 нм) с ионными мостиками-каналами между двумя контактирующими клетками. Петли тока, в частности при наличии потенциала действия (ПД), почти беспрепятственно перескакивают через такой щелевидный контакт и возбуждают, т.е. индуцируют генерацию ПД второй клетки. В целом, такие синапсы (они называются эфапсами) обеспечивают очень быструю передачу возбуждения. Но в то же время с помощью этих синапсов нельзя обеспечить одностороннее проведение, т. к. большая часть таких синапсов обладает двусторонней проводимостью. Кроме того, с их помощью нельзя заставить эффекторную клетку (клетку, которая управляется через данный синапс) тормозить свою активность. Аналогом электрического синапса в гладких мышцах и в сердечной мышце являются щелевые контакты типа нексуса.
Химические синапсы. По строению химические синапсы представляют собой окончания аксона (терминальные синапсы) или его варикозную часть (проходящие синапсы), которая заполнена химическим веществом – медиатором. В синапсе различают пресинаптический элемент, который ограничен пресинаптической мембраной, постсинаптический элемент, который ограничен постсипаптической мембраной, а также внесинаптическую область и синаптическую щель, величина которой составляет в среднем 50 нм. В литературе существует большое разнообразие в названиях синапсов. Например, синаптическая бляшка – это синапс между нейронами, концевая пластинка – это постсинаптическая мембрана мионеврального синапса, моторная бляшка – это пресинаптичсское окончание аксона на мышечном волокне.
Общие принципы работы химического синапса
Любой химический синапс, независимо от природы медиатора и хеморецептора, активируется под влиянием потенциала действия, прибегающего к пресинапсу от тела нейрона. В результате – происходит деполяризация пресинаптической мембраны, что повышает проницаемость кальциевых каналов пресинаптической мембраны и приводит к увеличению входа в пресинапс ионов кальция. В ответ на это происходит высвобождение квантов (выход из пресинапса) – 100–200 порций (квантов) медиатора. Выйдя в синаптическую щель, медиатор взаимодействует со специфическим рецептором постсинаптической мембраны, что вызывает изменение ионной проницаемости. В синапсах, в которых осуществляется возбуждение постсинантической структуры, обычно происходит повышение проницаемости для ионов натрия, что вызывает деполяризацию постсинаптической мембраны. Эта деполяризация получила определенное название: возбуждающий постсинаптический потенциал (ВПСП). Если его величина достаточно велика и достигает критического уровня деполяризации, то генерируется ПД. В тормозных синапсах в результате взаимодействия медиатора с рецепторами, наоборот, происходит гиперполяризация (за счет, например, увеличения проницаемости для ионов калия и хлора). Это называется тормозным постсинаптическим потенциалом (ТПСГ1). В гиперполяризоваином состоянии клетка снижает свою возбудимость и благодаря этому прекращает отвечать на внешние раздражители или (если она обладала свойством автоматии) уменьшает спонтанную активность.
После каждого цикла проведения импульса медиатор разрушается, например, ацетилхолин разрушается ацстилхолинэстеразой, норадреналин разрушается моноаминоксидазой (МАО) или катсхол-0-метилтрансферазой (КОМТ), либо идет обратный захват в преси-наптическую структуру. В одних случаях захватывается неразрушенный медиатор (например, норадреналип), в других – его осколки (например, холин из ацетилхолина).
Синтез медиатора совершается в пресинаптическом элементе, куда попадают исходные продукты для синтеза и ферменты, необходимые в синтезе. Ферменты образуются в соме нейрона и по аксону, примерно со скоростью 6 мм/сутки, попадают в пресинапс, где используются в процессе синтеза медиатора. Угнетение активности этих ферментов фармакологическим путем может привести к истощению запасов медиатора в синапсе и, следовательно, к снижению его функциональной способности.
Свойства химических синапсов
1. Односторонняя проводимость – одно из важнейших свойств химического синапса. Асимметрия – морфологическая и функциональная – является предпосылкой для существования односторонней проводимости.
2. Наличие синаптической задержки: для того, чтобы в ответ на генерацию ПД в области пресинапса выделился медиатор и произошло изменение постсинаптического потенциала (ВИСИ или ТПСП), требуется определенное время (синаптическая задержка). В среднем оно равно 0,2–0,5 мс.
3. Благодаря синаптическому процессу нервная клетка, управляющая данным постсинаптичсским элементом (эффектором), может оказывать возбуждающее воздействие или, наоборот, тормозное (это определяется конкретным синапсом).
4. В синапсах существует явление отрицательной обратной связи – антидромный эффект. Речь идет о том, что выделяемый в синаптическую щель медиатор может регулировать выделение следующей порции медиатора из этого же пресинаптического элемента путем воздействия на специфические рецепторы пресинаптичсской мембраны.
В мозге имеется ряд медиаторов, вызывающих возбуждение нейрона: норадреналин (его продуцируют адренергические нейроны), дофамин (дофаминергические нейроны), серото-нин, пептиды (пептидергические), глутаминовая кислота, аспарагиновая кислота и т.д. Во всех этих случаях выделяющийся медиатор взаимодействует со специфическим рецептором, в результате чего меняется проницаемость для ионов натрия, калия или хлора, и в итоге развивается деполяризация (ВПСП). Если она достигает критического уровня деполяризации, то возникает ПД (возбуждение нейрона).
Тормозные синапсы образованы специальными тормозными нейронами (точнее, их аксонами). Медиатором могут быть глицин, гамма-аминомасляная кислота (ГАМК) и ряд других веществ. Обычно глицин вырабатывается в синапсах, с помощью которых осуществляется постсинаптическое торможение. При взаимодействии глицина как медиатора с глициновыми рецепторами нейрона возникает гиперполяризация нейрона (ТПСП) и, как следствие, – снижение возбудимости нейрона вплоть до полной его рефрактсрности. В результате этого возбуждающие воздействия, оказываемые через другие аксоны, становятся малоэффективными или неэффективными. Нейрон выключается из работы полностью.
Химические синапсы — это преобладающий тип синапсов в мозге млекопитающих. В таких синапсах взаимодействие между нейронами осуществляется с помощью медиатора (нейротрансмиттера) — вещества, выделяющегося из пресинаптического окончания и действующего на постсинаптическую структуру.
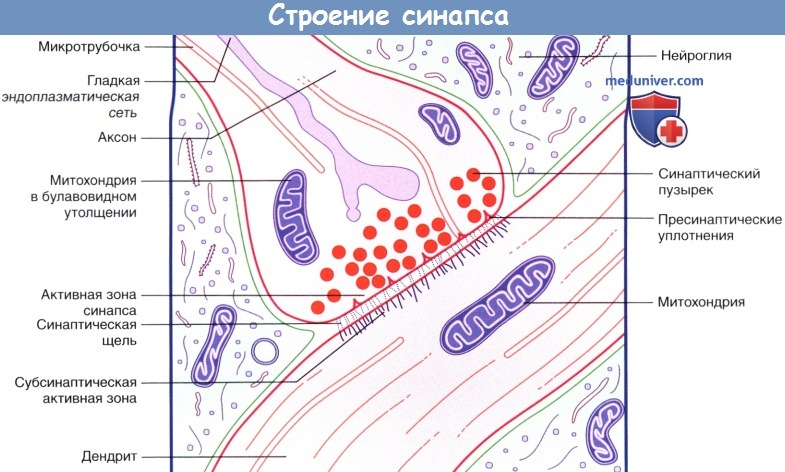
Химические синапсы — это самый сложный вид соединений в ЦНС (рис. 3.1). Морфологически он отличается от других форм соединений наличием хорошо выраженной синаптической щели, при этом виде контакта мембраны строго ориентированы или поляризованы в направлении от нейрона к нейрону.
Химический синапс состоит из двух частей: пресинаптической, образованной булавовидным расширением окончанием аксона передающей клетки, и постсинаптической, представленной контактирующим участком плазматической мембраны воспринимающей клетки. Между обеими частями имеется синаптическая щель — промежуток шириной 10-50 нм между постсинаптической и пресинап- тической мембранами, края которой укреплены межклеточными контактами. В синаптическом расширении имеются мелкие везикулы, так называемые пресинаптические или синаптические пузырьки, содержащие медиатор (вещество-посредник в передаче возбуждения) либо фермент, разрушающий этот медиатор. На постсинаптической, а часто и на пресинаптической мембранах присутствуют рецепторы к тому или иному медиатору.

Рис. 3.1. Химический синапс
Пузырьки (везикулы) размещаются напротив пресинаптической мембраны, что обусловлено их функциональным назначением для высвобождения медиатора в синаптическую щель. Также около пресинаптического пузырька находится большое количество митохондрий (производящих АТФ) и упорядоченные структуры протеиновых волокон. Везикулы имеют различные размеры (от 20 до 150 и более нм) и заполнены химическими веществами, способствующими передаче активности с одной клетки на другую. Одна аксонная терминаль нейрона может содержать несколько типов везикул.
Как правило, из всех окончаний одного нейрона выделяется один и тот же медиатор (правило Дейла). Этот медиатор может подействовать на разные клетки по-разному, в зависимости от их функционального состояния, химизма или степени поляризованности их мембраны. Однако, подчиняясь правилу Дейла, эта пресинаптическая клетка из всех своих аксонных окончаний всегда будет выделять одно и то же химическое вещество. Пузырьки группируются возле уплотненных частей мембраны.
Нервный импульс (возбуждение) с огромной скоростью продвигается по волокну и подходит к синапсу. Этот потенциал действия вызывает деполяризацию мембраны синапса, однако это не приводит к генерации нового возбуждения (потенциала действия), а вызывает открывание специальных ионных каналов. Эти каналы пропускают ионы кальция внутрь синапса. Специальная железа внутренней секреции — паращитовидная (она находится поверх щитовидной) — регулирует содержание кальция в организме. Многие заболевания связаны с нарушением обмена кальция в организме. Например, его недостаток приводит к рахиту у маленьких детей.
Попадая в цитоплазму синаптического окончания, кальций входит в связь с белками, образующими оболочку пузырьков, в которых хранится медиатор. Мембраны синаптических пузырьков сокращаются, выталкивая содержимое в синаптическую щель. Возбуждение (электрический потенциал действия) нейрона в синапсе превращается из электрического импульса в импульс химический. Другими словами, каждое возбуждение нейрона сопровождается выбросом в окончании его аксона порции биологически активного вещества — медиатора. Далее молекулы медиатора связываются с рецепторами (белковыми молекулами), которые находятся на постсинаптической мембране.
Т. е. через мембрану протекает ионный ток, который вызывает изменение потенциала на мембране. Этот потенциал получил название возбуждающего постсинаптического потенциала (рис. 3.2).

Рис. 3.2. Возбуждающий постсинаптический потенциал

Рис. 3.3. Соотношение величин потенциала действия (ПД), возбуждающего постсинаптического потенциала (ВПСП) и тормозного постсинаптического потенциала (ТПСП)
ВПСП — это основной синаптический процесс, обеспечивающий передачу возбуждающих влияний одной клетки на другую. ВПСП отличается от распространяющегося импульса отсутствием рефрактерности, значительной длительностью, способностью суммироваться с другими аналогичными синаптическими процессами, отсутствием способности к активному распространению (рис. 3.3).
Амплитуда потенциала определяется количеством молекул медиатора, связанного рецепторами. Благодаря этой зависимости амплитуда потенциала на мембране нейрона развивается пропорционально количеству открытых каналов.
Область контакта между двумя нейронами называют синапсом.
Внутреннее строение аксодендритического синапса.
а) Электрические синапсы. Электрические синапсы в нервной системе млекопитающих встречаются редко. Они образованы щелевидными контактами (нексусами) между дендритами или сомами соприкасающихся нейронов, которые соединяются с помощью цитоплазматических каналов диаметром 1,5 нм. Процесс передачи сигнала происходит без синаптической задержки и без участия медиаторов.
Посредством электрических синапсов возможно распространение электротонических потенциалов от одного нейрона к другому. Вследствие тесного синаптического контакта модуляция проведения сигнала невозможна. Задача этих синапсов — осуществление одновременного возбуждения нейронов, выполняющих одинаковую функцию. Примером служат нейроны дыхательного центра продолговатого мозга, которые во время вдоха синхронно генерируют импульсы. Кроме того, примером могут служить нейронные цепи, управляющие саккадами, при которых точка фиксации взора перемещается от одного объекта внимания к другому.
б) Химические синапсы. Большинство синапсов нервной системы — химические. Функционирование таких синапсов зависит от высвобождения медиаторов. Классический химический синапс представлен пресинаптической мембраной, синаптической щелью и постсинаптической мембраной. Пресинаптическая мембрана — часть булавовидного расширения нервного окончания клетки, передающей сигнал, а постсинаптическая мембрана — часть клетки, получающей сигнал.
Медиатор высвобождается из булавовидного расширения посредством экзоцитоза, проходит через синаптическую щель и связывается с рецепторами на постсинаптической мембране. Под постсинаптической мембраной расположена субсинаптическая активная зона, в которой после активации рецепторов постсинаптической мембраны происходят разнообразные биохимические процессы.
В булавовидном расширении расположены содержащие медиаторы синаптические пузырьки, а также большое количество митохондрий и цистерны гладкой эндоплазматической сети. Применение традиционных методик фиксации при исследовании клеток позволяет различить на пресинаптической мембране пресинаптические уплотнения, ограничивающие активные зоны синапса, к которым при помощи микротрубочек направляются синаптические пузырьки.
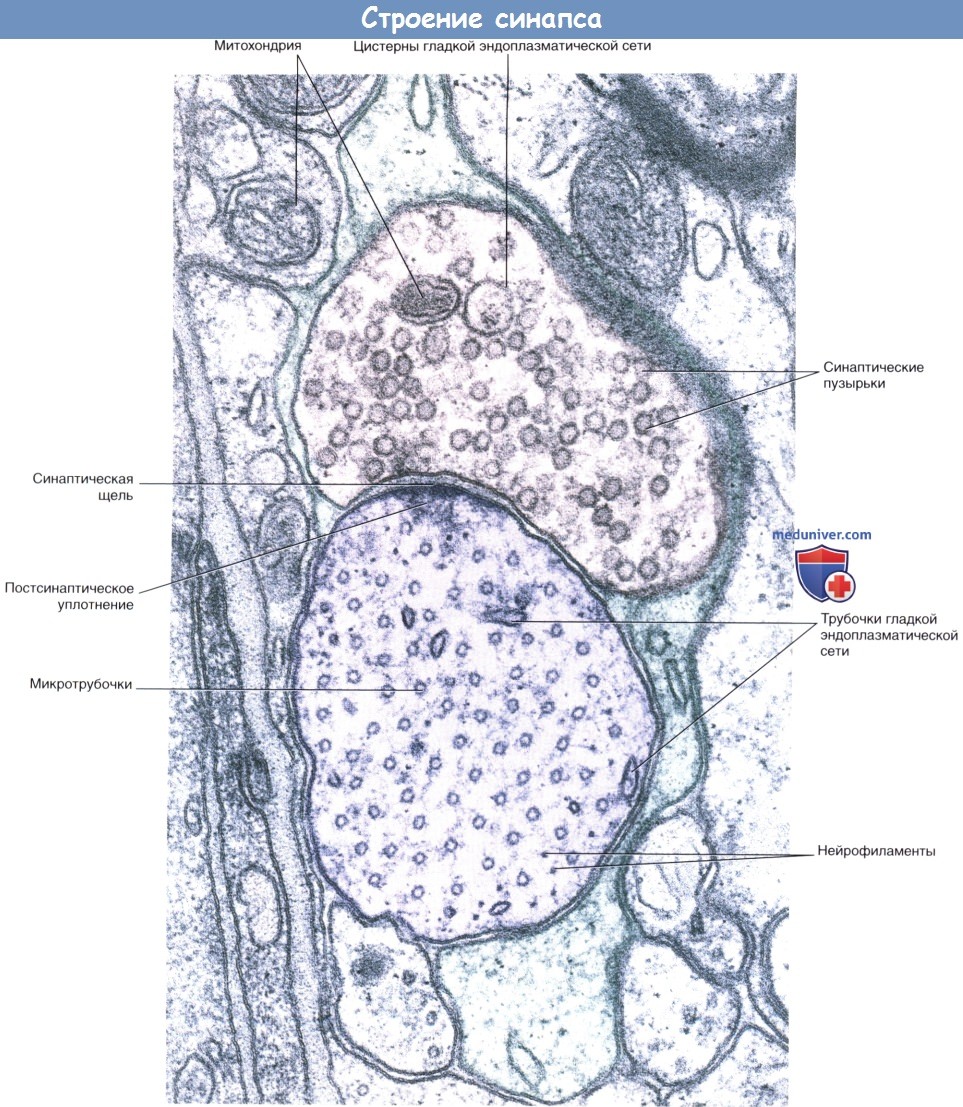
Аксодендритический синапс.
Срез препарата спинного мозга: синапс между концевым участком дендрита и, предположительно, двигательным нейроном.
Наличие округлых синаптических пузырьков и постсинаптического уплотнения характерно для возбуждающих синапсов.
Срез дендрита проведен в поперечном направлении, о чем свидетельствует наличие множества микротрубочек.
Кроме того, видны некоторые нейрофиламенты. Участок синапса окружен протоплазматическим астроцитом. 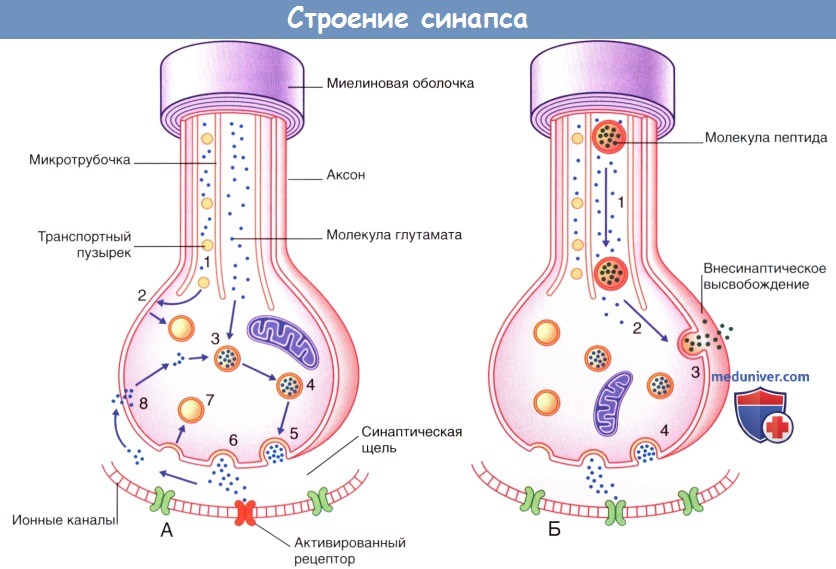
Процессы, происходящие в нервных окончаниях двух типов.
(А) Синаптическая передача небольших молекул (например, глутамата).
(1) Транспортные пузырьки, содержащие мембранные белки синаптических пузырьков, направляются вдоль микротрубочек к плазматической мембране булавовидного утолщения.
В это же время происходит перенос молекул ферментов и глутамата путем медленного транспорта.
(2) Мембранные белки пузырьков выходят из плазматической мембраны и формируют синаптические пузырьки.
(3) Глутамат погружается в синаптические пузырьки; происходит накопление медиатора.
(4) Пузырьки, содержащие глутамат, подходят к пресинаптической мембране.
(5) В результате деполяризации происходит экзоцитоз медиатора из частично разрушенных пузырьков.
(6) Высвобождающийся медиатор распространяется диффузно в области синаптической щели и активирует специфические рецепторы на постсинаптической мембране.
(7) Мембраны синаптических пузырьков транспортируются обратно в клетку путем эндоцитоза.
(8) Происходит частичный обратный захват глутамата в клетку для повторного использования.
(Б) Передача нейропептидов (например, субстанции Р), осуществляющаяся одновременно с синаптической передачей (например, глутамата).
Совместная передача этих веществ происходит в центральных нервных окончаниях униполярных нейронов, обеспечивающих болевую чувствительность.
(1) Синтезированные в комплексе Гольджи (в области перикариона) пузырьки и предшественники пептидов (пропептиды) транспортируются к булавовидному расширению путем быстрого транспорта.
(2) При их попадании в область булавовидного утолщения завершается процесс формирования молекулы пептида, и пузырьки транспортируются к плазматической мембране.
(3) Деполяризация мембраны и перенос содержимого пузырьков в межклеточное пространство путем экзоцитоза.
(4) Одновременно с этим происходит высвобождение глутамата.
1. Активация рецепторов. Молекулы медиаторов проходят через синаптическую щель и активируют рецепторные белки, расположенные парами на постсинаптической мембране. Активация рецепторов запускает ионные процессы, которые приводят к деполяризации постсинаптической мембраны (возбуждающее постсинаптическое действие) или гиперполяризации постсинаптической мембраны (тормозящее постсинаптическое действие). Изменение электротонуса передается в сому в виде затухающего по мере распространения электротонического потенциала, за счет которого происходит изменение потенциала покоя в начальном сегменте аксона.
Наиболее распространенный возбуждающий медиатор ЦНС — глутамат, а тормозной — гамма-аминомасляная кислота (ГАМК). В периферической нервной системе медиатором для двигательных нейронов поперечно-полосатой мускулатуры служит ацетилхолин, а для чувствительных нейронов — глутамат.
Последовательность процессов, происходящих в глутаматергических синапсах, показана на рисунке ниже. При передаче глутамата совместно с другими пептидами высвобождение пептидов осуществляется внесинаптическим путем.
Большинство чувствительных нейронов помимо глутамата выделяет и другие пептиды (один или несколько), высвобождающиеся в различных участках нейрона; однако основная функция этих пептидов — модуляция (повышение или снижение) эффективности синаптической передачи глутамата.
Кроме того, нейротрансмиссия может происходить путем диффузной внесинаптической передачи сигнала, характерной для моноаминергических нейронов (нейронов, использующих биогенные амины для обеспечения нейротрансмиссии). Выделяют две разновидности моноаминергических нейронов. В одних нейронах осуществляется синтез катехоламинов (норадреналина или дофамина) из аминокислоты тирозина, а в других — серотонина из аминокислоты триптофана. Например, дофамин высвобождается как в синаптической области, так и из варикозных утолщений аксона, в которых также происходит синтез этого нейромедиатора.
Дофамин проникает в межклеточную жидкость ЦНС и до момента деградации способен активировать специфические рецепторы на расстоянии до 100 мкм. Моноаминергические нейроны присутствуют во многих структурах ЦНС; нарушение передачи импульса этими нейронами приводит к различным заболеваниям, среди которых выделяют болезнь Паркинсона, шизофрению и глубокую депрессию.
Оксид азота (газообразная молекула) также участвует в диффузной нейропередаче в глутаматергической системе нейронов. Избыточное влияние оксида азота оказывает цитотоксическое действие, особенно в тех участках, кровоснабжение которых нарушено за счет тромбоза артерий. Глутамат также является потенциально цитотоксическим нейромедиатором.
Существует три вида химических межнейрональных взаимодействий: синаптическое (например, глутаматергическое), внесинаптическое (пептидергическое) и диффузное (например, моноаминергическое, серотонинергическое).
Химические синапсы классифицируют по анатомическому строению на аксодендритические, аксосоматические, аксоаксональные и дендро-дендритические. Синапс представлен пре- и постсинаптическими мембранами, синаптической щелью и субсинаптической активной зоной.
Электрические синапсы обеспечивают одновременную активацию целых групп нейронов, образуя между ними электрические связи за счет щелевидных контактов (нексусов).
![]()
Диффузная нейротрансмиссия в головном мозге.
Аксоны глутаматергического (1) и дофаминергического (2) нейронов образуют плотные синаптические контакты с отростком звездчатого нейрона (3) полосатого тела.
Дофамин высвобождается не только из пресинаптической области, но и из варикозного утолщения аксона, откуда диффузно распространяется в межклеточное пространство и активирует дофаминовые рецепторы дендритного ствола и стенки перицита капилляра. 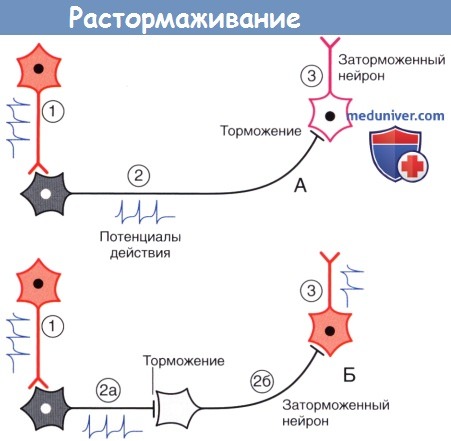
Растормаживание.
(А) Возбуждающий нейрон 1 активирует тормозной нейрон 2, который в свою очередь затормаживает нейрон 3.
(Б) Появление второго тормозного нейрона (2б) оказывает противоположное влияние на нейрон 3, поскольку происходит торможение нейрона 2б.
Спонтанно-активный нейрон 3 генерирует сигналы в условиях отсутствия тормозных влияний.
3. Торможение и растормаживание. Функционирование спонтанно-активных нейронов сдерживается под влиянием тормозных нейронов (обычно, ГАМКергических). Деятельность тормозных нейронов, в свою очередь, может быть ингибирована воздействующими на них другими тормозными нейронами, в результате чего происходит растормаживание клетки-мишени. Процесс растормаживания — важная особенность нейрональной активности в базальных ганглиях.
4. Редкие виды химических синапсов. Выделяют два типа аксоаксональных синапсов. В обоих случаях булавовидное утолщение образует тормозной нейрон. Синапсы первого типа образуются в области начального сегмента аксона и передают мощное ингибирующее влияние тормозного нейрона. Синапсы второго типа образуются между булавовидным утолщением тормозного нейрона и булавовидными утолщениями возбуждающих нейронов, что приводит к угнетению высвобождения медиаторов. Этот процесс получил название пресинаптического торможения. В этом плане традиционный синапс обеспечивает постсинаптичсекое торможение.
Дендро-дендритические (Д-Д) синапсы образуются между дендритными шипиками дендритов смежных шипиковых нейронов. Их задача — не генерирование нервного импульса, а изменение электротонуса клетки-мишени. В последовательных Д-Д-синапсах синаптические пузырьки располагаются только в одном дендритном шипике, а в реципрокном Д-Д-синапсе— в обоих. Возбуждающие Д-Д-синапсы изображены на рисунке ниже. Тормозные Д-Д-синапсы широко представлены в переключающих ядрах таламуса.
Кроме того, выделяют немногочисленные сомато-дендритические и сомато-соматические синапсы.
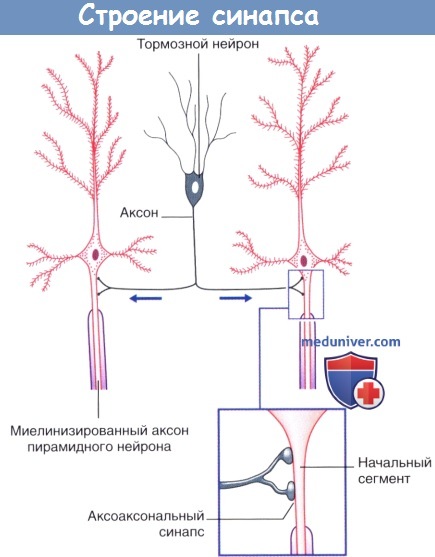
Аксоаксональные синапсы коры головного мозга.
Стрелками указано направление проведения импульсов. 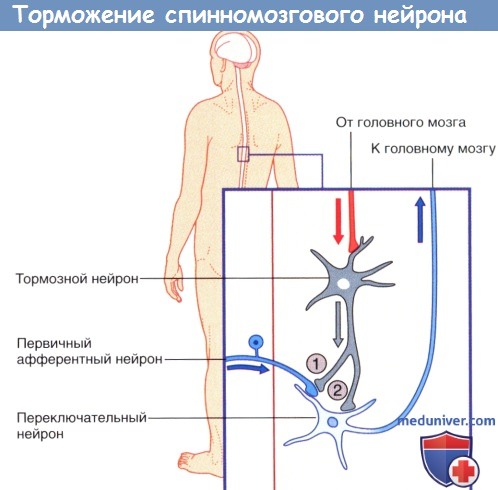
(1) Пресинаптическое и (2) постсинаптическое торможение спинномозгового нейрона, направляющегося к головному мозгу.
Стрелками указано направление проведения импульсов (возможно торможение переключательного нейрона под действием тормозных влияний). 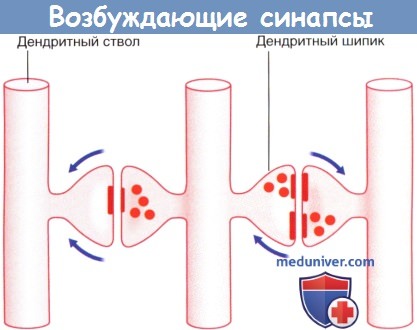
Возбуждающие дендро-дендритические синапсы. Изображены дендриты трех нейронов.
Реципрокный синапс (справа). Стрелками указано направление распространения электрото-нических волн.
Редактор: Искандер Милевски. Дата публикации: 11.11.2018
Синапс – это определенная зона контакта отростков нервных клеток и остальных невозбудимых и возбудимых клеток, которые обеспечивают передачу информационного сигнала. Синапс морфологически образуется контактирующими мембранами 2-х клеток. Мембрана, относящаяся к отростку нервных клеток, зовется пресинаптической мембраной клетки, в которую поступает сигнал, второе ее название – постсинаптическая. Вместе с принадлежностью постсинаптической мембраны синапс может быть межнейрональным, нейромышечным и нейросекреторным. Слово синапс было введено в 1897 г. Чарльзом Шеррингтоном (англ. физиологом).

Что же такое синапс?
Синапс – это специальная структура, которая обеспечивает передачу от нервного волокна нервного импульса на другое нервное волокно или нервную клетку, а чтобы произошло воздействие на нервное волокно от рецепторной клетки (области соприкосновения друг с другом нервных клеток и другого нервного волокна), требуется две нервные клетки.
Синапс – это небольшой отдел в окончании нейрона. При его помощи идет передача информации от первого нейрона ко второму. Синапс находится в трех участках нервных клеток. Также синапсы находятся в том месте, где нервная клетка вступает в соединение с разными железами или мышцами организма.
Из чего состоит синапс
Строение синапса имеет простую схему. Он образуется из 3-х частей, в каждой из которых осуществляются определенные функции во время передачи информации. Тем самым такое строение синапса можно назвать подходящим для передачи нервного импульса. Непосредственно на процесс передачи информации воздействуют две главные клетки: воспринимающая и передающая. В конце аксона передающей клетки находится пресинаптическое окончание (начальная часть синапса). Оно может повлиять в клетке на запуск нейротрансмиттеров (это слово имеет несколько значений: медиаторы, посредники или нейромедиаторы) – определенные химические вещества, с помощью которых между 2-мя нейронами реализуется передача электрического сигнала.

Медиаторы синапса
Медиатор (от латинского Media - передатчик, посредник или середина). Такие медиаторы синапса очень важны в процессе передачи нервного импульса.
Морфологическое различие тормозного и возбуждающего синапса заключается в том, что они не имеют механизм освобождения медиатора. Медиатор в тормозном синапсе, мотонейроне и другом тормозном синапсе считается аминокислотой глицином. Но тормозной или возбуждающий характер синапса определяется не их медиаторами, а свойством постсинаптической мембраны. К примеру, ацетилхолин дает возбуждающее действие в нервно-мышечном синапсе терминалей (блуждающих нервов в миокарде).
Ацетилхолин служит возбуждающим медиатором в холинэргических синапсах (пресинаптическую мембрану в нем играет окончание спинного мозга мотонейрона), в синапсе на клетках Рэншоу, в пресинаптическом терминале потовых желез, мозгового вещества надпочеников, в синапсе кишечника и в ганглиях симпатической нервной системы. Ацетилхоли-нестеразу и ацетилхолин нашли также во фракции разных отделов мозга, иногда в большом количестве, но кроме холинэргического синапса на клетках Рэншоу пока не смогли идентифицировать остальные холинэргические синапсы. По словам ученых, медиаторная возбуждающая функция ацетилхолина в ЦНС весьма вероятна.

Когда завершают свою функцию медиаторы синапса, катехоламин поглощается пресинаптическим нервным окончанием, при этом включается трансмембранный транспорт. Во время поглощения медиаторов синапсы находятся под защитой от преждевременного истощения запаса на протяжении долгой и ритмичной работы.
Синапс: основные виды и функции
Лэнгли в 1892 году было предположено, что синаптическая передача у вегетативной ганглии млекопитающих не электрической природы, а химической. Через 10 лет Элиоттом было выяснено, что из надпочечников адреналин получается от того же воздействия, что и стимуляция симпатических нервов.
Синапс химический
Химический синапс принципиально отличается передачей раздражения при помощи медиатора с пресинапса на постсинапс. Поэтому и образуются различия в морфологии химического синапса. Химический синапс более распространен в позвоночной ЦНС. Теперь известно, что нейрон способен выделять и синтезировать пару медиаторов (сосуществующих медиаторов). Нейроны тоже имеют нейромедиаторную пластичность – способность изменять главный медиатор во время развития.

Нервно-мышечный синапс
Данный синапс осуществляет передачу возбуждения, однако эту связь могут разрушить различные факторы. Передача заканчивается во время блокады выбрасывания в синаптическую щель ацетилхолина, также и во время избытка его содержания в зоне постсинаптических мембран. Множество ядов и лекарственных препаратов влияют на захват, выход, который связан с холинорецепторами постсинаптической мембраны, тогда мышечный синапс блокирует передачу возбуждения. Организм гибнет во время удушья и остановки сокращения дыхательных мышц.
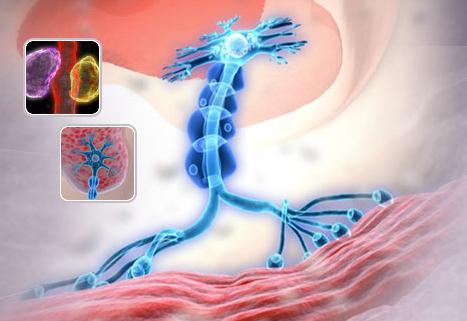
Синапс нервный: особенности и компоненты
Синапс – это соединение места контакта среди двух клеток. Причем каждая из них заключена в свою электрогенную мембрану. Нервный синапс состоит из трех главных компонентов: постсинаптическая мембрана, синаптическая щель и пресинаптическая мембрана. Постсинаптическая мембрана – это нервное окончание, которое проходит к мышце и опускается внутрь мышечной ткани. В пресинаптической области имеются везикулы – это замкнутые полости, имеющие медиатор. Они всегда находятся в движении.

Синаптическая щель среди пост- и пресинаптических мембран
В разных синапсах величина щели различна. Это пространство наполнено межклеточной жидкостью, в которой имеется медиатор. Постсинаптическая мембрана накрывает место контакта нервного окончания с иннервируемой клеткой в мионевральном синапсе. В определенных синапсах постсинаптическая мембрана создает складку, возрастает контактная площадь.
Дополнительные вещества, входящие в состав постсинаптической мембраны
В зоне постсинаптической мембраны присутствуют следующие вещества:
- Рецептор (холинорецептор в мионевральном синапсе).
- Липопротеин (обладает большой схожестью с ацетилхолином). У этого белка присутствует электрофильный конец и ионная головка. Головка поступает в синаптическую щель, происходит взаимодействие с катионовой головкой ацетилхолина. Из-за этого взаимодействия идет изменение постсинаптической мембраны, затем происходит деполяризация, и раскрываются потенциально зависимые Na-каналы. Деполяризация мембраны не считается самоподкрепляющим процессом;
- Градуален, его потенциал на постсинаптической мембране зависит от числа медиаторов, то есть потенциал характеризуется свойством местных возбуждений.
- Холинэстераза – считается белком, у которого имеется ферментная функция. По строению она схожа с холинорецептором и обладает похожими свойствами с ацетилхолином. Холинэстеразой разрушается ацетилхолин, вначале тот, который связан с холинорецептором. Под действием холинэстеразы холинорецептор убирает ацетилхолин, образуется реполяризация постсинаптической мембраны. Ацетилхолином расщепляется до уксусной кислоты и холина, необходимого для трофики мышечной ткани.
При помощи действующего транспорта выводится на пресинаптическую мембрану холин, он используется для синтеза нового медиатора. Под воздействием медиатора меняется проницаемость в постсинаптической мембране, а под холинэстеразой чувствительность и проницаемость возвращается к начальной величине. Хеморецепторы способны вступать во взаимодействие с новыми медиаторами.
Читайте также: