
Интраорганная нервная система это
А. Общая характеристика.
1. Внутренние органы после экстраорганной денервации -перерезки симпатических и парасимпатических нервов -продолжают функционировать. Вначале наблюдаются некоторые дистрофические явления, которые затем исчезают. Через несколько недель чувствительность внутренних органов к биологически активным веществам, и в первую очередь к медиаторам, повышается, их функция улучшается. Многие внутренние органы продолжают функционировать, будучи изолированными, находясь в физиологическом растворе: сокращаются перфузируемое сердце, мочеточники, желчный пузырь, кишечник. Причем, в пищеварительном тракте после перерезки симпатических и парасимпатических путей сохраняются все виды двигательной активности: перистальтика, ритмическая сегментация, маятникообразные хвижения, продолжается всасывание. Двигательная активность пищеварительных органов после перерезки их нервов регулируется рефлекторными дугами, замыкающимися в пределах мышечного и подслизистого сплетений его стенок, а также в экстраорганных ганглиях. Возбуждение афферентных нейронов кишечной стенки под действием пищевого комка приводит к возникновению рефлексов, при которых пищевой комок продвигается в каудальном направлении путем сокращения мускулатуры орального участка кишки и одновременного расслабления каудального участка. Дуги этих двух рефлексов, обусловливающих перистальтику, лежат в пределах стенки кишечника. Тормозные нейроны этих дуг не являются ни холинергическими, ни адренергическими, возможно, их медиатором служит АТФ. Возбуждающие нейроны выделяют ацетилхолин, имеются основания полагать, что их действие может быть опосредовано и другими медиаторами, например серотонином. Имеются спонтанно активные эфферентные нейроны, на них могут конвергировать интра- и экстраорганные нервные волокна.
 |
2. Интраорганная нервная система содержит все элементы рефлекторной дуги: афферентный, вставочный и эффекторный нейроны. Чувствительные интрамуральные нейроны - это клетки Догеля II типа, они образуют первое звено - рецептор и второе -афферентный путь рефлекторной дуги. Показано наличие механо-, хемо- и термочувствительных клеток. Обнаружены быстро и медленно адаптирующиеся тонические нейроны, возбуждающие и тормозящие сокращения кишки.
3. На одну и ту же клетку интрамуральных ганглиев конвергируют межганглионарные (вставочные) и экстраорганные - постганглионарные симпатические и преганглионарные парасимпатические волокна. Парасимпатические преганглионарные волокна образуют синапсы на возбуждающих и, по-видимому, тормозящих нейронах. Симпатические постганглионарные волокна оказывают непосредственное тормозное влияние на гладкую мышцу кишки, а также тормозят холинергические ганглионарные нейроны, угнетают выделение медиатора из пресинаптических окончаний преганглионарных парасимпатических волокон. Местные рефлекторные дуги выявлены и в сердце (Г. И. Косицкий и сотр.). Плотность расположения интраорганных нейронов весьма высока - в кишечнике, например, приходится около 20 тыс. нейрсг-нов на 1 см 2 . В целом в кишке число нейронов (1.-10 8 ) превосходит таковое в спинном мозге.
Б. В интраорганной нервной системе богат набор медиаторов и соответствующих рецепторов. Только в энтераль-ной ее части функционирует около 20 различных медиаторов и модуляторов. Наиболее изученные: ацетилхолин, катехоламины, серотонин, ГАМК, пептиды, АТФ.
ПАРАСИМПАТИЧЕСКАЯ НЕРВНАЯ СИСТЕМА
Парасимпатические нервные волокна имеются в черепных нервах (III пара - мезэнцефальный отдел, VII, IX и X пары — бульбарный отдел) и в тазовом нерве — сакральный отдел спинного мозга (S2—S4). Парасимпатические волокна III -й пары (глазодвигательный нерв) иннервируют глазные мышцы, регулируя диаметр зрачка и степень аккомодации. Парасимпатические веточки VII пары (лицевой нерв): иннервируют слизистую оболочку носа, неба, слезную железу; и секреторные клетки подчелюстной и подъязычной слюнных желез. Парасимпатические секреторные волокна IX пары (языкоглоточный нерв) подходят к околоушной железе. X пара (блуждающий нерв) своими ветвями снабжает дыхательные органы, большую часть пищеварительного тракта (до нисходящей ободочной кишки), сердце, печень, поджелудочную железу, почки. Парасимпатические нервы сакрального отдела спинного мозга (S2—S4) иннервируют нисходящую часть ободочной кишки и тазовые органы (прямую кишку, мочевой пузырь, половые органы). Парасимпатической иннервации не имеют скелетные мышцы, матка, мозг, подавляющее большинство кровеносных сосудов (кожи, органов брюшной полости, мышц), органы чувств и мозговое вещество надпочечников.
Парасимпатические ганглиии отдельные нейроны расположены внутри органов, а в тазовой области и в области головы — в непосредственной близости от органов. Отнервных клеток парасимпатических ганглиев идут короткие постганглионарные парасимпатические волокна, иннервирующие все перечисленные органы; преганглионарные волокна обычно длинные (у симпатической нервной системы, наоборот, преганглионарные короткие, постганглионарные длинные).
Медиаторы и рецепторы.Передача возбуждения с преганглионарногопарасимпатического волокна на эффекторный нейрон осуществляется с помощью ацетилхолина. Медиатор действует на Н-холинорецепторы постсинаптической мембраны ганглионарного нейрона.
Постганглионарное волокно свое влияние на эффекторную клетку передает также с помощью ацетилхолина. Действует на ионотропные Н-холинорецепторы и метаботропные М-холинорецепторы.
Внутренние органы после экстраорганной денервации — перерезки симпатических и парасимпатических нервов, продолжают функционировать. Вначале наблюдаются незначительные дистрофические явления, которые скоро исчезают. Через несколько недель чувствительность внутренних органов к биологически активным веществам и в первую очередь к медиаторам повышается, их функция улучшается. Многие внутренние органы продолжают функционировать, будучи изолированными, находясь в физиологическом растворе: сокращаются перфузируемое сердце, мочеточники, желчный пузырь, кишечник. В пищеварительном тракте после перерезки симпатических и парасимпатических путей сохраняются все виды двигательной активности: перистальтика, ритмическая сегментация, маятникообразные движения, продолжается всасывание. Двигательная активность пищеварительных органов после перерезки их нервов регулируется рефлекторными дугами, замыкающимися в пределах межмышечного и подслизистого сплетений его стенок, а также в экстраорганных ганглиях.
Интраорганная нервная система содержит все элементы рефлекторной дуги: афферентный, вставочный и эффекторный нейроны. Чувствительные интрамуральные нейроны — клетки II типа Догеля; они образуют первое звено — рецептор, и второе — афферентный путь рефлекторной дуги. Показано наличие механо-, хемо- и термочувствительных клеток. Обнаружены быстро- и медлен:но адаптирующиеся тонические нейроны,возбуждающие и тормозящие сокращения кишки.
В интраорганной нервной системе богат набор медиаторов и соответствующих рецепторов.Только в энтеральной ее части функционирует около 20 различных медиаторов и модуляторов. Наиболее изучены ацетилхолин, катехоламины, серотонин, ГАМК, пептиды, АТФ.
| | | следующая лекция ==> | |
| СИМПАТИЧЕСКАЯ НЕРВНАЯ СИСТЕМА | | | Центры автономной нервной системы |
Нам важно ваше мнение! Был ли полезен опубликованный материал? Да | Нет
Интраорганная нервная система. Многие внутренние органы, такие как сердце, мочеточники, желчный пузырь, кишечник, продолжают функционировать даже в изолированном от организма состоянии. Такую возможность им обеспечивает интраорганная нервная система – часть вегетативной нервной системы. Интраорганная нервная система имеет и другое название – метасимпатическая часть вегетативной нервной системы. Эта часть образована совокупностью микроганглионарных образований, находящихся в стенках внутренних органов (сердце, кишечник и т.д.). Данные образования обладают моторной активностью и имеют ряд особенностей:
1.
Находятся только во внутренних органах, которые имеют свою собственную моторную активность;
2.
Не вступают в антагонистические отношения с другими частями нервной системы;
3.
Обладают независимостью от ЦНС;
4.
Обладает холин-, адрен- и пуринергической видами передачи.
Рефлекторные дуги интраорганной нервной системы содержат в себе следующие нейроны: афферентные, вставочные и эффекторные. Чувствительные интрамуральные нейроны (афферентные) – клетки Догеля II типа образуют первое звено – рецептор, и второе – афферентный путь рефлекторной дуги. Двигательными являются клетки Догеля Iтипа (эфферентные), которые образуют эфферентный путь. Клетки интраорганной нервной системы могут быть механо-, хемо- и термочувствительными.
Интраорганная нервная система выполняет роль фактора надежности регуляторных механизмов организма.
Δ Медиаторы интраорганной нервной системы. Медиаторами интраорганной нервной системы являются более 20 видов химических веществ, к которым относят: серотонин, вазоактивный интестинальный пептид (ВИП), гистамин, дофамин, вещество Р и т.д.
■ Взаимодействие между отделами ВНС. Взаимодействие между отделами ВНС осуществляется следующими путями:
1.
В основном синергизмом противоположных симпатических и парасимпатических влияний;
2.
Взаимодействием в центральном отделе;
3.
Взаимодействием в периферическом отделе.
4.
Иногда однонаправленным синергичным влиянием на один и тот же орган;
Рефлекторные дуги интраорганной нервной системы содержат все необходимые для осуществления рефлексов элементы: афферентный, вставочный и эффекторный нейроны. Чувствительные интрамуральные нейроны – клетки Догеля II типа; они образуют первое звено – рецептор, и второе – афферентный путь рефлекторной дуги, двигательными являются клетки Догеля I типа. Обнаружены быстро- и медленно адаптирующиеся тонические нейроны, активирующие и тормозящие сокращения органа. Имеются механо-, хемо- и термочувствительные клетки.
Медиаторы и рецепторы интраорганной нервной системывесьма разнообразны: в энтеральной ее части функционирует около 20 различных медиаторов и модуляторов.
Серотонин обнаружен в различных тканях организма животных. Однако больше всего серотонина (90 % от его общего количества, содержащегося в организме) приходится на энтерохро-маффинные клетки пищеварительного тракта. Серотонин вызывает сокращение гладких мышц внутренних органов, в том числе кровеносных сосудов и бронхов. При прямом действии серотонина возрастает сила сердечных сокращений, хотя этот эффект маскируется рефлекторными барорецептивными влияниями противоположного направления. В центральном и периферическом отделах нервной системы выявлено несколько типов серотонино-рецепторов: 5-НТ1–5-НТ5 (S1–S5). Имеются не только пост-, но и пресинаптические серотонинорецепторы. S1,2-рецепторы находятся в гладкой мышце желудочно-кишечного тракта, S3-peцепторы – в вегетативных ганглиях, S4,5-рецепторы обнаружены в ЦНС.
Медиаторная роль пуринов. Считают, что в интрамуральном нервном аппарате имеются пуринергические нейроны, аксоны которых выделяют продукты распада АТФ (например, аденозин). АТФ и сама оказывает тормозящее влияние на моторику желудочно-кишечного тракта. Выделено два типа пуринорецепторов (Р1 и Р2); P1 более чувствительны к аденозину, Р2 –к АТФ. Р1-рецепторы преобладают в сердечно-сосудистой системе, трахее, мозге. Они связаны со специфической аденилатциклазой. Р2-рецепторы расположены в основном на клетках органов желудочно-кишечного тракта и мочеполовой системы. Активация пуринергических рецепторов вызывает расслабление кишки, а возникающие вслед за этим ее сокращения под влиянием холинергических нейронов обеспечивают прохождение содержимого по кишке (пропульсия). Пуринергические нейроны участвуют также в механизме релаксации желудка, расслаблении пищеводного и анального сфинктеров. Тормозящее (гиперполяризующее) влияние АТФ реализуется, по-видимому, через повышение проницаемости клеточных мембран для ионов калия.
Гистамин довольно широко распространен в организме. Пост-синаптические гистаминорецепторы (подтипы H1 и Н2) имеются на клетках всех органов и участвуют в регуляции различных функций организма (табл. 1). Н3-рецепторы, обнаруженные в ЦНС на пресинапсах, по-видимому регулируют выделение ими гистамина. Антагонистами Н1-рецепторов являются супрастин, дипразин (пипольфен), Н2-рецепторов – цимитидин.
Таблица 1. Эффекты активации гистаминовых рецепторов.
| Тип рецепторов | Локализация рецепторов | Эффекты |
| Н1 | Бронхи, вены, крупные артерии, мочевой и желчный пузыри, кишечник, желудок | Повышение тонуса, сокращение |
| Сердце | Угнетение атриовентрикулярной проводимости, усиление сокращений предсердий | |
| Мелкие артерии и коронарные сосуды | Расслабление (Н2–то же) | |
| Капилляры | Повышение проницаемости | |
| Н2 | Париетальные клетки желудка | Повышение секреции НС1 |
| Слюнные железы | Повышение секреции | |
| Сердце | Увеличение частоты и силы сокращений | |
| Матка | Расслабление | |
| Бронхи, желчный пузырь, мочевой пузырь, кишечник | Расслабление (мало) |
Дофамин. Установлено два типа дофаминорецепторов: тип D, выявлен в симпатических ганглиях, паращитовидных железах. По-видимому, все дофаминрецепторы являются постсинаптическими. D2-рецепторы найдены на мембранах лимфоцитов. D1- и D2-рецепторы более широко распространены в ЦНС.
ВИП – вазоактивный пептид, который через образование цАМФ вызывает усиление моторной и секреторной активности, а также кровоснабжения желудочно-кишечного тракта.
Вещество Р. Рецепторы к веществу Р имеются на гладкомы-шечных клетках кишки, клетках семявыносящих протоков, мочевого пузыря.
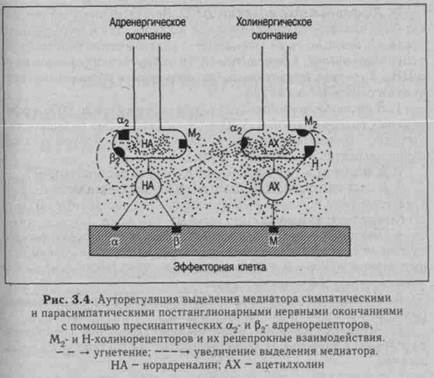
Экстраорганную иннервацию имеют М-холино-, α1- и β1-ад-ренорецепторы, серотонино- и пуринорецепторы. Степень иннервации других рецепторов изучена мало. Активность синапсов экстра- и интраорганной нервной системы регулируется различными способами.
Дата добавления: 2014-11-24 ; просмотров: 35 | Нарушение авторских прав
А. Общая характеристика.
1. Внутренние органы после экстраорганной денервации -перерезки симпатических и парасимпатических нервов -продолжают функционировать. Вначале наблюдаются некоторые дистрофические явления, которые затем исчезают. Через несколько недель чувствительность внутренних органов к биологически активным веществам, и в первую очередь к медиаторам, повышается, их функция улучшается. Многие внутренние органы продолжают функционировать, будучи изолированными, находясь в физиологическом растворе: сокращаются перфузируемое сердце, мочеточники, желчный пузырь, кишечник. Причем, в пищеварительном тракте после перерезки симпатических и парасимпатических путей сохраняются все виды двигательной активности: перистальтика, ритмическая сегментация, маятникообразные хвижения, продолжается всасывание. Двигательная активность пищеварительных органов после перерезки их нервов регулируется рефлекторными дугами, замыкающимися в пределах мышечного и подслизистого сплетений его стенок, а также в экстраорганных ганглиях. Возбуждение афферентных нейронов кишечной стенки под действием пищевого комка приводит к возникновению рефлексов, при которых пищевой комок продвигается в каудальном направлении путем сокращения мускулатуры орального участка кишки и одновременного расслабления каудального участка. Дуги этих двух рефлексов, обусловливающих перистальтику, лежат в пределах стенки кишечника. Тормозные нейроны этих дуг не являются ни холинергическими, ни адренергическими, возможно, их медиатором служит АТФ. Возбуждающие нейроны выделяют ацетилхолин, имеются основания полагать, что их действие может быть опосредовано и другими медиаторами, например серотонином. Имеются спонтанно активные эфферентные нейроны, на них могут конвергировать интра- и экстраорганные нервные волокна.

Интраорганная нервная система содержит все элементы рефлекторной дуги: афферентный, вставочный и эффекторный нейроны. Чувствительные интрамуральные нейроны - это клетки Догеля II типа, они образуют первое звено - рецептор и второе -афферентный путь рефлекторной дуги. Показано наличие механо-, хемо- и термочувствительных клеток. Обнаружены быстро и медленно адаптирующиеся тонические нейроны, возбуждающие и тормозящие сокращения кишки.
На одну и ту же клетку интрамуральных ганглиев конвергируют межганглионарные (вставочные) и экстраорганные - постганглионарные симпатические и преганглионарные парасимпатические волокна. Парасимпатические преганглионарные волокна образуют синапсы на возбуждающих и, по-видимому, тормозящих нейронах. Симпатические постганглионарные волокна оказывают непосредственное тормозное влияние на гладкую мышцу кишки, а также тормозят холинергические ганглионарные нейроны, угнетают выделение медиатора из пресинаптических окончаний преганглионарных парасимпатических волокон. Местные рефлекторные дуги выявлены и в сердце (Г. И. Косицкий и сотр.). Плотность расположения интраорганных нейронов весьма высока - в кишечнике, например, приходится около 20 тыс. нейрсг-нов на 1 см 2 . В целом в кишке число нейронов (1.-10 8 ) превосходит таковое в спинном мозге.
Б. В интраорганной нервной системе богат набор медиаторов и соответствующих рецепторов. Только в энтераль-ной ее части функционирует около 20 различных медиаторов и модуляторов. Наиболее изученные: ацетилхолин, катехоламины, серотонин, ГАМК, пептиды, АТФ.

Лекция 11 . Физиология вегетативной нервной системы
Автономная (вегетативная) нервная система – это часть нервной системы, регулирующая работу внутренних органов, просвет сосудов, обмен веществ и энергии, обеспечивающих гомеостаз. Деятельность автономной нервной системы не подчиняется сознанию человека. Вегетативная нервная система делится на две части:
1. Симпатическую систему, задача которой состоит в мобилизации организма на решение жизненно важных задач;
2. Парасимпатическую систему, задача которой состоит в восстановлении нарушенного во время активности организма гомеостаза и восполнении потраченных ресурсов.
Эффекты этих двух систем часто противоположный друг другу. Большинство систем внутренних органов получают волокна от обеих систем ВНС. Обе системы работают согласованно. Кроме этого в ВНС выделяют интраорганную нервную систему, которая располагается непосредственно в органах человека и оказывает регулирующее воздействие на эти органы. Она регулирует моторику и сократительную активность.
Δ Общий план строения ВНС. Вегетативная нервная система состоит из четырех уровней организации: периферического, промежуточного, центрального и высшего.
Таблица 1. Уровни организации ВНС
Уровень организации
Локализация нервных центров
Основные функции
Интрамуральные сплетения во внутренних органах
Местные рефлекторные реакции на уровне отдельных органов или их участков
Паравертебральные и превертебральные ганглии
Периферические рефлекторные реакции; координация работы отдельных участков органа или органов одной системы
Краниобульбарный отдел продолговатого мозга, поясничный и крестцовый отделы спинного мозга
Запуск реакций мобилизации ресурсов в экстремальных состояниях (симпатическая нс) или восстановление потраченных ресурсов (парасимпатическая нс)
Гипоталамус, мозжечок, базальные ганглии, кора больших полушарий
Координация деятельности симпатической и парасимпатической нс; адаптация внутренних процессов, протекающих в организме, к требованиям окружающей среды
Δ Нейронный состав ВНС :
1. Чувствительные нейроны располагаются в спинномозговых узлах.
2. Преганглионарные нейроны располагаются в центральной нервной системе: в стволе мозга (парасимпатические) или в спинном мозге (в грудном отделе – симпатические, в крестцовом – парасимпатические). Их отростки – преганглионарные волокна идут до соответствующих вегетативных ганглиев (симпатические - до паравертебральных и превертебральных, парасимпатические – до интрамуральных), где они заканчиваются синапсами на постганлионарных нейронах.
3. Постганглионарные нейроны расположены в вегетативных ганглиях. Ганглии могут находиться или на некотором расстоянии от органов (симпатические ганглии), либо непосредственно в стенке иннервируемого органа (парасимпатические ганглии).
Δ Рефлекторный принцип работы ВНС. Дуга вегетативной нервной системы построена по общему принципу, но имеются особенности её организации:
1. Рефлекторная дуга ВНС может замыкаться вне ЦНС. Эффекторный нейрон для симпатического отдела ВНС расположен экстраорганно – в симпатических ганглиях, а для парасимпатической – внутри органа или непосредственно вблизи него.
2. Афферентное звено дуги вегетативного рефлекса может быть образовано как собственными – вегетативными, так и соматическими афферентными волокнами.
3. В дуге вегетативного рефлекса слабее выражена сегментарность, что повышает надежность вегетативной иннервации.
Вегетативные рефлексы подразделяются на следующие группы:
1. Периферические рефлексы , которые подразделяют на интраорганные, экстраорганные и центральные (разделение по уровню замыкания рефлекторной дуги).
2. Висцеро-соматические рефлексы – изменение соматической деятельности при возбуждении сенсорных рецепторов ВНС.
3. Висцеро-висцеральные рефлексы – рефлексы, рецептивные поля которых локализуются в одном органе или разных органах, при этом ответные реакции проявляются в изменениях активности других внутренних органов.
4. Сомато-висцеральные рефлексы – изменение деятельности внутренних органов при раздражении соматических рецепторов. Афферентное звено этих рефлексов относится к соматической нервной системе, а эфферентное – к ВНС.
5. Интероцептивные: механо-, хемо-, термо-, осмо-, ноцицепторы; проприоцептивные и экстероцептивные : термо - и ноцицепторы, рецепторы органов чувств (деление по рецепторам рефлексогенных зон).
ΔФункциональные отличия вегетативной нервной системы от соматической нервной системы :
1. Регуляция функций внутренних органов с помощью ВНС может осуществляться при полном нарушении связи с ЦНС, но менее совершенно, так как эффекторные нейроны ВНС находятся за пределами ЦНС.
2. Влияние ВНС на органы и ткани организма не контролируется непосредственно сознанием.
3. Нейроны ВНС имеют невысокую лабильность и небольшую скорость проведения импульсов.
4. Влияния симпатического отдела ВНС имеют генерализованный (диффузный) характер.
■ Физиология симпатической нервной системы . Центральные образования симпатической части ВНС расположены в ядрах боковых рогов серого вещества спинного мозга (с I по XII грудных сегментов и со II по IV поясничных сегментов). Симпатические нервные волокна выходят из спинного мозга в составе передних корешков, а затем отделяются от них. При этом выделяются две части этих волокон:
· Первая часть преганглионарных волокон прерывается в узлах симпатического ствола (паравертебральных ганглиях). Паравертебральные ганлии расположены в виде цепочек по обе стороны вдоль позвоночника от основания черепа до копчика. Здесь эта часть волокон прерывается и переключается на тела вторых нейронов. Симпатические ганглии входят в состав следующих вегетативных нервных сплетений:
1. Сердечное сплетение расположено у основания сердца; иннервирует сердце и легкие.
2. Чревное сплетение (солнечное) лежит позади желудка; иннервирует органы брюшной полости.
3. Подчревное сплетение находится впереди крестца; иннервирует органы малого таза.
· Вторая часть преганглионарных волокон идет через паравертебральные узлы не прерываясь. Эти волокна заканчиваются в превертебральных ганглиях.
Волокна, выходящие из паравертебральных узлов, называются постганглионарными. Постганглионарные волокна (после выхода из узла) разделяются: одни в составе нервов соматической нервной системы достигают скелетных мышц, другие идут к внутренним органам.
Δ Функции симпатической нервной системы . Симпатическая нервная система иннервирует почти все органы: сердце, сосуды, бронхи, желудочно-кишечный тракт, органы мочеполовой системы, потовые железы, печень, мышцы зрачка, матку, надпочечники и другие эндокринные железы. Её задача состоит в мобилизации организма на решение жизненно важных задач, для чего необходимо осуществить следующие физиологические процессы:
· Расширить кровеносные сосуды скелетной мускулатуры;
· Вовлечь в общий кровоток депонированную кровь, находящуюся в селезенке и подкожной клетчатке;
· Обеспечить более интенсивную работу сердца путем повышения частоты и силы сердечных сокращений, а также улучшения коронарного кровотока;
· Уменьшить доступ крови к органам, деятельность которых не нужна в данный момент;
· Извлечь резервные запасы питательных веществ и перевести их в доступную для организма форму;
· Усилить легочную вентиляцию для большего поступления кислорода в организм.
Комплекс данных реакций объединяют в понятие эрготропная функция симпатической нервной системы.
Δ Медиаторы симпатической нервной системы . В преганглионарных волокнах, которые контактируют с постганглионарным нейроном, выделяется ацетилхолин, то есть волокна и сам преганглионарный нейрон являются холинергическими. Ацетилхолин взаимодействует с Н-холинорецепторами (никотин-чувствительные рецепторы), в результате чего происходит передача возбуждения с преганглионарного волокна на постганглионарный нейрон. Никотин в малых концентрациях возбуждает Н-холинорецепторы, в больших – тормозит. Постганглионарные волокна симпатической нервной системы являются адренергическими (в их окончаниях выделяется норадреналин). Исключением являются постганглионарные симпатические волокна потовых желез, где выделяется ацетилхолин, поэтому эти волокна называются симпатическими холинергическими. Реакция органа на выделение медиатора зависит не столько от самого медиатора, сколь от типа рецептора, с которым данный медиатор взаимодействует. Конечный эффект возбуждения симпатических волокон зависит от того, какая популяция адренорецепторов (ά1,ά2,β1,β2) преобладает в органе на постсинаптической мембране. Механизм влияния норадреналина на клетку может быть различным: норадреналин после взаимодействия с адренорецептором меняет проницаемость мембраны для ионов, что приводит либо к деполяризации (увеличение потока входящих в клетку ионов натрия) - возбуждению, либо к гиперполяризации (повышение проницаемости для ионов калия) – торможению.
■ Физиология парасимпатической нервной системы . Центральные (преганглионарные) нейроны парасипатичекой нервной системы расположены в среднем, продолговатом мозге и в люмбально-сакральном отделе спинного мозга.
→ В среднем мозге расположены два парасимпатических ядра, относящихся к III паре черепно-мозговых нервов, которые обеспечивают иннервацию сфинктера зрачка и ресничную мышцу глаза.
→ В продолговатом мозге имеются парасимпатические ядра VII , IX , X пар черепно-мозговых нервов:
· Ядро VII пары чмн иннервирует слизистые железы носа, слезную железу, две слюнные железы (подъязычную и подчелюстную).
· Ядро IX пары чмн иннервирует околоушную железу.
· Ядро X пары чмн иннервирует сердце, легкие, желудочно-кишечный тракт.
→ В пояснично-сакральном отделе спинного мозга расположены нейроны, образующие центры мочеиспускания, дефекации и эрекции.
От преганглионарных нейронов аксоны образуют преганглионарные волокна, которые идут к органу. Парасимпатические ганглии расположены внутри органов, а в области головы и тазовых органов – в непосредственной близости от них. От нервных клеток ганглиев отходят постганглионарные волокна. По длине они короче, чем преганглионарные. Постганглионарные волокна заканчиваются либо на клетках органа, либо действуют на орган через интраорганную систему.
Функции парасимпатической нервной системы можно охарактеризовать следующим образом: возбуждение парасимпатической системы приводит к восстановлению гомеостаза, то есть к тропотрофному эффекту.
→ Медиаторы парасимпатической нервной системы . В преганглионарных волокнах парасимпатической нервной системы медиатором является ацетилхолин. Ацетилхолин взаимодействует на постсинаптической мембране постганглионарного нейрона с Н-холинорецепторами. В окончаниях постганглионарных волокон парасимпатической нервной системы также выделяется ацетилхолин, а рецепторы, расположенные на постсинаптической мембране органа – это М-холинорецепторы (мускаринчувствительные холинорецепторы).
■ Интраорганная нервная система . Многие внутренние органы, такие как сердце, мочеточники, желчный пузырь, кишечник, продолжают функционировать даже в изолированном от организма состоянии. Такую возможность им обеспечивает интраорганная нервная система – часть вегетативной нервной системы. Интраорганная нервная система имеет и другое название – метасимпатическая часть вегетативной нервной системы. Эта часть образована совокупностью микроганглионарных образований, находящихся в стенках внутренних органов (сердце, кишечник и т. д.). Данные образования обладают моторной активностью и имеют ряд особенностей:
1. Находятся только во внутренних органах, которые имеют свою собственную моторную активность;
2. Не вступают в антагонистические отношения с другими частями нервной системы;
3. Обладают независимостью от ЦНС;
4. Обладает холин-, адрен- и пуринергической видами передачи.
Рефлекторные дуги интраорганной нервной системы содержат в себе следующие нейроны: афферентные, вставочные и эффекторные. Чувствительные интрамуральные нейроны (афферентные) – клетки Догеля II типа образуют первое звено – рецептор, и второе – афферентный путь рефлекторной дуги. Двигательными являются клетки Догеля Iтипа (эфферентные), которые образуют эфферентный путь. Клетки интраорганной нервной системы могут быть механо-, хемо - и термочувствительными.
Интраорганная нервная система выполняет роль фактора надежности регуляторных механизмов организма.
Δ Медиаторы интраорганной нервной системы . Медиаторами интраорганной нервной системы являются более 20 видов химических веществ, к которым относят: серотонин, вазоактивный интестинальный пептид (ВИП), гистамин, дофамин, вещество Р и т. д.
■ Взаимодействие между отделами ВНС . Взаимодействие между отделами ВНС осуществляется следующими путями:
1. В основном синергизмом противоположных симпатических и парасимпатических влияний;
2. Взаимодействием в центральном отделе;
3. Взаимодействием в периферическом отделе.
4. Иногда однонаправленным синергичным влиянием на один и тот же орган;
■ Высшие центры ВНС . К высшим нервным центрам вегетативной нервной системы относят: гипоталамус, моноаминергическую систему, кору и мозжечок.
1. Гипоталамус является одним из важнейших образований мозга, участвующих в регуляции активности нейронов парасимпатической и симпатической нервной системы. Кроме того он является компонентом лимбической системы, в которую входят многие структуры старой и древней коры больших полушарий, в том числе гиппокамп и миндалевидный комплекс. В гипоталамусе имеются центры регуляции следующих функций:
· Центры регуляции водного и солевого обмена;
· Центры белкового, углеводного и липидного обмена;
· Центры регуляции сердечно-сосудистой системы, эндокринных желез;
· Центр голода и насыщения;
· Центры жажды и отказа от питья;
· Центр регуляции мочеотделения;
· Центр сна и бодрствования;
· Центр полового поведения;
· Центры, обеспечивающие эмоциональные переживания человека.
Гипоталамус представляет собой скопление более чем 32 пар ядер. Наиболее значимые из них – это
1. Супрооптическое и паравентрикулярные ядра – участвуют в процессах регуляции водно-солевого обмена, лактации, активности матки, способны продуцировать гормоны окситоцин, антидиуретический гормон;
2. Аркуатное и вентромедиальные ядра – вырабатывают пептидные гормоны либерины (высвобождающие другие гормоны) и статины (тормозящие выработку других гормонов);
2. Моноаминергическая система объединяет в себе скопление нейронов ствола, аксоны которых идут в составе медиального пучка переднего мозга и достигают практически всех структур мозга, в том числе гипоталамуса, таламуса, базальных ганглиев и некоторых участков коры. Моноаминергическая система работает с такими медиаторами как норадреналин, серотонин и дофамин.
→Тела норадренергических нейронов расположены в продолговатом мозге и в мосту. Особенно их много в голубом пятне. Эти нейроны оказывают тормозящее действие на структуры ЦНС.
→Тела дофаминергических нейронов лежат в среднем мозге, в основном черной субстанции. Аксоны их связаны с базальными ганглиями и другими структурами мозга.
→Серотонинергические нейроны локализованы в продолговатом мозге, в основном в ядрах шва, в мосту и в нижних отделах среднего мозга.
Норадреналин создает условия для появления у человека положительных эмоций – благодаря норадреналину в основном функционируют центры удовольствия.
Дофамин также способствует созданию положительных эмоций.
Серотонин играет роль в создании отрицательных эмоций – при этом активируются центры избегания.
Моноаминергическая система играет важную роль в создании эмоций и вегетативного обеспечения эмоционального состояния. Это осуществляется за счет воздействия нейронов моноаминергической системы на структуры лимбической системы и на ядра гипоталамуса, которые управляют вегетативными нейронами ствола и спинного мозга.
3. Мозжечок играет важную роль в регуляции вегетативных функций, так как влияет на возбудимость вегетативных центров и тем самым способствует адаптации организма к выполнению двигательных актов. То есть, мозжечок представляет собой посредника между вегетативной и соматической нервной системой. Мозжечок влияет на деятельность сердца, меняет величину артериального давления, регионарный кровоток, влияет на глубину и частоту дыхания, на моторную, секреторную, всасывательную функции желудочно-кишечного тракта, на тонус мышц мочевого пузыря. При повреждении мозжечка нарушается углеводный, белковый и минеральный обмен, нарушаются энергообразование и процессы терморегуляции.
4. Кора больших полушарий оказывает значительное влияние на вегетативную нервную систему. Особую роль играет гиппокамп: он задействован не только в процессах памяти и эмоциональных реакциях, но и в процессах регулирования вегетативных реакций. При раздражении гиппокампа происходит активация парасимпатического отдела автономной нервной системы, изменяется деятельность сердечно-сосудистой системы, меняются частота и глубина дыхания.
■ Тонус автономной нервной системы . Для оценки состояния тонуса ВНС применяется анализ вариабельности сердечного ритма, который не просто показывает общий тонус ВНС, а может избирательно вычленять, выделять вклад симпатического и парасимпатического отдела в общую картину. Интегральным показателем тонуса ВНС можно считать индекс напряжения (ИН) регуляторных систем по , отражающий соотношение активности симпатического и парасимпатического отделов ВНС. Тонус ВНС делят на пять типов:
1. Ваготонический – до 30 ед;
2. Нормотонический – 31-120 ед;
3. Симпатикотонический – 121-300ед;
4. Сверхсимпатикотонический – 301-600ед;
5. Запредельный – более 600ед.
При стрессе тонус автономной нервной системы сдвигается в область симпатикотонии не только за счет усиления активности симпатической нервной системы, но и за счет существенного снижения активности блуждающего нерва. При этом ИН увеличивается в 2-3 раза, достигая 150-180 единиц вместо обычных 65-80 единиц. Однако, если человеку не удается адаптироваться к острому стрессу, то со временем наступает обратная картина – сдвиг равновесия в сторону парасимпатической системы, сопровождающийся запредельным торможением коры больших полушарий.
Читайте также: