
Как доказать что секреция пищеварительного сока поджелудочной железы регулируется нервным и гуморальным путями
Слабое сокоотделение натощак резко усиливается во время (через 2— 3 мин) и после приема пищи. Панкреатическое сокоотделение начинается уже на вид, запах пищи как условно-рефлекторный механизме секреции. При раздражении пищевыми веществами рецепторов слизистой оболочки ротовой полости, глотки и пищевода возникает безусловно-рефлекторное отделение панкреатического сока. Нервные импульсы от раздражаемых рецепторов по афферентным путям достигают бульбарного центра панкреатической секреции, где они переключаются на преганглионарные нейроны ядер блуждающего нерва, по эфферентным волокнам которого достигают постганглионарных нейронов. Их аксоны образуют синаптические окончания на базальных мембранах панкреацитов.
Выделяющийся при возбуждении этих окончаний ацетилхолин активирует М-холинорецепторы постсинаптической мембраны.
При раздражении чревных нервов, осуществляющих симпатическую иннервацию поджелудочной железы, ее сокоотделение тормозится (за счет активации р-адренорецепторов медиатором норадреналином). Но торможение секреции сопровождается накоплением секреторного материала в панкреацитах.
Поступление пищи в желудок во время приема пищи вызывает раздражение хемо- и механорецепторов желудка.
Поступление в двенадцатиперстную кишку желудочного содержимого вызывает вместе с дуоденопанкреатическим секреторным рефлексом (с рецепторов слизистой оболочки) выделение из эндокринных клеток гастро-интестинальных гормонов, стимулирующих или тормозящих сокоотделение поджелудочной железой.
Роль основных гуморальных регуляторов выполняют секретин и ХЦК, которые вырабатываются S- и ССК-клетками слизистой оболочки двенадцатиперстной кишки. Секретин и ХЦК усиливают влияние друг друга на панкреоциты, особенно на фоне выделения ацетилхолина в синаптических окончаниях холинергических нервных волокон, иннервирующих железу. Важным гуморальным возбудителем панкреатической секреции является гастрин.
Изменится ли амплитула потенциала действия нервной клетки, если на нее воздейсвовать стимулами сверхпороговой силы
Билет № 5
1.потенциал действия,с графиком, возбудимость,мера возбудимости,
Потенциал действия (ПД) – быстрое изменение мембранного потенциала в ответ на действия раздражителя пороговой силы. ПД имеет стандартные амплитуду и временные параметры, не зависящие от силы стимула - правило "ВСЕ ИЛИ НИЧЕГО".
Начальная деполяризация мембраны под действием раздражителя:
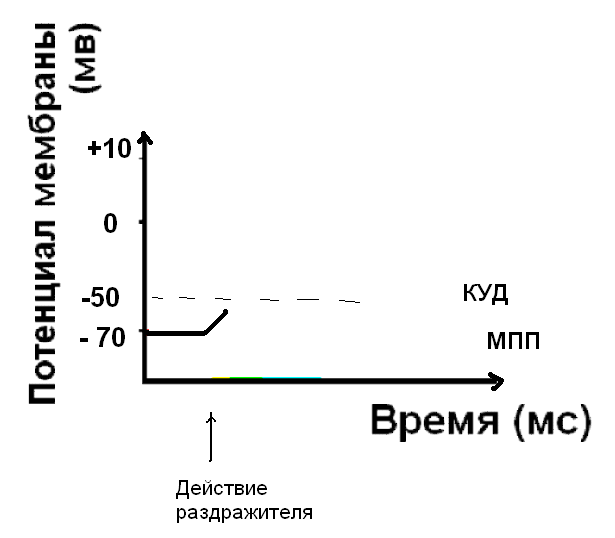
Если сила раздражителя достаточна, чтобы деполяризовать мембрану до КУД, открываются быстрые потенциал-зависимые натриевые каналы. Клетка возбуждена – возник нервный импульс.
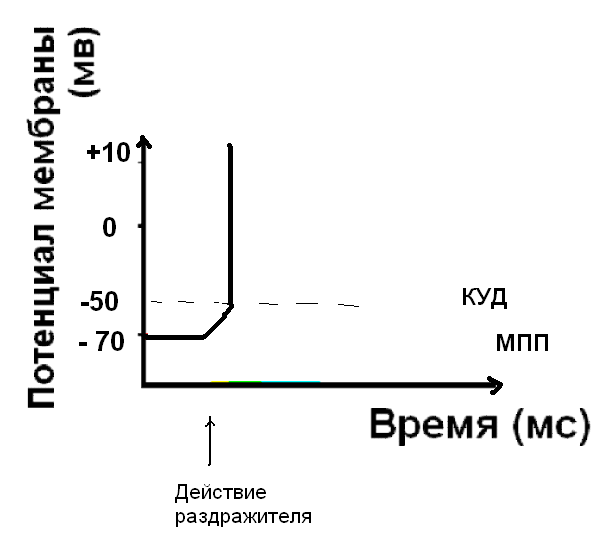
Восстановление мембранного потенциала покоя - реполяризация мембраны.
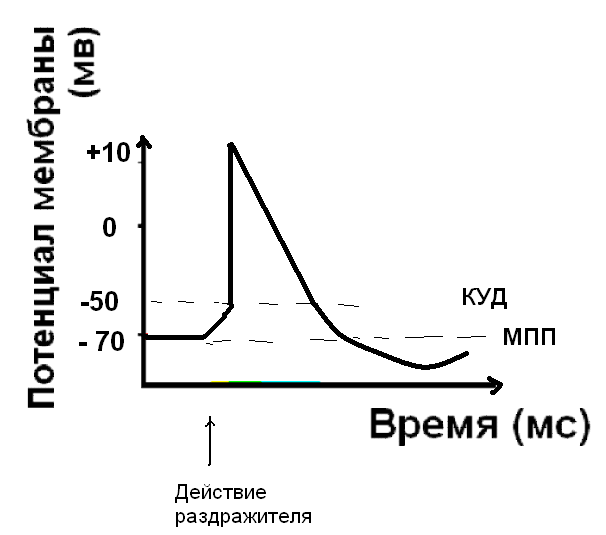
Следующий этап – восстановление мембранного потенциала покоя - реполяризация, обусловлена активным ионным транспортом. Наиболее важен процесс активного транспорта - это работа Na/K - насоса, который выкачивает ионы натрия из клетки, одновременно закачивая ионы калия внутрь клетки. Восстановление мембранного потенциала происходит благодаря току ионов калия из клетки – калиевые каналы активируются и пропускают ионы калия до достижения равновесного калиевого потенциала. Это процесс важен потому, что до тех пор, пока не восстановлен МПП, клетка не способна воспринимать новый импульс возбуждения.
ГИПЕРПОЛЯРИЗАЦИЯ - кратковременное увеличение МП после его восстановления, которое обусловлено повышением проницаемости мембраны для ионов калия и хлора. Гиперполяризация бывает только после ПД.
- Деполяризация мембраны до КУД – могут открыться любые натриевые каналы, иногда кальциевые, и быстрые, и медленные, и потенциал-зависимые, и рецептор-управляемые. Это зависит от вида раздражителя и типа клеток
- Быстрое поступление натрия в клетку - открываются быстрые, потенциал-зависимые натриевые каналы, и деполяризация достигает точки реверса потенциала – происходит перезарядка мембраны, знак заряда меняется на положительный.
- Восстановление градиента концентрации по калию – работа насоса. Калиевые каналы активированы, калий переходит из клетки во внеклеточную среду – реполяризация, начинается восстановление МПП
- Следовая деполяризация, или отрицательный следовой потенциал - мембрана еще деполяризована относительно МПП.
- Следовая гиперполяризация. Калиевые каналы остаются открытыми и дополнительный ток калия гиперполяризует мембрану. После этого клетка возвращается к исходному уровню МПП. Длительность ПД составляет для разных клеток от 1 до 3-4 мс.
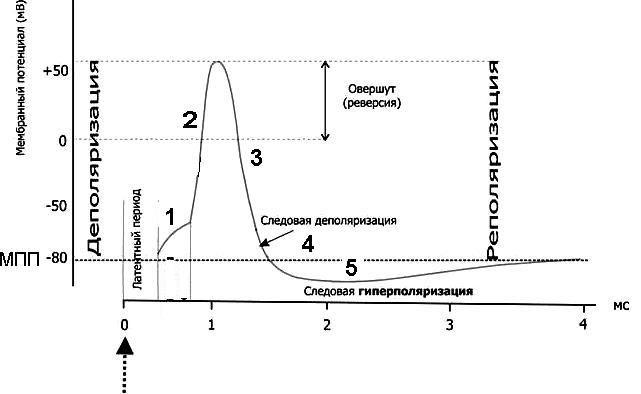
Обратите внимание на три величины потенциала, важные и постоянные для каждой клетки ее электрические характеристики.
- МПП - электроотрицательность мембраны клетки в покое, обеспечивающая способность к возбуждению - возбудимость.
- КУД - критический уровень, величина мембранного потенциала, при достижении которой открываются быстрые, потенциал зависимые натриевые каналы и происходит перезарядка мембраны за счет поступления в клетку положительных ионов натрия. Чем выше электроотрицательность мембраны, тем труднее деполяризовать ее до КУД, тем менее возбудима такая клетка.
- Точка реверса потенциала (овершут) - такая величина положительного мембранного потенциала, при которой положительно заряженные ионы уже не проникают в клетку - кратковременный равновесный натриевый потенциал.
При действии раздражителя подпороговой силы возникает неполная деполяризация - ЛОКАЛЬНЫЙ ОТВЕТ (ЛО). Неполная, или частичная деполяризация – это такое изменение заряда мембраны, которое не достигает критического уровня деполяризации (КУД).
Участие поджелудочной железы в пищеварении
Основную роль в переваривании пищи играет двенадцатиперстная кишка. Из поджелудочной железы в нее поступают пищеварительные ферменты, а из печени - желчь. Поджелудочный сок крайне важен для пищеварения. За сутки поджелудочной железой вырабатывается около 1,5-2 л густого сока. В нем, помимо пищеварительного сока, воды и бикарбоната, содержится множество пищеварительных ферментов, расщепляющих питательные вещества. Только в таком виде они могут всасываться через стенки кишки и с кровью или лимфой поступать в клетки. Основные питательные вещества, необходимые для организма человека - это белки, жиры и углеводы. Белки состоят из аминокислот и растворимы в воде. Углеводы - это т.н. полисахариды. Под действием ферментов эти полисахариды расщепляются на растворимые в воде моносахариды. Жиры состоят из глицерина и жирных кислот, они также должны быть расщеплены на составные части. Пищеварение начинается в полости рта и продолжается в желудке. В слюне содержатся ферменты, расщепляющие углеводы на моносахариды; в желудочном соке - соляная кислота и пепсин, расщепляющие белки. Пищевая масса, поступающая из желудка в двенадцатиперстную кишку, смешивается с поджелудочным соком, содержащим пищеварительные ферменты: одни из них расщепляют белки, другие - углеводы, третьи - жиры. Протеазы расщепляют белки, карбогидразы - углеводы, эстеразы - жиры. В поджелудочном соке имеются ферменты, расщепляющие нуклеиновые кислоты. Секреция поджелудочного сока зависит от пищи, напр., при употреблении хлеба его выделяется больше, при употреблении молочных продуктов - меньше. При нарушении выделения поджелудочного сока из поджелудочной железы неизбежно нарушается расщепление белков, углеводов, жиров и нуклеиновых кислот. В результате клетки организма человека получают недостаточно питательных веществ, и железы не способны нормально функционировать. Как правило, выделение поджелудочного сока нарушается при заболевании поджелудочной железы или при наличии опухоли.
СОК ПОДЖЕЛУДОЧНОЙ ЖЕЛЕЗЫ
За сутки вырабатывается 1,5—2,5литра сока. С момента начала пищеварения и в течение 4—6 часов происходит интенсивное выделение этого сока, в дальнейшем (если нет следующего приема) интенсивность секреции снижается. Количество сока и его состав зависят от вида пищи. Имеется четкая зависимость — меняется рацион, меняется состав сока.
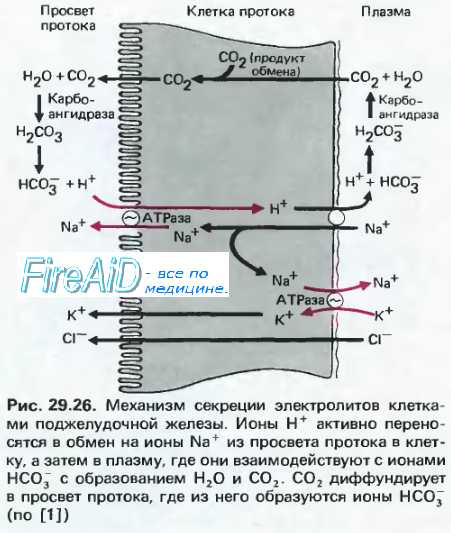
Сок имеет щелочную среду: рН = 7,5—8,8. Это обеспечивается огромным количеством" бикарбонатов в соке их концентрация в соке достигает 150ммоль/л (сравним в плазме крови — 24 ммоль/л). Панкреатический сок секретируется, главным образом, ацинозными панкреацитами.. Помимо бикарбонатов сок имеет набор всех гидролаз: амилаза, мальтаза, инверталаза, липаза, протеазы проэластаза. Протеазы (трипсиноген, химоТрипсиноген.) вырабатываются в неактивном виде. Попав в 12-перстную кишку, трипсиноген превращается под влиянием энтерокиназы в трипсин, и этот активированный фермент, вызывает активацию остальных протеаз панкреатического сока. Сок панкреатической железы выделяется в 12-перстную кишку через единый с общим желчным протоком сфинктер. В ряде случаев возможно попадение в панкреатическую железу сока из 12-перстной кишки, либо желчи или смеси их. В этом случае возможно внутрипанкреатическое активирование трипсиногена и остальных протеаз, что в конечном итоге вызывает развитие острого панкреатита
Регуляция выделения осуществляется за счет нервных и гуморальных влияний. Фазы: мозговая, желудочная и кишечная. Центры панкреатического сокоотделения расположены в тех же участках мозга, что и центры регуляции желудочного сокоотделения. Все влияния ЦНС осуществляются через вагус (он повышает секреторную активность) и симпатические волокна (торможение). Местно (в ЖКТ) вырабатываются стимуляторы панкреатического сокоотделения: секретин (усиливает в основном продукцию бикарбонатов), холецистокинин-панкреозимин (повышает продукцию ферментов), гастрин, серотонин, химоденин (повышает продукцию химотрипсиногена), желчные кислоты. Часть гормонов оказывает двойной эффект: вначале возбуждают, а потом — угнетают секрецию (глюкагон, соматостатин, калыштонин, ГИП, ПП, ВИП).
Назначение панкреатического сока — нейтрализация кислого содержимого в 12-перст- ной кишке (чем выше кислотность вышедшего из желудка химуса, тем выше продукция панкреатического сока и выше содержание в нем бикарбонатов) и гидролиз углеводов, жиров, белков, нуклеиновых кислот за счет полостного пищеварения.
Клетки панкреатической железы способны секретировать гормоны: инсулин (бета-клетки, или В-клетки), глюкагон (альфа-клетки, или А-клетки), соматостатин (дельта-клетки, или Д-клетки), панкреатический полипептид — ПП (РР-клетки). Здесь же, в панкреатической железе, секретируются серотонин, ВИП, гастрин, энкефалин, калликремн, а в клетках выводных протоков поджелудочной железы — липоксин (влияющий на жировой обмен) и ваготонин (его продукция повышает тонус вагуса).
пищи
—
трипсина и химотрипсина, а прием жирной
пищи вызывает секрецию сока с более
высокой его липолитической активностью.
Прием
одинаковой по характеру пищи длительное
время оказывает выраженное влияние на
деятельность поджелудочной железы.
Это влияние в основном состоит в
приспособлении количества и ферментного
состава сока к преобладающему
питательному веществу в рационе
питания человека и животного. Так,
преобладание белков повышает выделение
в составе панкреатического сока протеаз,
преимущественно углеводное питание
увеличивает выделение с соком амилазы,
а большое количество жира в рационе
снижает объем поджелудочной-секреции
и увеличивает содержание липазы в соке.
Секреция
поджелудочной железы регулируется
нервными и гуморальными механизмами.
И. П. Павлов в хронических и острых
опытах показал, что раздражение
блуждающего нерва обусловливает
выделение небольшого количества
поджелудочного сока, богатого ферментами.
Начальная
секреция поджелудочной железы вызывается
видом, запахом пищи и другими раздражителями
(условнорефлекторные
сигналы),
а также жеванием и глотанием
(безусловнорефлекторнЫе
сигналы).
При этом нервные сигналы, формирующиеся
в рецепторах полости рта и глотки,
достигают продолговатого мозга и затем
эфферентные влияния по волокнам
блуждающего нерва поступают к железе
и вызывают ее секрецию.
‘
У
человека, с фистулой поджелудочного
протока наблюдали.выделение
панкреатического сока через
2—3
мин после того, как ему говорили о пище,
которую дадут. Возбуждение
панкреатической секреции в данном
случае происходило условнорефлекторным
путём.
Симпатические
волокна, иннервирующие поджелудочную
железу, тормозят ее секреторную
активность. Поэтому после перерезки
чревных нервов у собак поджелудочная
секреция увеличивается. Симпатические
влияния, кроме того, имеют для железы
трофическое значение
—
они изменяют реактивность железы по
отношению к другим воздействиям,
усиливают синтез органических веществ
в ней.
Торможение
панкреатической секреции наблюдается
при раздражении многих центростремительных
нервов, при болевых реакциях, во время
сна, при напряженной физической и
умственной работе.
В
стимуляции панкреатической секреции
прямые нервные влияния имеют меньшее
значение, чем гуморальные.
Ведущее значение в гуморальной регуляции
секреции поджелудочной железы принадлежит
гастроинтестинальным гормонам
(рис.
187).
В
лаборатории И. П. Павлова было установлено,
что введение соляной кислоты в
двенадцатиперстную кишку вызывает
обильную секрецию поджелудочного сока.
В
1902
г. Бейлис и Старлинг показали, что
солянокислая вытяжка слизистой оболочки
двенадцатиперстной кишки, введенная
внутрйвенно, стимулирует поджелудочную
секрецию. Образующееся под влиянием
соляной кислоты в двенадцатиперстной
кишке вещество они назвали гормоном
секретином.
Секретин
вызывает выделение большого количества
поджелудочного сока, богатого
бикарбонатами, но бедного ферментами,
так как почти не действует на клетки
ацинусов, секретирующие ферменты.
Вторым
гормоном, усиливающим секрецию
поджелудочной железы, является
холецистокинин-панкреозимин.
Сначала считали, что это два разных
гормона. Один из них ‘(панкреозимин)
стимулирует секрецию поджелудочной
железы, а другой (холе-цистокин)
—
выход желчи в двенадцатиперстную кишку.
Теперь доказано, что это один гормон,
вызывающий несколько эффектов. В
наибольшей мере стимулируют высвобождение
этого гормона продукты начального
гидролиза пищевого белка и жира, а также

некоторые
аминокислоты. Стимулируют высвобождение
холе-цистокинина-панкреозимина соляная
кислота и углеводы.
Холецистокинин-панкрео-зимин
действует преимущественно на
панкреоциты ацинусов .поджелудочной
железы, поэтому выделяющийся в ответ
на действие этого гормона сок богат
ферментами. Одновременное влияние
на железу секретина и
холе-цистокинина-панкреозимина во время
приема пищи усиливает друг друга.
Рис.
187.
Регуляция секреции поджелудочной
железы.
I3
—
интерорецепторы;
G
—
гастринпродуцирующие клетки;
S
—секретинпродуцирующие
клетки:
1 —
холециетокинин-панкреози-мин (ХЦК-ПЗ)
-продуцирующие клетки;
Секреция
поджелудочной железы, усиливается’
также га-стрином,
серотонином, инсулином, бомбезином,
субстанцией П, солями желчных кислот.
Тормозят выделение поджелудочного
сока глюкагон,
кальцитонин, ЖИП, ПП, соматостатин. ВИП
может
возбуждать и тормозить панкреатическую
секрецию. Эффекты гормонов частично
опосредуются через их влияние на
желудочную секрецию:
с
усилением ее более кислое содержимое
поступает в двенадцатиперстную кишку
и посредством ее гормонов повышает
панкреатическую секрецию.
Нервные
влияния при приеме пищи обеспечивают
лишь пусковые
воздействия на железу,
а в коррекции панкреатической секреции
большую роль играют гуморальные
механизмы. Действие гормонов на железу
более выражено при сохраненной ее
иннервации, что подчеркивает единство
нервных и гуморальных механизмов
регуляции поджелудочной секреции.
Стимуляторы секреции поджелудочной
железы усиливают ее кровоснабжение,
что немаловажно для поддержания функции
железы на высоком уровне достаточно
длительное время.
Фазы
панкреатической секреции при стимуляции
ее приемом пищи те же, что и для желудочной
секреции, однако более выражены
гормональные влияния на поджелудочную
железу, особенно в кишечную фазу.
1. Всасывание. Функция всасывания кишечника. Транспорт питательных веществ. Щеточная кайма энтероцита. Гидролиз питательных веществ.
2. Всасывание макромолекул. Трансцитоз. Эндоцитоз. Экзоцитоз. Всасывание микромолекул энтероцитами. Всасывание витаминов.
3. Нервная регуляция секреции пищеварительных соков и моторики желудка и кишечника. Рефлекторная дуга центрального пищеводно — кишечного моторного рефлекса.
4. Гуморальная регуляция секреции пищеварительных соков и моторики желудка и кишечника. Гормональная регуляция пищеварительного тракта.
5. Схема механизмов регуляции функций желудочно-кишечного тракта (жкт). Обобщенная схема механизмов регуляции функций пищеварительного тракта.
6. Периодическая деятельность пищеварительной системы. Голодная периодическая деятельность пищеварительного тракта. Мигрирующий моторный комплекс.
7. Пищеварение в ротовой полости и функция глотания. Ротовая полость.
8. Слюна. Слюноотделение. Количество слюны. Состав слюны. Первичный секрет.
9. Отделение слюны. Секреция слюны. Регуляция выделения слюны. Регуляция секреции слюны. Центр слюноотделения.
10. Жевание. Акт жевания. Регуляция жевания. Центр жевания.
Гуморальная регуляция секреции пищеварительных соков и моторики желудка и кишечника. Гормональная регуляция пищеварительного тракта.
Центральные, периферические и местные рефлексы осуществляются в тесном взаимодействии с гуморальным механизмом регуляции миоцитов, гландулоцитов и нервных клеток.
В слизистой оболочке желудочно-кишечного тракта и в поджелудочной железе имеются эндокринные клетки, которые вырабатывают гастроинте-стинальные гормоны (регуляторные пептиды, энтерины). Эти гормоны через кровоток и местно (паракринно, диффундируя через межклеточную жидкость) оказывают влияние на миоциты, гландулоциты, интрамураль-ные нейроны и эндокринные клетки. Их выработка запускается рефлекторно (через блуждающий нерв) во время приема пищи и длительное время поддерживается за счет раздражающего влияния продуктов гидролиза пищевых веществ и экстрактивных веществ.
| Название гормона | Место выработки гормона | Типы эндокринных клеток | Эффект действия гормонов |
| Соматостатин | Желудок, проксимальный отдел тонкой кишки, поджелудочная железа | D-клетки | Тормозит выделение инсулина и глюкагона, большинства известных желудочно-кишечных гормонов (секретина, ГИПа, мотилина, гастрина); тормозит активность париетальных клеток желудка и ацинарных клеток поджелудочной железы |
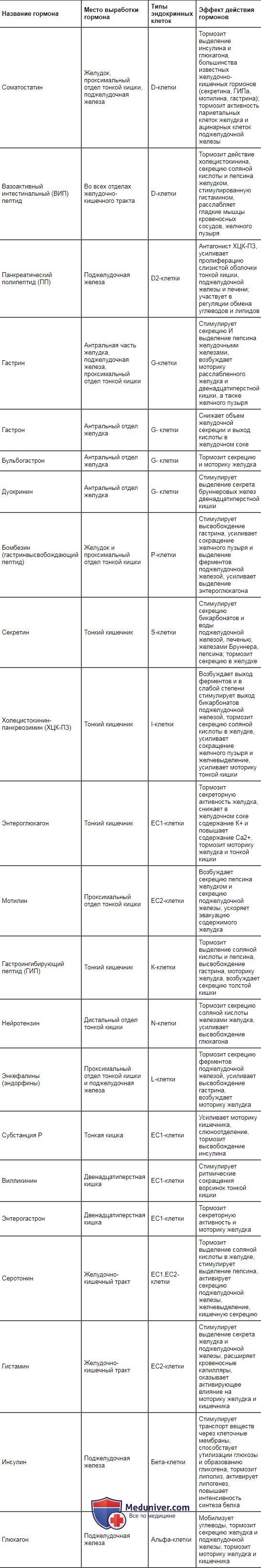
Место выработки основных гастроинтестинальных гормонов, вызываемые ими эффекты и продуцирующие их клетки представлены в табл. 11.1. В настоящее время обнаружено около 30 регуляторных пептидов. Как следует из представленной таблицы, они оказывают стимулирующее, тормозное и модулирующее влияние на секрецию пищеварительных соков, моторику гладкой мускулатуры желудочно-кишечного тракта, всасывание, выделение энтеринов эндокринными элементами слизистой оболочки желудка, кишечника и поджелудочной железы.
Выделение гастроинтестинальных гормонов имеет каскадный характер. Например, под влиянием гастрина обкладочные клетки желез желудка увеличивают выработку соляной кислоты, которая в слизистой оболочке тонкой кишки стимулирует выделение S- и J-клетками секретина и холеци-стокинна — панкреозимина. Секретин усиливает секрецию воды и бикарбонатов поджелудочной железой и печенью, а холецистокинин — панкреозимин — возбуждает выделение ферментов поджелудочной железой и тормозит секрецию соляной кислоты обкладочными клетками, усиливает моторику тонкой кишки и желчного пузыря.
Регуляторные пептиды, поступая в кровоток, быстро разрушаются в печени и почках и тем самым создают условия для осуществления эффектов других гастроинтестинальных гормонов.
1. Регуляция сократительной деятельности желудка. Сокращение желудка.
2. Эвакуация содержимого желудка в двенадцатиперстную кишку. Эвакуация пищи (пищевого комка ) в двенадцатиперстную кишку. Энтерогастральный рефлекс.
3. Пищеварение в двенадцатиперстной кишке. Пищеварительные функции поджелудочной железы.
4. Состав панкреатического сока. Свойства панкреатического сока. Ферменты поджелудочной железы.
5. Нервная регуляция секреторной функции поджелудочной железы. Гуморальная ( гормональная ) регуляция секреции поджелудочной железы.
6. Секреция сока поджелудочной железы. Фазы ( этапы ) секреции сока поджелудочной железы.
7. Пищеварительная функция печени. Желчь. Механизм образования желчи. Образование желчи.
8. Состав желчи. Свойства желчи. Печеночная желчь. Пузырная желчь.
9. Регуляция желчеобразования. Регуляция желчевыведения.
10. Непищеварительные функции печени. Функции печени.
Нервная регуляция секреторной функции поджелудочной железы. Гуморальная ( гормональная ) регуляция секреции поджелудочной железы.
Слабое сокоотделение натощак резко усиливается во время (через 2— 3 мин) и после приема пищи. В привычных условиях приема пищи панкреатическое сокоотделение начинается уже на вид, запах пищи и другие раздражители, сопутствующие приему пищи, что свидетельствует об условно-рефлекторном механизме секреции. При раздражении пищевыми веществами рецепторов слизистой оболочки ротовой полости, глотки и пищевода возникает безусловно-рефлекторное отделение панкреатического сока. Нервные импульсы от раздражаемых рецепторов по афферентным путям достигают бульбарного центра панкреатической секреции, где они переключаются на преганглионарные нейроны ядер блуждающего нерва, по эфферентным волокнам которого достигают постганглионарных нейронов. Их аксоны образуют синаптические окончания на базальных мембранах панкреацитов. Выделяющийся при возбуждении этих окончаний ацетилхо-лин активирует М-холинорецепторы постсинаптической мембраны. При этом освобождаются вторичные посредники (Са++ и ГЦ-цГМФ), которые и вызывают секреторную деятельность панкреоцитов и эпителиальных клеток протоков.
При раздражении чревных нервов, осуществляющих симпатическую иннервацию поджелудочной железы, ее сокоотделение тормозится (за счет активации р-адренорецепторов медиатором норадреналином). Но торможение секреции сопровождается накоплением секреторного материала в панкреацитах.
Поступление пищи в желудок во время приема пищи вызывает раздражение хемо- и механорецепторов желудка, что приводит к продолжению безусловно-рефлекторного сокоотделения поджелудочной железой, вызванного стимуляцией рецепторов ротовой полости.
Поступление в двенадцатиперстную кишку желудочного содержимого вызывает наряду с дуоденопанкреатическим секреторным рефлексом (с рецепторов слизистой оболочки) выделение из эндокринных клеток гастро-интестинальных гормонов, стимулирующих или тормозящих сокоотделение поджелудочной железой. Роль основных гуморальных регуляторов выполняют секретин и ХЦК, которые вырабатываются S- и ССК-клетками слизистой оболочки двенадцатиперстной кишки. Секретин и ХЦК усиливают влияние друг друга на панкреоциты, особенно на фоне выделения ацетилхолина в синаптических окончаниях холинергических нервных волокон, иннервирующих железу. Важным гуморальным возбудителем панкреатической секреции является гастрин, выделяемый У-клетками слизистой оболочки антрального отдела желудка. Возбуждающее влияние этих гормонов подкрепляется бомбезином, серотонином и инсулином.
Тормозят выделение сока соматостатин, глюкагон, энкефалины, вещество Р, ТИП, ПП, а также кальцитонн и АКТГ. Но их роль в комплексной гуморальной регуляции поджелудочной железы изучена недостаточно.

Поджелудочный сок выделяется только во время пищеварения. Когда пищеварение кончается, прекращается и выделение сока. Поджелудочный сок начинает выделяться через 2— 3 минуты после начала приема пищи.
Количество и свойство поджелудочного сока меняются в зависимости от пищи. На разные пищевые вещества поджелудочный сок, как и желудочный сок, выделяется в разных
количествах и с разным содержанием ферментов.
Рис. ОТДЕЛЕНИЕ ПОДЖЕЛУДОЧНОГО СОКА У СОБАКИ ПРИ КОРМЛЕНИИ МЯСОМ, ХЛЕБОМ И МОЛОКОМ.
Кривые выделения поджелудочного сока на мясо, хлеб и молоко, приведенные на рис , почти с точностью совпадают с кривыми выделения желудочного сока на эти же вещества.
Рефлекторное выделение поджелудочного сока
Рефлекторный механизм выделения сока поджелудочной железы был доказан И. П. Павловым в опытах на собаках, в частности в опыте с эзофаготомированной собакой. У такой собаки, помимо перерезки пищевода и желудочной фистулы, была наложена фистула поджелудочной железы. У оперированной таким образом собаки при мнимом кормлении через 2—3 минуты начинается выделение поджелудочного сока. Такой короткий промежуток между раздражением рецепторов полости рта и началом выделения сока говорит о том, что здесь имеет место рефлекторный механизм.
К. М. Быков и его сотрудники проводили наблюдение над человеком с фистулой поджелудочной железы. Выделение поджелудочного сока у него начиналось через 2—3 минуты после разговора о еде. В данном случае имело место условно-рефлекторное сокоотделение.
В эксперименте с предварительной перерезкой блуждающего нерва и последующим перерождением его волокон, тормозящих секрецию поджелудочной железы, И. П. Павлов доказал, что секреторным нервом поджелудочной железы является именно блуждающий нерв, раздражение которого вызывает отделение небольшого количества сока. При еде пища раздражает окончания центростремительных нервов, находящиеся в слизистой оболочке рта и глотки. Возникшее в них возбуждение передается в центр поджелудочного сокоотделения, который находится в продолговатом мозгу, и оттуда по блуж дающему нерву достигает поджелудочной железы; последняя начинает выделять сок.
Нервно-гуморальный механизм выделения поджелудочного сока
При введении в двенадцатиперстную кишку желудочного сока, который содержит соляную кислоту, начинается выделение поджелудочного сока. Поджелудочная железа начинает секретировать и при введении в кишку одной соляной кислоты.
Было высказано предположение, что соляная кислота раздражает нервные окончания и рефлекторно вызывает сокоотделение. Это предположение не оправдалось, так как поджелудочная железа продолжает секретировать и после того, как все нервы, идущие к ней, перерезаны.
Однако соляная кислота вызывает сокоотделение только в том случае, когда ее вводят в двенадцатиперстную кишку. Если соляную кислоту ввести в кровь, минуя кишку, то сокоотделения не наступает; следовательно, не сама соляная кислота вызывает секрецию поджелудочной железы.
Механизм действия соляной кислоты при введении ее В двенадцатиперстную кишку был исследован в специальных опытах. Он состоит в том, что в слизистой оболочке кишки под влиянием соляной кислоты образуется особое вещество — с е к р е т и н. Секретин, всасываясь и поступая в ток крови, кровью доставляется к поджелудочной железе, действует на ее нервно-железистый аппарат и вызывает сокоотделение. Окончательным доказательством явился следующий опыт: у собаки вырезали двенадцатиперстную кишку, положили ее в 0 ,5% раствор соляной кислоты, а через некоторое время этот раствор ввели в кровь другой собаке; тогда поджелудочная
железа второй собаки начала секретировать. В естественных условиях пища, обильно смоченная кислым желудочным соком, поступает в двенадцатиперстную кишку. Соляная кислота желудочного сока действует на слизистую оболочку и вызывает образование в ней из недеятельного просекретина секретина, который затем разносится кровью и возбуждает поджелудочную железу.
Данные, полученные в последнее время, показывают, что секретин действует при участии нервной системы. Это было установлено в опытах, в которых симпатическая нервная система выключалась эрготоксином. Если такому животному в двенадцатиперстную кишку ввести соляную кислоту, то сок из поджелудочной железы почти не будет выделяться. Отсюда следует, что, вероятно, секретин действует на секреторные клетки поджелудочной железы через нервную систему.
Жир, продукты расцеплении которого —мыла — обладают сильным сокогонным действием, также вызывает выделение поджелудочного сока; кроме того, поступление жира в кишечник способствует усиленному образованию секретина в слизистой оболочке кишечника.
Выделение поджелудочного сока усиливает вода, особенно газированная, клюквенный морс и т. д.
Деятельность поджелудочной железы нарушается у людей, желудочный сок которых лишен соляной кислоты или беден ею. Пониженное содержание соляной кислоты в желудочном соке или ее отсутствие называется ахилией. Ввиду того что у таких людей соляная кислота в двенадцатиперствую кишку не поступает, то не происходит и образования секретина и деятельность поджелудочной железы нарушается. Это нарушение отчасти восполняется тем, что вода и некоторые другие вещества способствуют деятельности поджелудочной железы и обеспечивают ее функционирование.
Статья на тему Выделение поджелудочного сока
Читайте также: