
Мякотные нервные волокна седалищный нерв лягушки
Препарат № 116. Мякотное нервное волокно (не фиксированное). Седалищный нерв лягушки (рис. 120)
Для изучения строения мякотного нервного волокна необходимо рассмотреть препараты № 116 и 117. Первый препарат дает общее представление о нервном волокне - о соотношении мякотной оболочки и аксона, о структуре перехватов Ранвье и насечек Лантермана (последние особенно хорошо видны на неокрашенном нерве).
Второй препарат (№ 117) следует рассмотреть специально с целью подробного исследования мякотной оболочки. Только при изучении данного препарата становится ясным, что оболочка нерва имеет синцитиальное строение.
Убивают лягушку, обезглавливая ее или разрушая спинной мозг, и прикрепляют препаровальными иглами к восковой ванночке, спинной стороной кверху. Разрезают кожу бедра, отодвигают ее и обнажают мышцы. Если осторожно раздвинуть мышцы, то становится виден седалищный нерв. Этот нерв имеет сравнительно большую толщину и белую окраску, благодаря чему хорошо заметен.
Для того чтобы изолировать седалищный нерв, удаляют загораживающие его мышцы, приподнимают нерв пинцетом и подрезают его в местах прикрепления. Затем разрезают нерв на куски длиной около 1 см и помещают в физиологический раствор. На предметном стекле в капле физиологического раствора нерв
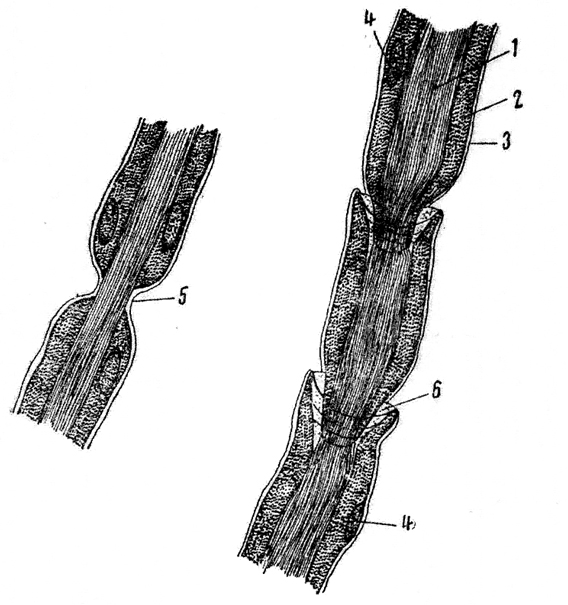
Рис. 120. Мякотное нервное волокно из седалищного нерва лягушки (увеличение - ок. 10, об. 40):
1 - аксон, 2-мякотяая оболочка, 3 - иейрилемма, 4 - шванновское ядро, 5 - перехват Ранвье, 6 -насечка Лантермава
тщательно расщипывают иголками в продольном направлении, покрывают покровным стеклом и рассматривают под микроскопом.
На препарате будет заметна группа нервных волокон разной толщины (отростки клеток). Следует наметить для наблюдения волокно, лежащее свободно и несколько изолированно от других волокон. При большом увеличении микроскопа сразу видно, что нервное волокно имеет центральную часть - осевой цилиндр, или аксон, и оболочку. Применяя иммерсионную систему, в осевом
цилиндре можно заметить слабую продольную исчерченность - это нейрофибриллы, идущие из тела нервной клетки и проходящие параллельно друг другу в осевом цилиндре. Оболочка, окружающая нерв, так называемая мякотная оболочка, содержит сильно преломляющее свет вещество-миелин. Миелин одевает осевой цилиндр не на всем его протяжёнии. В некоторых местах волокно сужается и образуются перетяжки, лишенные миелина, так называемые перехваты Ранвье. Расстояния между перехватами в разных нейритах различны. Следует внимательно рассмотреть перехват Ранвье. В нем не только прерывается миелиновая оболочка, но иногда сужается и сам осевой цилиндр. Кроме того, можно заметить, что в местах перехвата проходит непрерывная, очень тонкая, совершенно прозрачная оболочка - нейрилемма, которая покрывает с поверхности все нервное волокно; она плохо заметна вследствие незначительной толщины. Таким образом, нейрилемма вместе с миелином составляет мякотную оболочку нервного волокна.
Иногда в мякотной оболочке под нейрилеммой заметны шванновские ядра удлиненной формы.
Иммерсионная система дает возможность рассмотреть в нервном волокне особые образования - насечки Лантермана. В некоторых местах с обеих сторон миелиновой оболочки имеются светлые полоски, как бы надрезы, расположенные косо и направленные в ту или иную сторону. Вокруг этих надрезов иногда бывают заметны циркулярно расположенные тонкие волоконца. Каждая насечка, по-видимому, представляет собой протоплазматическую воронку, которая залегает в мякотной оболочке нерва.
Кроме мякотных волокон, в состав седалищного нерва входят и безмякотные нервные волокна (см. препарат № 118). Они отличаются тем, что значительно тоньше мякотных и не имеют, миелинозой оболочки.
При подсыхании препарата нерв претерпевает посмертные изменения. Мякотная оболочка становится неровной, миелин собирается в капли и часто вытекает из волокна.
В связи с тем, что не всегда имеется возможность получить свежий препарат нервного волокна, мы рекомендуем для изучения также и осмированный препарат.
Вырезают кусочек седалищного нерва лягушки способом, описанным выше. Осторожно расщипывают его иглами в продольном направлении и помещают на 20-30 мин в 1%-ный раствор осмиевой кислоты. Затем тщательно промывают препарат в дистиллированной воде в течение 30-40 мин. и помещают для окрашивания в пикрокармин или 2%-ный кислый фуксин на время от нескольких часов до суток. Обезвоживая нерв для заключения в бальзам, нужно еще раз дополнительно расщипать его в 96° спирте и в ксилоле. Можно приготовлять
непостоянные препараты, помещая нерв после окрашивания и промывания в глицерин.
Изучаемый препарат аналогичен предыдущему. Следует отметать только то, что мякотная оболочка здесь черная, так как миелин окрашивается осмием в черный цвет, шванновские ядра красные, а лантермановские насечки имеют вид светлых, косо расположенных полосок.
Физиологические свойства нервных волокон. Мякотные и безмякотные нервные волокна
Нерв, или нервный ствол, представляет собой сложное образование, состоящее из большого количества нервных волокон, заключенных в общую соединительнотканную оболочку. Нервные волокна - это отростки нервных клеток. В состав нерва входят длинные отростки нервных клеток (аксоны), которые несут возбуждение в центральную нервную систему или от нее на периферию. Нервы, как правило, являются смешанными, в их состав входят двигательные и чувствительные нервные волокна.
Нервные волокна, как и все возбудимые структуры, обладают следующими физиологическими свойствами: возбудимостью, проводимостью, рефрактерностью, лабильностью.
При сравнении основных физиологических свойств нервной и мышечной ткани обращает на себя внимание то обстоятельство, что возбудимость и лабильность нервного волокна выше, а рефрактерный период короче, чем у мышечной ткани. Данная особенность связана с более высоким уровнем обменных процессов в нерве. Например, абсолютный рефрактерный период мякотного нерва продолжается 0,002 с, а скелетной мышцы - 0,005 с. Лабильность двигательного нерва нервно-мышечного препарата лягушки составляет 500-1000 имп/с, лабильность же мышцы - 200-250 имп/с.
Проведение возбуждения является специализированной функцией нервных волокон. Скорость проведения возбуждения по нервным волокнам в основном зависит от диаметра и гистологических особенностей строения нервных волокон. Чем больше диаметр нервного волокна, тем скорость распространения возбуждения в нем выше. Так, скорость распространения возбуждения по нервному волокну с диаметром 12-22 мкм составляет 70-120 м/с, а по нервному волокну с диаметром 8-12 мкм - 40-70 м/с.
В зависимости от гистологических особенностей строения нервные волокна делятся на мякотные (миелиновые) и безмякотные (безмиелиновые).
Миелинизированное волокно (рис. 57) состоит из осевого цилиндра и покрывающих его миелиновой и шванновской оболочек. Миелиновая оболочка состоит из жироподобных веществ, обладает высоким удельным сопротивлением и выполняет в связи с этим роль своеобразного изолятора. Миелиновая оболочка через промежутки равной длины прерывается, оставляя открытыми участки осевого цилиндра шириной около 1 мкм. Эти участки получили название перехватов Ранвье. Длина межперехватных участков зависит от диаметра волокна и колеблется в пределах от 0,2·10 -3 м (0,2 мм) до 1·10 -3 -2·10 -3 м (1-2 мм). Поверхность осевого цилиндра представлена плазматической мембраной, а его содержимое - аксоплазмой.
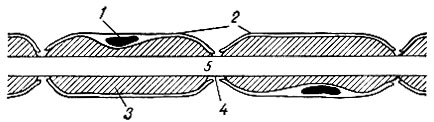
Рис. 57. Строение миелинизированного нервного волокна (схема). 1 - ядро шванновской плетки; 2 - шванновская оболочка; 3 - миелин; 4 - перехват Ранвье; 5 - аксоплазма
Безмякотные нервные волокна не имеют миелиновой оболочки, они покрыты только шванновскими клетками (леммоциты). Между шванновскими клетками и осевым цилиндром имеется щель 15 нм (150 А), которая заполнена межклеточной жидкостью. В связи с указанной особенностью строения поверхностная мембрана осевого цилиндра сообщается с окружающей нервное волокно средой (межклеточная жидкость).
Основоположником учения о проведении возбуждения по нервным волокнам следует считать немецкого физиолога Германа (1885). Он полагал, что возбуждение по нервным волокнам распространяется за счет малых круговых токов, которые возникают внутри волокна и в окружающей его жидкости (рис. 58).
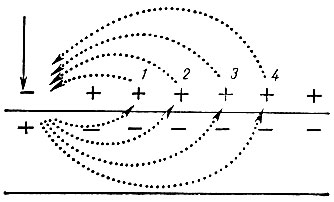
Рис. 58. Схема распространения возбуждения по нервному волокну за счет малых круговых токов. Вертикальной стрелкой обозначено место нанесения раздражения. Круговые стрелки показывают направление движения электрического тока внутри волокна и в окружающей жидкости
В настоящее время теория круговых токов Германа получила теоретическую разработку и экспериментальное подтверждение в работах Ходжкина и других исследователей.
Между возбужденным и невозбужденным участками нервного волокна в аксоплазме и в окружающей жидкости ток будет распространяться от плюса к минусу. Это приведет к возникновению так называемых малых, или круговых, токов, которые, выходя из нервного волокна, последовательно возбуждают его участки (1, 2 и т. д.). По мере удаления от очага возбуждения (участки 3 и 4) раздражающее действие круговых токов ослабевает и они становятся неспособными вызвать возбуждение. Таким образом, в случае последовательного распространения возбуждения по каждому участку нервного волокна нервные импульсы передаются с затуханием за счет непрерывно бегущей волны.
Из-за гистологических особенностей строения мякотных нервных волокон, в частности наличия миелиновой оболочки, обладающей высоким сопротивлением, электрические токи могут входить в волокна указанного типа и выходить из них только в области перехватов Ранвье (рис. 59).

Рис. 59. Скачкообразное распространение возбуждения в мякотном нервном волокне от перехвата к перехвату. Стрелками показано направление тока, возникающего между возбужденным (А) и соседним покоящимся (Б) перехватом
При нанесении раздражения возникает деполяризация в области ближайшего перехвата Ранвье - А. Соседний перехват Ранвье - Б находится в состоянии поляризации. Между перехватами возникает разность потенциалов, которая приводит к появлению круговых токов (см. рис. 58). Ионный поток в аксоплазме и в окружающей среде течет от плюса к минусу. Выход круговых токов в перехвате Б приводит к его деполяризации и возникновению потенциала действия. Далее за счет круговых токов возбуждаются последующие перехваты Ранвье. Таким образом, возбуждение в мякотных нервных волокнах передается скачкообразно (сальтаторно) от одного перехвата Ранвье к другому, Сальтаторный способ передачи возбуждения более экономичен, нежели распространение возбуждения по безмякотным нервным волокнам. Возбуждение по мякотным нервным волокнам распространяется без затухания. Скорость распространения возбуждения по мякотным нервным волокнам гораздо выше, чем по безмякотным. Так, скорость распространения возбуждения по двигательным нервным волокнам (мякотные нервы) составляет 80-120 м/с, по волокнам, не покрытым миелиновой оболочкой, - от 0,5 до 2 м/с.
При нанесении раздражения на нервное волокно происходит двустороннее распространение возбуждения - в центростремительном и центробежном направлениях. Это доказывается следующим опытом. К нервному волокну прикладывают две пары электродов - А и Б, связанных с электроизмерительными приборами (рис. 60). Раздражение наносят между электродами А и Б с помощью раздражающих электродов С. В результате двустороннего проведения возбуждения вдоль клеточной мембраны приборы зарегистрируют прохождение нервных импульсов как под электродом А, так и под электродом Б.
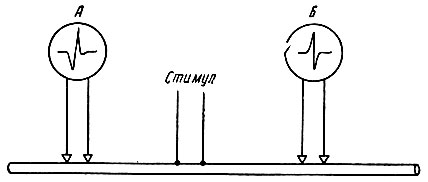
Рис. 60. Схема опыта для доказательства двустороннего проведения возбуждения по нерву. Объяснение в тексте
Двустороннее проведение возбуждения по нервному волокну впервые описано русскими учеными Р. И. Бабухиным (1887) и Кюне (1886). Оно не противоречит одностороннему распространению нервных импульсов в целостном организме, которое объясняется местом возникновения нервных импульсов (рецептор или нервный центр), а также наличием специальных образований - синапсов.
Возбуждение проводится только по одному нервному волокну, не распространяясь на соседние волокна, что обусловливает осуществление строго координированной рефлекторной деятельности. Периферический нервный ствол обычно состоит из большого количества нервных волокон. Так, в состав седалищного нерва входят тысячи нервных волокон: мякотные и безмякотные, афферентные и эфферентные, соматические и вегетативные. В случае неизолированного проведения возбуждения наблюдалась бы хаотическая ответная реакция.
Изолированное проведение возбуждения в мякотных нервных волокнах обеспечивается миелиновой оболочкой, в безмякотных - высоким удельным сопротивлением окружающей нервное волокно жидкости. Доказать наличие изолированного проведения возбуждения можно в эксперименте на препарате задней лапки лягушки, раздражая отдельные корешки седалищного нерва. Более строгое доказательство может быть получено при отведении потенциала действия от отдельных нервных волокон, входящих в состав нервного ствола.
Химические изменения в нервных волокнах при возбуждении. Неспецифическим признаком возбуждения, как уже отмечалось, является повышение обменных процессов. Возбуждение нерва сопровождается увеличением распада богатых энергией фосфорных соединений АТФ (аденозинтрифосфорная кислота) и креатинфосфата, а также повышенным образованием молочной кислоты. Кроме того, наблюдается усиление процессов распада и синтеза углеводов, белков и липидов.
Стимуляция обменных процессов приводит к увеличению потребления кислорода и выделению соответствующих количеств углекислого газа. Так, нерв лягушки в состоянии покоя потребляет 1·10 -3 м 3 (1 мм 3 ) кислорода на 1·10 -3 кг/мин (1 г/мин), при возбуждении потребление кислорода возрастает на 10-20%, то же происходит и с продукцией углекислого газа.
Обмену веществ и его усилению при возбуждении соответствуют и термические соотношения. По данным Хилла, 1·10 -3 кг (1 г) вещества нерва в состоянии покоя образует в 1 с 2·10 -5 кал, а при возбуждении - 6,9·10 -5 кал. Таким образом, по теплопродукции нерва можно судить о динамике обмена веществ в нем.
Утомление нервного волокна. Н. Е. Введенский в 1883 г. впервые установил, что нерв мало утомляем. Он проводил опыт на нервно-мышечном препарате лягушки, раздражая седалищный нерв индукционным током в течение 6-8 ч. О возбуждении нерва Н. Е. Введенский судил по сокращению мышцы. Для того чтобы исключить утомление мышцы, распространение нервных импульсов к ней прерывалось действием катода постоянного тока. При выключении постоянного тока возбуждение вновь достигало мышцы и она сокращалась. В этих условиях установлено, что нерв длительно (на протяжении многих часов) сохранял способность возбуждаться и проводить волны возбуждения. Данные экспериментов Н. Е. Введенского подтверждены современными исследованиями с регистрацией биотоков нерва.
Малая утомляемость нервных волокон объясняется тем, что энергетические затраты в них при возбуждении незначительны, а восстановительные процессы протекают быстро.
В целостном организме малой утомляемости нервных волокон способствует также их работа с постоянной недогрузкой. Так, двигательное нервное волокно обладает высокой лабильностью (может воспроизводить до 2500 имп/с, из нервных же центров на периферию обычно проводится прерывисто не более 50-100 волн возбуждения в 1 с).
Таким образом, практическая неутомляемость нервных волокон связана с низкими энергетическими затратами при возбуждении, с высокой лабильностью нервных волокон и постоянной работой их с недогрузкой.
Синапс - функциональный контакт между возбудимыми клетками, цитоплазма каждой из которых заключена в отдельную электрогенную мембрану.
В зависимости от локализации синапсы делят на центральные и периферические.
Центральные синапсы осуществляют контакт между нервными клетками центральной нервной системы.
Периферические синапсы могут быть мионевральными и невроэпителиальными. За счет невроэпителиальных синапсов осуществляется нервная регуляция деятельности железистого аппарата. Мионевральные синапсы представляют собой функциональную связь между аксоном мотонейрона и мышечными волокнами. На основании результатов электронно-микроскопических исследований в мионевральном синапсе различают три основные структуры: пресинаптическую мембрану, постсинаптическую мембрану и синаптическую щель (рис. 61).
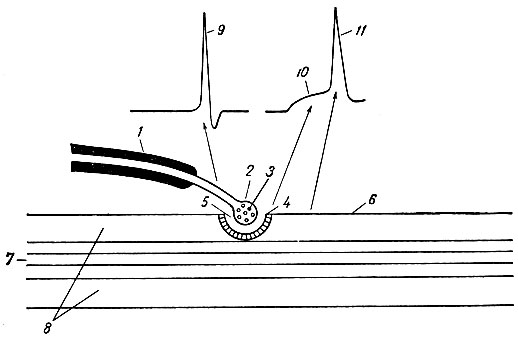
Рис. 61. Взаимоотношение между нервным волокном, нервным окончанием и скелетным мышечным волокном (схема). 1 - миелинизированное нервное волокно; 2 - нервное окончание с пузырьками медиатора; 3 - пресинаптическая мембрана; 4 - постсинаптическая мембрана мышечного волокна; 5 - синаптическая щель; 6 - внесинаптическая мембрана мышечного волокна; 7 - миофибриллы; 8 - саркоплазма; 9 - потенциал действия нервного волокна; 10 - потенциал концевой пластинки (постсинаптический потенциал); 11 - потенциал действия мышечного волокна
Пресинаптическая мембрана представляет собой электрогенную мембрану нервного окончания, аксоплазма которого включает большое количество гранул, или пузырьков, содержащих ацетилхолин.
Постсинаптическая мембрана, или концевая пластинка, - это электрогенная мембрана мышечного волокна, имеющая большое количество складок, что приводит к увеличению ее площади. Постсинаптическая мембрана содержит холинорецепторы, представляющие собой специальные белки мембраны. Кроме того, на постсинаптической мембране обнаружен фермент холинэстераза, который разрушает ацетилхолин,
Пресинаптическая и постсинаптическая мембраны разделяются узкой синаптической щелью - 20-50 нм (200-500 А), что обеспечивает быструю диффузию ацетилхолина. Синаптическая щель открывается во внеклеточное пространство и заполнена межклеточной жидкостью.
Передача возбуждения через мионевральный синапс осуществляется в результате выделения нервными окончаниями высокоактивных химических соединений, называемых медиаторами (посредниками, передатчиками нервного импульса). Предположение о химической передаче возбуждения через мионевральный синапс было высказано в 1924 г. русским физиологом А. Ф. Самойловым. Впоследствии было показано, что этим веществом является ацетилхолин.
Проведение возбуждения через мионевральный синапс осуществляется следующим образом. Кратковременная деполяризация пресинаптической мембраны, вызванная потенциалом действия нервного волокна, обеспечивает освобождение ацетилхолина из пузырьков терминали. Освобожденный ацетилхолин проникает через синаптическую щель к постсинаптической мембране и связывается с холинорецепторами. Указанное взаимодействие сопровождается повышением проницаемости постсинаптической мембраны для ионов натрия и калия, что обусловливает деполяризацию, которая проявляется в виде электроотрицательного возбуждающего постсинаптического потенциала (ВПСП). ВПСП имеет характер нераспространяющейся волны возбуждения, или локального ответа.
По мере развития ВПСП между деполяризованной постсинаптической мембраной и недеполяризованной мембраной мышечного волокна, примыкающей к ней, возникает разность потенциалов и появляются местные или круговые токи. При достижении ими порогового уровня в мембране мышечного волокна возникает распространяющийся мышечный потенциал действия.
Следует отметить, что связь ацетилхолина с холинорецептором непрочная. Молекулы ацетилхолина могут вступать в контакт с холинэстеразой, что приводит к распаду медиатора. В результате этого восстанавливается готовность синапса к проведению следующего нервного импульса.
Сильный и частый раздражитель приводит к выделению избыточных количеств ацетилхолина на пресинаптической мембране. Проникающий на постсинаптическую мембрану медиатор не успевает разрушаться холинэстеразой, что является причиной суммации постсинаптических потенциалов, возникновения стойкой деполяризации постсинаптической мембраны и блока проведения возбуждения.
Таким образом, в основе передачи возбуждения через мионевральный синапс лежат потенциал действия и сложные взаимодействия ацетилхолина с постсинаптической мембраной.
Синапсы обладают следующими основными физиологическими свойствами: односторонностью проведения возбуждения (в отличие от двустороннего проведения возбуждения в нервном волокне) и синаптической задержкой.
Одностороннее проведение возбуждения в синапсе связано с морфологическими его особенностями. Ацетилхолин вырабатывается в пресинаптической мембране, диффундирует через синаптическую щель и взаимодействует с холинорецептором, локализованным на постсинаптической мембране. Кроме того, синаптическая щель препятствует электрическому распространению возбуждения с нервного волокна на мышцу и с мышцы на нервное волокно.
Синаптическая задержка обусловливается следующими факторами: временем, необходимым для освобождения и диффузии медиатора через синаптическую щель, временем взаимодействия ацетилхолина с холинорецептором. В мионевральном синапсе задержка равна примерно 0,2-0,5 мс.
Н. Е. Введенский показал, что ответная реакция мышцы при увеличении силы или частоты раздражителя не может беспредельно возрастать.
На классическом объекте - нервно-мышечном препарате лягушки - было установлено, что при нарастании частоты раздражителя от 10 до 50 имп/с наблюдается увеличение амплитуды мышечного сокращения. Максимальная ответная реакция обнаруживалась при частоте 40-50 имп/с. Дальнейшее увеличение частоты раздражителя приводило к снижению амплитуды мышечного сокращения или к отсутствию механической реакции мышцы. Уменьшение частоты раздражителя сопровождалось восстановлением амплитуды мышечного сокращения.
Таким образом, Н. Е. Введенским было открыто и изучено совершенно новое явление в физиологии, которое свидетельствовало о том, что отсутствие ответной реакции мышцы на раздражение не связано с наступлением утомления в ней. Аналогичная закономерность в сократительном эффекте мышцы была установлена H. Е. Введенским и при увеличении силы раздражения, наносимого на двигательный нерв нервно-мышечного препарата лягушки.
Изменение реакции мышцы в зависимости от силы и частоты раздражителя позволило Н. Е. Введенскому дать определение оптимума и пессимума.
Оптимум (наилучший) - такая сила и частота раздражителя, которая обусловливает максимальную амплитуду мышечного сокращения.
Пессимум (наихудший) - такая чрезмерная (большая) сила и частота раздражителя, которая вызывает резкое уменьшение амплитуды мышечного сокращения или даже отсутствие механической реакции мышцы.
Снижение ответной реакции мышцы или ее отсутствие при пессимальном раздражении Н Е. Введенский связывал с понижением лабильности ткани. В результате снижения лабильности падает возбудимость, уменьшается скорость проведения возбуждения, удлиняется рефрактерный период. В итоге снижение лабильности приводит к блоку проведения возбуждения.
Пессимальная реакция, по Н. Е. Введенскому, развивается в синапсе как наиболее низколабильном образовании.
Современные представления о механизмах синаптической передачи позволяют трактовать механизм оптимума и пессимума следующим образом. При оптимальной частоте раздражителя каждый последующий нервный импульс попадает в фазу убывания постсинаптического потенциала, вызванного предыдущим нервным импульсом.
Препарат 1.Миелиновые нервные волокна. Седалищный нерв лягушки. Обработка осмиевой кислотой.
При малом увеличении видны отдельные и собранные в пучки миелиновые нервные волокна. Надо выбрать отдельно лежащее, не поврежденное после расщипывания нерва миелиновое волокно и просмотреть его по всей длине. Толщина его неодинакова. Через некоторые интервалы оно многократно истончается, образуя на поверхности короткие узловые перехваты, или перехваты Ранвье. Надо изучить строение участка миелинового волокна с перехватом Ранвье при большом увеличении. Миелиновое нервное волокно состоит из сравнительно толстого, светлого осевого цилиндра (1), занимающего его центральную часть. Осевой цилиндр может быть аксоном двигательного или дендритом чувствительного нейрона. Он одет миелиновой оболочкой (2), интенсивно окрашенной осмиевой кислотой в темно-коричневый цвет. Эта оболочка образована леммоцитами — клетками, гомологичными олигодендроцитам центральной нервной системы. Леммоциты (3) располагаются цепочками. Их границы соответствуют перехватам Ранвье (4). Отрезок миелинового волокна, заключенный между перехватами, называют сегментом (междоузлием); его оболочка представлена одной глиальной клеткой с ядром (5), лежащим в ее наружной части, приблизительно в центре сегмента. Миелиновая оболочка имеет светлые, в виде косых линий структуры — насечки нервного волокна, или насечки Шмидта—Лантермана (6).
Электронная микроскопия показала, что эти насечки являются прослойками цитоплазмы между завитками мезаксона. В области контакта леммоцитов миелиновая оболочка прерывается, нервное волокно суживается, образуя перехват Ранвье. Область перехвата имеет большое значение для доступа к осевому цилиндру необходимых веществ, находящихся в тканевой жидкости. Поверхность нервного волокна покрыта тонкой соединительной мембраной. В оболочке миелинового нервного волокна различают внутренний толстый и наружный (7) тонкий слой, состоящий из цитоплазмы леммоцитов и их ядер. Проведение импульса по нервному волокну связано с функцией аксолеммы. Миелиновые нервные волокна характеризуются быстрым и точным проведением нервных импульсов, что объясняют относительной толщиной осевого цилиндра, наличием перехватов Ранвье и сильным развитием миелинового слоя. Миелиновые нервные волокна обычно располагаются пучками, входят в состав периферических нервов или образуют белое вещество—проводящие пути спинного и головного мозга. Миелиновые волокна центральной нервной системы имеют оболочку, образованную олигодендроцитами. В них отсутствуют перехваты Ранвье, насечки Шмидта—Лантермана и соединительнотканная базальная мембрана, поскольку роль соединительной ткани в мозге выполняет нейроглия, в частности олигодендроглия. Физиологическая регенерация леммоцитов обеспечивается процессами, происходящими в активном центре — перикарионе нервной клетки. Синтезируемые вещества (в частности, белки) и образующиеся органеллы (митохондрии) непрерывно оттекают вместе с нейроплазмой по аксону. Разрушившиеся в процессе функциональной активности нейрона завитки мезаксона заменяются новыми в результате синтетических процессов в леммоцитах. В случае нарушения, задержки или прекращения синтетических процессов в леммоцитах разрушение миелиновых пластинок продолжается, а замена новыми отсутствует. Следствием этого является демиелинизация нервных волокон, сопровождающаяся тяжелейшими расстройствами их функции. Репаративная регенерация миелинового нервного волокна возможна только при сохранности тела нейрона.
Препарат 2. Безмиелиновые нервные волокна. Симпатический нерв. Окраска гематоксилином и эозином.
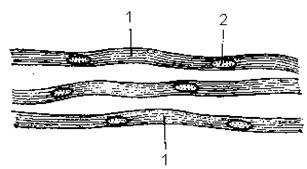 |
При малом увеличении надо выбрать расщипанные пучки нервных волокон и изучить строение нервного волокна при большом увеличении. Безмиелиновое нервное волокно значительно тоньше миелинового и, как показывает название, не имеет миелиновой оболочки. Оно состоит из тонких осевых цилиндров (1), являющихся нейритами двигательных или вставочных нейронов и леммоцитов (шванновских клеток), располагающихся цепочкой. Световая микроскопия не выявляет границ шванновских клеток и их отношений с осевыми цилиндрами. Безмиелиновая оболочка видна как однородный тяж цитоплазмы, содержащей ядра (2) овальной формы, одевающей несколько осевых цилиндров. Цитоплазма леммоцитов образует шванновскую оболочку. Снаружи нервное волокно одето соединительнотканной базальной мембраной. Осевые цилиндры располагаются рыхло и переходят в. соседние безмякотные волокна. Они не изолированы так тщательно, как в мякотных нервных волокнах, что связано с особенностями иннервации внутренних органов. Электронная микроскопия дает представление об иных, чем в миелиновом волокне, морфофункциональных отношениях осевых цилиндров и леммоцитов. Осевые цилиндры (1), принадлежащие различным нейронам, погружаются в цитоплазматический тяж, образованный леммоцитами (2), оболочка которых (3) охватывает их и образует складки (4). На дне этих складок располагаются отростки нейронов. Последние отграничены от цитоплазмы (5) леммоцитов как их собственной мембраной, так и мембраной шванновской клетки. Ядра (6) леммоцитов располагаются центрально. Физиологическая регенерация безмиелинового волокна обеспечивается процессами, происходящими в теле нейрона и в леммоцитах (внутриклеточная физиологическая регенерация). Репаративная регенерация возможна лишь при сохранности тела нейрона.
При малом увеличении (А) видна поверхность нерва, покрытая соединительнотканной оболочкой — эпиневрием (1), в котором находятся кровеносные сосуды (2) и жировые клетки (3). Плотная соединительная ткань
делит нервы на отдельные пучки различной величины, округлой или овальной формы. От оболочки нервного пучка — периневрия (4) идут прослойки рыхлой соединительной ткани, окружающей группы и отдельные нервные волокна. Соединительнотканные образования внутри нервного пучка называются эндоневрием (5). Пучки состоят из нервных волокон, в состав которых входят длинные отростки различных в функциональном и морфологическом отношении нейронов. Нервные волокна располагаются не строго параллельно и оказываются перерезанными поперек (6) и косо. Надо изучить строение нервных волокон в поперечном сечении при большом увеличении (Б). Нервные волокна отличаются толщиной, цветом и строением. В наиболее толстых миелиновых волокнах различают миелиновую оболочку (1), имеющую вид периферически расположенного черного кольца, и внутреннюю светлую часть, представленную осевым цилиндром (2). Миелиновая оболочка нервных волокон, лежащих в центральных отделах пучка, может быть бесцветной или светло-серого цвета, что объясняется неравномерным проникновением осмиевой кислоты при фиксации. Иногда видна в виде тонкого ободка цитоплазма (3) шванновских клеток, свободная от миелина. Попавшие в срез ядра (4) леммоцитов округлой формы прилежат к миелиновой оболочке. Встречаются безмиелиновые волокна (5); в светлой цитоплазме их леммоцитов иногда видны осевые цилиндры в виде мелких точек.
Дата добавления: 2015-09-04 ; просмотров: 2368 . Нарушение авторских прав
Читайте также: