
Нервная регуляция онтогенеза это
Нервная регуляция начинается с закладки отделов ЦНС и продолжается в течение жизни особи.
Взаимодействие между центрами ЦНС и иннервируемыми органами устанавливается на ранних этапах эмбриогенеза, причем эти структуры взаимно стимулируют развитие друг на друга. Отходящие от центров ЦНС периферические нервы подрастают к зачаткам органов и стимулируют их развитие. Отсутствие периферических нервов или их повреждение (например лекарственными препаратами, токсинами токсоплазмы и др.) вызывает нарушение формирования иннервируемых ими структур. Так, например, в Европе родилось несколько сотен детей с отсутствием конечностей, матери которых в период беременности принимали снотворное талидомид, блокирующее рост периферических нервов.
В постнатальном периоде сохраняется взаимосвязь между нервной системой и иннервируемыми органами. Родовые травмы головного мозга и периферических нервов приводят не только к параличам, но и к атрофии мышц и отставанию роста соответствующих конечностей или односторонней гипотрофии структур лица (при врожденном параличе VI-VII нервов). Способствуют восстановлению поврежденных структур головного и спинного мозга пассивные движения конечностей (для этого созданы специальные аппараты), массаж и физиотерапевтическая стимуляция иннервируемых органов.
При нейрофиброматозе (аутосомно-доминантный тип наследования) развиваются опухоли периферических нервов. Если заболевание начинается в раннем детстве, то на той стороне тела, где развиваются опухоли, возникает гипертрофия костей и мягких тканей. Например, развивается дизморфоз лица (несимметричное, непропорциональное развитие структур формирующих лицо, рисунок приложения 5).
Установлено, что в раннем детстве игры, способствующие движению кистей рук, особенно мелкие, точные формы деятельности, стимулируют развитие структур головного мозга, в том числе и развитие интеллекта.
На рис. 5 представлены схемы экспериментов на аксолотле по изучению роли периферического нерва в развитии конечностей, а также формирование двигательных центров спинного мозга при отсутствии конечностей. Удаление нерва на левой стороне зародыша аксолотля привело к отсутствию конечности на оперированной стороне тела.
Отсутствие конечности может быть обусловлено действием нейротропных тератогенов (токсины при токсоплазмозе, талидомид и др.) (рис. 5а).
Удаление зачатка конечности у зародыша аксолотля приводит к уменьшению размеров ганглиев и рогов серого вещества спинного мозга на оперированной стороне (рис. 5б).
Разберите рисунки экспериментов по изучению взаимосвязи нервных центров и иннервируемых органов.
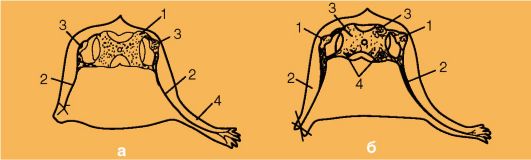
Рис. 5.Взаимосвязь нервных центров и иннервируемых органов (Дьюкар Э., 1978, с изменениями):
а - влияние спинномозговых нервов на развитие конечности: 1 - спинной мозг; 2 - спинномозговой нерв, иннервирующий в конечность; 3 - спинномозговой ганглий; 4 - конечность; б - влияние зачатка конечности на развитие сегментов спинного мозга (поперечный срез зародыша аксолотля с удаленным зачатком конечности: 1 - спинномозговой ганглий; 2 - спинномозговой нерв; 3 - дорсальные рога серого вещества спинного мозга; 4 - вентральные рога серого вещества спинного мозга
Онтогенез – это процесс индивидуального развития организма, включающий в себя систему его эмбрионального и постэмбрионального развития.
Механизмы регуляции онтогенеза
Следует отметить тот факт, что на каждом этапе онтогенеза (особенно эмбрионального периода) происходит реализация наследственной информации, которая сильно зависит от влияния окружающей среды, всей совокупности ее факторов при учете внутренних механизмов развития организма.
Онтогенез, как и любой глобальный процесс жизнедеятельности подчиняется определенной системе регуляции. Разновидностей такой системы выделяют три:
- эмбриональная детерминация;
- эмбриональная индукция;
- нейрогуморальная регуляция окончательного (дефинитивного типа).
Что касается эмбриональной детерминации, то она развивается на основе позиционной информации, которая заложена в яйце. В процессе роста и созревания яйцеклетки, которая находится в материнском организме внутри ее цитоплазмы откладываются различные виды РНК, а также белки – регуляторы, которые предопределяют всю будущую систему развития и дифференцировки клеток.
- Курсовая работа Регуляция онтогенеза 470 руб.
- Реферат Регуляция онтогенеза 280 руб.
- Контрольная работа Регуляция онтогенеза 210 руб.
Неоднородность цитоплазмы яйца задает его полярное положение в яичнике. Один полюс яйца контактирует со стенкой яичника и таким образом питается. Другой полюс яйца направляется в просвет, в котором сосредотачиваются продукты сложного синтеза. Таким образом, положение яйца в яичнике можно назвать оптимальным.
При этом еще до начала активного развития хромосомы ядра находятся в неоднородной среде, которая насыщается биологически активными регуляторами. Уже в яйце можно отследить сочетание хромосомного материала и эпигенетического факторов развития. После оплодотворения зигота вступает в период многократного деления и дочерние клеточные ядра попадают в детерминированные участки цитоплазмы с различными регуляторами. Такие регуляторы становятся внутренними индукторами дифференцировки эмбриональных клеток.
Этап эмбриональной индукции заключается во влиянии одних клеточных зачатков на другие при помощи веществ – регуляторов. Такой механизм регуляции онтогенеза включается на ранних стадиях развития зародыша и, по сути, является эмбриональной гуморальной регуляцией. Индукторы в данном случае выступают в роли первых гормонов. Именно под влиянием таких гормонов начинается дифференцировка нервной системы эмбрионов.
Задай вопрос специалистам и получи
ответ уже через 15 минут!
Нейрогуморальная регуляция онтогенеза – это процесс, который осуществляется сложной системой желёз внутренней секреции и нервной системой эмбриона.
При этом гормоны вырабатываются железами под контролем нервной деятельности, а сама нервная система во многом зависит от внешней среды.
Тем самым вся совокупность факторов, которые в той или иной мере влияют на систему регуляции онтогенеза входят в состав сложнейшей эпигенетической программы развития, так как генная активность одних генов стимулируется, а других достаточно быстро подавляется. Достигается некоторая степень баланса.
Уровни регуляции онтогенеза
Регуляция онтогенеза происходит на следующих уровнях:
- модификация ДНК;
- регуляция на уровне транскрипции;
- регуляция на уровне сплайсинга;
- регуляция на уровне трансляции;
- регуляция на уровне посттрансляционной модификации белков.
Модификация ДНК происходит путем дифференциальной амплификации ДНК или за счет образования политенных хромосом. Регуляция на уровне транскрипции происходит путем транскрипции мРНК. При этом происходит достаточно интенсивное функционирование отдельных генов или цельных блоков генов, что соответствует конкретным этапам развития и клеточной генетической дифференцировки. Для животных регуляторами транскрипции являются стероидные гормоны.
Регуляция на уровне сплайсинга обеспечивает возможность образования различных типов зрелой, активной мРНК, которая образуется на ДНК – матрице.
Сплайсинг – это процесс посттранскрипционной модификации матричной рибонуклеиновой кислоты (происходит по принципу комплементарности).
Процессинг РНК при этом регулируется с помощью рибозимов (которые являются катализаторами рибонуклеиновой природы), а также ферментов матураз. Некоторые генетические заболевания человека очень сильно зависят от нарушений сплайсинга. К ним относят фенилкетонурию и нарушения формирования гемоглобина.
Наконец, регуляция на уровне трансляции обуславливается различной активностью разных типов рибонуклеиновой кислоты (рибосомальной, матричной, транспортной).
Последний тип регуляции РНК заключается в посттрансляционной модификации белков. В данном случае происходят процессы фосфорилирования, ацетилирования, расщепления исходной полипептидной цепи на мельчайшие фрагменты.
Таким образом, процесс регуляции онтогенеза происходит в достаточно сложной системе и под влиянием многих важных веществ – регуляторов. Все вышеописанные примеры говорят о многообразии различных способов регуляции генетической информации и ее продуктов. Также необходимо понимать, что клетке более выгодная регуляция онтогенеза на уровне транскрипции, поскольку в этот момент образуется мРНК, которая в полной мере соответствует белку, и острая потребность в них для клетки отпадает. Таким образом, этот процесс регуляции энергетически выгоден.
Вместе с тем регуляция на уровне транскрипции идет сравнительно медленно, тогда как, например, активация белков путем расщепления молекул-предшественников хотя и неэкономична, но происходит очень быстро. При этом нельзя утверждать, что какие – либо механизмы регуляции онтогенеза имеют большую или меньшую степень значимости. Чаще всего они рассматриваются как равноценные механизмы влияния на процесс дифференцировки генетической информации.
Подводя итог всему вышесказанному, можно сделать вывод о том, что ход онтогенеза во многом определяется генно-регуляторными сетями или каскадами. В ходе регуляции большую роль играют также сигнальные белки и специализированные морфогены, рецепторы.
Так и не нашли ответ
на свой вопрос?
Просто напиши с чем тебе
нужна помощь
Для каждого этапа онтогенеза характерно определенное соотношение активности желез внутренней секреции. В эмбриогенезе определяющую роль играют гормоны плаценты и эндокринные железы организма матери. Они контролируют закладку органов, их рост и развитие в течение того времени, пока формируются железы внутренней секреции плода. Первыми созревают клетки поджелудочной железы, продуцирующие инсулин, и кора надпочечников, которая производит кортикостероиды.
Гормоны этих желез эмбриона начинают регулировать углеводный и минеральный обмен веществ, а также закладки половых желез. В это время формируется тимус, начинается работа иммунной системы плода. Несколько позже начинает функционировать гипофиз и щитовидная железа. Гормон роста регулирует темпы роста всех органов, а гормоны щитовидной железы — энергетический обмен. Недостаток этих гормонов в эмбриогенезе приводит к тяжелым нарушениям физического и психического развития плода. Действие половых гормонов проявляется уже на 10-12 неделе развития эмбриона. В это время закладываются основные признаки, характерные для женского и мужского организмов.
Важно помнить следующее: гормон взаимодействует только с клетками, имеющими рецептор к нему. Таким образом, он может оказывать действие лишь на определенные органы. Кроме того, в разных клетках-мишенях гормоны воздействуют на различные группы генов, и поэтому они могут оказывать разнонаправленное воздействие. Так, сложные морфогенезы в онтогенезе амфибий, обеспечивающие превращение головастика в лягушку, происходят под действием гормонов щитовидной железы, главным образом, тироксина. Его влияние приводит к исчезновению хвоста и жаберных щелей, перестройке черепа, позвоночника и всего пищеварительного тракта, формированию конечностей, изменению строения кожи, в которой появляются многоклеточные слизистые железы. Другими словами, под действием гормона на данном этапе развития меняется вся организация особи.
Нервная регуляция начинается с закладки отделов ЦНС и продолжается в течение жизни особи.
Взаимодействие между центрами ЦНС и иннервируемыми органами устанавливается на ранних этапах эмбриогенеза, причем эти структуры взаимно стимулируют развитие друг на друга. Отходящие от центров ЦНС периферические нервы подрастают к зачаткам органов и стимулируют их развитие. Отсутствие периферических нервов или их повреждение (например лекарственными препаратами, токсинами токсоплазмы и др.) вызывает нарушение формирования иннервируемых ими структур.
В постнатальном периоде сохраняется взаимосвязь между нервной системой и иннервируемыми органами. Родовые травмы головного мозга и периферических нервов приводят не только к параличам, но и к атрофии мышц и отставанию роста соответствующих конечностей или односторонней гипотрофии структур лица.
41) Межклеточные взаимодействия на разных этапах онтогенеза. Эмбриональная индукция и ее виды. Опыты г.Шпемана в изучении явления эмбриональной индукции.
Эмбриональная индукция — взаимодействие между частями развивающегося организма у многоклеточных беспозвоночных и всех хордовых.
Важную роль в эмбриогенезе играют контактные и дистантные взаимодействия.
Контактные взаимодействия – контакт как минимум 2-х бластомеров, являются условием для нормального развития зародыша. Обуславливают дальнейшую судьбу бластомеров, определяя направление перемещения клеточных слоёв, миграцию, подавление деления и т.д. (Сосед определяет судьбу рядом лежащих бластомеров).
Дистантные взаимодействия: в процессах эмбриональной индукции. Взаимодействие частей зародыша, при котором 1 участок определяет судьбу другого, побуждая его к делению. Явление эмбриональной индукции – опыты Грегора Шпемана на амфибиях (1924 г.).
(опыт из рабочей тетради №3)
. Межклеточные взаимодействия чрезвычайно важны в развитии и являются одним из механизмов, обеспечивающих интегрированность развития особи. Этот механизм действует на протяжении всего онтогенеза, но особую значимость имеет на ранних этапах эмбриогенеза, а именно, в период дробления.
Со стадии гаструляции, если в эксперименте у зародыша амфибии взять дорзальную губу бластопора и пересадить её другому зародышу амфибии, но не на спинную, а на вентральную (брюшную) сторону, то развивается 2-я нервная трубка (на брюшной стороне). Вывод: Дорзальная губа бластопора гаструлы у амфибии в норме индуцирует закладку нервной трубки (в норме на спинной/дорзальной стороне).
Для осуществления эмбриональной индукции необходимо:
наличие индуцируемой структуры, отвечающей на действие индуктора;
наличие состояния компетентности (способности воспринимать этот стимул).
Виды эмбриональной индукции:
первичная: обнаруживается первой, при закладке нервной трубки;
вторичная: проявляется на более поздней стадии, чем гаструляция, при закладке всех структур зародыша.;
последующая: при закладке глазного яблока, почек; каждая новая структура последовательно играет роль индуктора;
взаимная: при закладке конечностей.
42) Целостность онтогенеза. Эмбриональная регуляция. Детерминация частей развивающегося зародыша; канализация развития. Концепции морфогенеза (физиологических градиентов, позиционной информации, морфогенетических полей).
Целостность организма — его внутреннее единство, относительная автономность, несводимость его свойств к свойствам отдельных его частей, подчиненность частей целому — проявляется в течение всех стадий онтогенеза. Таким образом, онтогенез представляет собой упорядоченное единство последовательно чередующихся состояний целостности. В целостности индивидуального развития проявляется органическая целесообразность.
Целостность онтогенеза базируется на действии системно-регуляторных факторов: цитогенетических, морфогенетических, морфофизиологических, гормональных, а у большинства животных также нейрогуморальных. Эти факторы, действуя по принципу обратной связи, координируют ход развития и жизнедеятельность организма как активного целого в тесной связи с условиями окружающей среды.
Эмбриональная регуляция – явление восстановления нормального хода развития зародыша после естественного или искусственного его [развития] нарушения.
Детерминация (предопределение) – выбор конкретного пути развития, приобретение клетками способности развиваться в определенном направлении и одновременно ограничение их будущих возможностей развития. В начале эмбриогенеза бластомеры тотипотентны (могут дать начало целому организму) и их развитие зависит от внешних индукторов и соседних клеток. На более поздних стадиях клетки становятся более детерминированными (их развитие предопределено) и они развиваются по намеченному плану.
Процесс развития канализирован; он устойчив к внешним давлениям, которые могли бы заставить его отклониться от нормального пути. Если фенотипический продукт развития адаптивен, то можно предполагать, что отбор благоприятствовал канализированым генотипам, т. е. генотипам, которые приводят к развитию одного и того же признака в различных средах. Таким образом, канализация развития представляет собой консервативную силу в эволюции. Канализированное онтогенетическое развитие устойчиво к радикальному изменению. Генные мутации или рекомбинации, коренным образом изменяющие нормальное развитие, будут элиминироваться. Сохраниться могут только те генетически детерминированные изменения онтогенеза, которые означают относительно небольшие сдвиги в процессе развития.
Морфогенез — это процесс возникновения новых структур и изменения их формы в ходе индивидуального развития организмов. Морфогенез, как рост и клеточная дифференцировка, относится к ациклическим процессам, т.е. не возвращающимся в прежнее состояние и по большей части необратимым.
В настоящее время разрабатывают несколько подходов к проблеме регуляции и контроля морфогенеза:
Концепция физиологических градиентов - интенсивность жизненных процессов неодинакова в разных частях тела: она закономерно понижается по какой-либо оси тела или его органов (Ч. Чайлд). Основным показателем интенсивности жизненных процессов, по Чайлду, является уровень метаболизма, изучаемый по интенсивности окислительно-восстановительных процессов. По мнению автора, количественные различия в уровне метаболизма, или градиенты, имеют значение простейших систем, определяющих интеграцию тех организмов, у которых в эволюции не выработались еще или находятся в примитивном состоянии интеграционные механизмы более высокого порядка, как-то: нервная система, железы внутренней секреции и др. Ч. Чайлд открыл также, что верхний конец градиента является доминирующим. Выделяя некоторые факторы, он подавлял развитие таких же структур из других клеток зародыша. Наряду с подтверждающими имеются явления, которые не укладываются в упрощенную схему, и поэтому концепцию Чайльда нельзя рассматривать как универсальное объяснение пространственной организации развития.
Более современной является концепция позиционной информации, по которой клетка как бы оценивает свое местоположение в координатной системе зачатка органа, а затем дифференцируется в соответствии с этим положением. По мнению современного английского биолога Л. Вольперта, положение клетки определяется концентрацией некоторых веществ, расположенных вдоль оси зародыша по определенному градиенту. Ответ клетки на свое местоположение зависит от генома и всей предыдущей истории ее развития. По мнению других исследователей, позиционная информация есть функция полярных координат клетки. Существует также мнение о том, что градиенты представляют собой стойкие следы периодических процессов, распространяющихся вдоль развивающегося зачатка. Концепция позиционной информации позволяет формально интерпретировать некоторые закономерности онтогенетического развития, но она очень далека от общей теории целостности.
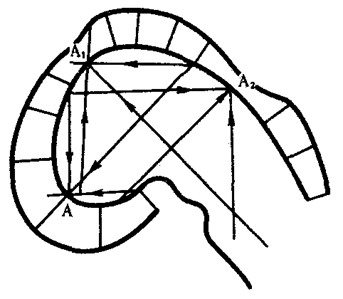
Концепция морфогенетических полей, базирующаяся на предположении о дистантных либо контактных взаимодействиях между клетками зародыша, рассматривает эмбриональное формообразование как самоорганизующийся и самоконтролируемый процесс. Предыдущая форма зачатка определяет характерные черты его последующей формы. Кроме того, форма и структура зачатка способны оказать обратное действие на биохимические процессы в его клетках. Наиболее последовательно эту концепцию разрабатывал в 20—30-х гг. отечественный биолог А. Г. Гурвич, предложивший впервые в мировой литературе математические модели формообразования. Он, например, моделировал переход эмбрионального головного мозга из стадии одного пузыря в стадию трех пузырей.
Модель исходила из гипотезы об отталкивающих взаимодействиях между противоположными стенками зачатка. На рис. 8.17 эти взаимодействия отображены тремя векторами (А, А1, А2). Гурвич впервые указал также на важную роль неравновесных надмолекулярных структур, характер и функционирование которых определяются приложенными к ним векторами поля. В последние годы К. Уоддингтон создал более обобщенную концепцию морфогенетического векторного поля, включающую не только формообразование, но и любые изменения развивающихся систем.
43) Влияние факторов среды на онтогенез. Критические периоды в онтогенезе человека. Тератогенез и канцерогенез. Понятие аномалий и пороков развития. Значение нарушений частных и интегративных механизмов онтогенеза в формировании ВПР.
Среди факторов среды различают:
Под биологическими факторами понимают взаимодействие живых организмов. Абиологические факторы - это факторы неживой природы (климат и пр.).
Факторы могут быть:
Однако даже при кратковременном воздействии они могут оказать весьма существенное влияние на развитие организма.
Онтогенез представляет собой последовательное развитие, при котором ранее образовавшиеся структуры обусловливают развитие последующих, причем и эта тенденция проявляется в единстве с условиями среды. При одном и том же генотипе развиваются различные фенотипические особенности. Однако совершенно очевидно, что в фенотипе не может быть ни одного признака, который не был бы обусловлен генотипом. Из этого вытекает, что именно противоречивое единство внутренних и внешних факторов определяет развитие организма.
Критические периоды эмбриогенеза человека – зародыш наиболее чувствителен к действию факторов внешней среды, т.к. изменяются условия его существования (включаются новые блоки генов):
Имплантация (6-7 сутки после оплодотворения);
Плацентация (14-15 сутки после оплодотворения);
Роды (38-40 неделя после оплодотворения).
Действие неблагоприятных факторов среды в эти периоды развития приводит к порокам развития данных систем.
Тератогенез - это возникновение пороков развития под влиянием факторов внешней среды (тератогенных факторов). Тератогенные факторы включают лекарственные средства, наркотики и многие другие вещества.
Канцерогенез – это процесс появления злокачественной клетки.
Пороки развития (синоним аномалии развития) — собирательный термин, обозначающий отклонения от нормального строения организма вследствие нарушения внутриутробного или постнатального (реже) развития.
Эти изменения вызывают нарушения соответствующих функций. Под аномалиями развития и понимают только такие пороки, при которых анатомические изменения не приводят к существенному нарушению функций, например деформации ушных раковин, не обезображивающие лица больного и существенно не отражающиеся на восприятии звуков. Грубые пороки развития, при которых обезображивается внешний облик ребенка, нередко называют уродствами.
1) эндогенные (внутренние) факторы:
а) изменения наследственных структур (мутации);
б) "перезревание" половых клеток; в) эндокринные заболевания;
г) влияние возраста родителей;
2) экзогенные (внешние) факторы:
а) физические - радиационные, механические воздействия;
б) химические - лекарственные препараты, химические вещества, применяемые в промышленности и в быту, гипоксия, неполноценное питание, нарушения метаболизма;
в) биологические - вирусные заболевания, протозойные инвазии, изоиммунизация.
Формирование пороков происходит преимущественно в период эмбрионального морфогенеза (3-10-я неделя беременности) в результате нарушения процессов размножения, миграции, дифференциации и гибели клеток. Эти процессы происходят на внутриклеточном, экстраклеточном, тканевом, межтканевом, органном и межорганном уровнях. Нарушением размножения клеток объясняют гипоплазию и аплазию органов. Нарушение их миграции лежит в основе гетеротопий. Задержка дифференциации клеток обусловливает незрелость или персистирование эмбриональных структур, а ее полная остановка - аплазию органа или его части. Нарушение физиологической гибели клеток, как и нарушение механизмов адгезии ("склеивание" и срастание эмбриональных структур), лежат в основе многих дизрафий (например, спинномозговых грыж).
Биология развития изучает способы генетического контроля индивидуального развития и особенности реализации генетической программы в фенотип в зависимости от условий. Под условиями понимаются различные внутриуровневые и межуровневые процессы и взаимодействия – внутриклеточные, межклеточные, тканевые, внутриорганные, организменные, популяционные, экологические.
Очень важными являются исследования конкретных онтогенетических механизмов роста и морфогенеза. К ним относятся процессы пролиферации (размножения) клеток, миграции (перемещения) клеток, сортировки клеток, их запрограммированной гибели, дифференцировки клеток, контактных взаимодействий клеток (индукция и компетенция), дистантного взаимодействия клеток, тканей и органов (гуморальные и нервные механизмы интеграции). Все эти процессы носят избирательный характер, т.е. протекают в определенных пространственно-временных рамках с определенной интенсивностью, подчиняясь принципу целостности развивающегося организма. Поэтому одной из задач биологии развития является выяснение степени и конкретных путей контроля со стороны генома и одновременно уровня автономности различных процессов в ходе онтогенеза.
Большую роль в процессах онтогенеза играет делениеклеток, поскольку:
– благодаря делению из зиготы, которая соответствует одноклеточной стадии развития, возникает многоклеточныйорганизм;
– пролиферация клеток, происходящая после стадии дробления, обеспечивает рост организма;
– избирательному размножению клеток принадлежит заметная роль в обеспечении морфогенетических процессов.
В постнатальном периоде индивидуального развития благодаря клеточному делению осуществляется обновление многих тканей в процессе жизнедеятельности организма, а также восстановление утраченных органов, заживление ран.
Исследования показали, что количество циклов клеточных делений в ходе онтогенеза генетически предопределено. Однако известна мутация, которая изменяет размеры организма за счет одного дополнительного клеточного деления. Эта мутация описана у Drosophila melanogaster, она наследуется по рецессивному сцепленному с полом типу. У таких мутантов развитие протекает нормально на протяжении всего эмбрионального периода. Но в тот момент, когда нормальные особи окукливаются и начинают метаморфоз, особи-мутанты продолжают оставаться в личиночном состоянии еще дополнительно 2–5 суток. За это время у них происходит 1–2 дополнительных деления в имагинальных дисках, от количества клеток которых зависит размер будущей взрослой особи. Затем мутанты образуют куколку вдвое крупнее обычной. После метаморфоза несколько удлиненной по времени стадии куколки на свет появляется морфологически нормальная взрослая особь удвоенного размера.
Описан ряд мутаций у мышей, вызывающих снижение пролиферативной активности и следующие за этим фенотипические эффекты – микрофтальмия (уменьшение размеров глазных яблок), отставание роста и атрофия некоторых внутренних органов из-за мутаций, затрагивающих центральную нервную систему.
Таким образом, деление клеток является чрезвычайно важным процессом в онтогенетическом развитии. Оно протекает с разной интенсивностью в разное время и в разных местах, носит клональный характер и подвержено генетическому контролю. Все это характеризует клеточное деление как сложнейшую функцию целостного организма, подчиняющегося регулирующим влияниям на различных уровнях: генетическом, тканевом, онтогенетическом.
Миграция клеток имеет очень большое значение, начиная с процесса гаструляции и далее в процессах морфогенеза. Нарушение миграции клеток в ходе эмбриогенеза приводит к недоразвитию органов или к их гетеротопиям, изменениям нормальной локализации. Все это представляет собой врожденные пороки развития. Например, нарушение миграции нейробластов приводит к возникновению островков серого вещества в белом веществе, при этом клетки утрачивают способность к дифференцировке. Более выраженные изменения миграции приводят к микрогирии и полигирии (большое число мелких и аномально расположенных извилин больших полушарий), либо к макрогирии (утолщение основных извилин), или же к агирии (гладкий мозг, отсутствие извилин и борозд больших полушарий). Все эти изменения сопровождаются нарушением цитоархитектоники и послойного строения коры, гетеротопиями нервных клеток в белом веществе. Подобные пороки отмечены и в мозжечке.
Для миграции клеток очень важны их способность к амебоидному движению и свойства клеточных мембран. Все это генетически детерминировано, следовательно, и сама миграция клеток находится под генетическим контролем, с одной стороны, и влияниями окружающих клеток и тканей – с другой.
Агрегация клеток зародышевых листков с себе подобными объясняется способностью к избирательному слипанию (адгезии) клеток одного типа между собой. Одновременно это является проявлением ранней дифференцировки клеток на стадии гаструлы.
Избирательная сортировка клеток возможна за счет того, что контакты между подобными клетками сильнее, чем между чужеродными клетками из-за различий в поверхностном заряде их мембран. Установлено, что поверхностный заряд клеток мезодермы ниже, чем клеток экто- и энтодермы, поэтому клетки мезодермы легче деформируются и втягиваются в бластопор в начале гаструляции. Есть также мнение, что контактные взаимодействия между одинаковыми клетками основываются на антигенных свойствах их мембран.
Избирательная адгезия клеток определенного зародышевого листка друг с другом является необходимым условием нормального развития организма. Примером потери клетками способности к избирательной сортировке и слипанию является их беспорядочное поведение в злокачественной опухоли. По-видимому, в обеспечении сортировки клеток важное место принадлежит генетическим механизмам.
Дифференцировка клеток – это постепенное (на протяжении нескольких клеточных циклов) возникновение все больших различий и направлений специализации между клетками, происшедшими из более или менее однородных клеток одного зачатка. Этот процесс сопровождают морфогенетические преобразования, т.е. возникновение и дальнейшее развитие зачатков определенных органов в дефинитивные органы. Первые химические и морфогенетические различия между клетками, обусловленные самим ходом эмбриогенеза, обнаруживаются в период гаструляции.
Процесс, в результате которого отдельные ткани в ходе дифференцировки приобретают характерный для них вид, называется гистогенезом. Дифференцировка клеток, гистогенез и органогенез совершаются в совокупности, причем в определенных участках зародыша и в определенное время. Это свидетельствует о координированности и интегрированности эмбрионального развития.
В настоящее время общепринятой считается точка зрения на дифференцировку клеток в процессе онтогенеза как на результат последовательных реципрокных (взаимных) влияний цитоплазмы и меняющихся продуктов активности ядерных генов. Таким образом, впервые прозвучала идея о дифференциальной экспрессии генов как основном механизме цитодифференцировки. Уровни регуляции дифференциальной экспрессии генов соответствуют этапам реализации информации в направлении ген → полипептид → признак и включают не только внутриклеточные процессы, но и тканевые и организменные.
Эмбриональная индукция – это взаимодействие частей развивающегося зародыша, при котором один участок зародыша влияет на судьбу другого участка. В настоящее время установлено, что первичным эмбриональным индуктором является хордомезодермальный зачаток в спинной губе бластопора. Но явления индукции многочисленны и разнообразны. Кроме первичной индукции, различают вторичные и третичные, которые могут происходить на более поздних, чем гаструляция, этапах развития. Все эти индукции представляют собой каскадные взаимодействия, потому что индукция многих структур зависит от предшествующих индукционных событий. Например, глазной бокал возникает только после развития передней части головного мозга, хрусталик – после формирования бокала, а роговица – после образования хрусталика.
Индукция носит не только каскадный, но и переплетающийся характер, т.е. в индукции той или иной структуры может участвовать не одна, а несколько тканей. Например, глазной бокал служит главным, но не единственным индуктором хрусталика.
Различают два вида индукции. Гетерономная индукция – когда один кусочек зародыша индуцирует иной орган (хордомезодерма индуцирует появление нервной трубки и всего зародыша в целом). Гомономная индукция – индуктор побуждает окружающий материал к развитию в том же направлении, что и он сам. Например, область нефротома, пересаженная другому зародышу, способствует развитию окружающего материала в сторону формирования головной почки, а прибавление в культуру фибробластов сердца маленького кусочка хряща влечет за собой процесс образования хряща.
Для того чтобы воспринять действие индуктора, компетентная ткань должна обладать хотя бы минимальной организацией. Одиночные клетки не воспринимают действие индуктора, а чем больше клеток в реагирующей ткани, тем активнее ее реакция. Для оказания индуцирующего действия иногда достаточно лишь одной клетки индуктора. Установлена химическая природа индукторов – это могут быть белки, нуклеопротеиды, стероиды и даже неорганические вещества. Но специфичность ответа прямо не связана с химическими свойствами индуктора.
Таким образом, генетический контроль онтогенеза очевиден, однако в процессе развития зародыш и его части обладают способностью к саморазвитию, регулируемому самой целостной развивающейся системой и не запрограммированному в генотипе зиготы.
2. Ведущая роль ядра в регуляции формообразования
Реализация наследственной информации в онтогенезе многоступенчатый процесс. Он включает в себя различные уровни регуляции – клеточный, тканевый, организменный. На каждом этапе развития организма функционирует большое количество генов. Каждый из них контролирует ход той или иной биохимической реакции и через нее принимает участие в осуществлении формообразовательных процессов. Локализация генов в хромосомах ядер определяетведущую роль ядра в регуляции формообразования. Однако по этому поводу длительное время происходили дискуссии, в особенности между эмбриологами и генетиками. Первые отводили основную роль цитоплазме, вторые – ядру. Затем был найден компромиссный вариант, согласно которому ядро отвечает за видоспецифические признаки организмов, а цитоплазма – за более общие признаки.
Правота генетиков была продемонстрирована лишь в 30-е годы ХХ века в опытах физиолога растений Г.Хеммерлинга. Он обнаружил, что у одноклеточной водоросли АсеtаЬulаriа форма шляпки (зонтика) – органа размножения, развивающегося на верхушке стебля, зависит только от ядра. Так, если у водоросли одного вида – АсеtаЬulаriа mediterranea удалить содержащий ядро ризоид и срастить со стебельком ризоид с ядром другого вида – А. wettsteini или А. crenulata, то образуется шляпка, свойственная А. wettsteini или А.crenulata , и наоборот (рис. 15).
В 50-е годы ХХ в. Б.Л.Астауров использовал для доказательства ведущей роли ядра в развитии животных разную чувствительность ядра и цитоплазмы к действию радиации – ядро во много раз чувствительнее к облучению, чем цитоплазма. Исследования проводились на яйцах тутового шелкопряда. Яйца, лишенные женского ядерного аппарата (путем облучения высокой дозой рентгеновских лучей), при оплодотворении необлученной спермой образуют ядро дробления посредством слияния ядер двух спермиев. Соответствующие особи всегда самцы и их легко узнают при помощи генетической маркировки. Если, используя эту методику, соединить цитоплазму яиц одного вида с ядром яиц другого вида тутового шелкопряда, отличающимся по многим морфологическим, физиологическим признакам и поведению, то оказывается, что развивающийся организм целиком и полностью подобен отцовскому, т.е. соответствует информации, содержащейся в ядре.
Аналогичные исследования проводились и с позвоночными животными. Первым этот вопрос исследовал французский эмбриолог К.Гальен-младший. Он использовал метод трансплантации ядер в яйцеклетки амфибий, который, как считают, разработан американскими эмбриологами Бриггсом и Кингом в 50-е годы прошлого столетия и позднее усовершенствован английским ученым Джоном Гердоном. В действительности этот метод был разработан еще в 40-е годы ХХ в. русским ученым, основоположником отечественной экспериментальной эмбриологии Георгием Викторовичем Лопашовым. Суть метода заключается в том, что собственное ядро яйцеклетки удаляется и чужеродное ядро–донор впрыскивается в яйцеклетку.
Именно путем межвидовых пересадок ядер Гальен получил ядерно-цитоплазматические гибриды с разной конституцией. Начиная со стадии ранней гаструлы, у них обнаруживались тяжелые нарушения развития. Однако небольшое число таких гибридов (около 2%) достигает взрослого состояния. Все особи по своим признакам подобны представителям того вида, от которого взято трансплантированное ядро.
Таким образом, можно утверждать, чтоспецифические особенности индивидуального развития контролируются клеточным ядром.
Ядро, несущее наследственный материал, в котором записана программа индивидуального развития, характеризуется следующими особенностями:
– играет ведущую роль в регуляции формообразовательных процессов.
– осуществляет эту роль посредством ядерно-цитоплазматических взаимоотношений, т.е. разная цитоплазма индуцирует разные функциональные состояния находящегося в клетке ядра.
– в ходе регуляции индивидуального развития проявляет периодичность морфогенетической активности.

Рис. 15. Эксперименты Хеммерлинга, доказывающие выработку ядром ацетобулярии вещества, необходимого для регенерации шляпки (Л.И.Корочкин,1999)

Папиллярные узоры пальцев рук - маркер спортивных способностей: дерматоглифические признаки формируются на 3-5 месяце беременности, не изменяются в течение жизни.

Общие условия выбора системы дренажа: Система дренажа выбирается в зависимости от характера защищаемого.

Опора деревянной одностоечной и способы укрепление угловых опор: Опоры ВЛ - конструкции, предназначенные для поддерживания проводов на необходимой высоте над землей, водой.

Организация стока поверхностных вод: Наибольшее количество влаги на земном шаре испаряется с поверхности морей и океанов (88‰).
Читайте также: