
Нервная система и энергообеспечение
Основной путь получения энергии — аэробный распад глюкозы по ГБФ-пути. Глюкоза — почти единственный энергетический субстрат, поступающим в нервную ткань, который может быть использован ее клетками для образования АТФ.
Проникновение глюкозы в ткань мозга не зависит от действия инсулина, который не проникает через гематоэнцефалический барьер. Влияние инсулина проявляется лишь в периферических нервах.
Постоянный и непрерывный приток глюкозы и кислорода из кровеносного русла — необходимое условие энергетического обеспечения нервных клеток, так как содержание гликогена в нервной ткани ничтожно (0,1% от массы мозга) и не может обеспечить мозг энергией даже на короткое время.
Глюкоза — основной источник энергии, так как через ГЭБ в нервные клетки поступает только глюкоза, которая, расщепляясь в аэробном гликолизе, образует ПВК, превращающуюся с помощью пируватдегидрогеназного комплекса в ацетил-КоА, который вступает в ЦТК, давая восстановленные эквиваленты для окислительного фосфорилирования, приводящего к образованию АТФ. В отличие от других тканей организма человека ВЖК не проникают через ГЭБ и не могут быть использованы в качестве энергетического материала. В пируватдегидрогеназный и -кетоглутаратдегидрогеназный комплексы входит витамин В1 в виде тиаминпирофосфата, поэтому недостаток витамина В1 в первую очередь сказывается на функции нервной системы, в клетках которой будет нарушено образование АТФ. Это приводит к возникновению полиневритов. При голодании, сахарном диабете нервная ткань использует кетоновые тела в качестве энергетического материала.
Высокая скорость потребления глюкозы нервными клетками обеспечивается работой высокоактивной гексокиназы мозга. Здесь гексокиназа не является ключевым ферментом всех путей метаболизма глюкозы. Ключевые ферменты ГБФ-пути в нервной ткани — фосфофруктокиназа и изоцитратдегидрогеназа.
Фосфофруктокиназу ингибируют фруктозо-1,6-бифосфат, АТФ и цитрат, активируют фруктозо-6-фосфат, АДФ, АМФ и неорганический фосфат. Активность изоцитрат ДГ даже при нормальном уровне утилизации глюкозы в состоянии покоя максимальна. Поэтому при повышенном энергопотреблении нет возможностей ускорения реакций цикла трикарбоновых кислот.
Образование НАДФН2, используемого в основном для синтеза жирных кислот и стероидов, обеспечивается сравнительно высокой скоростью протекания ГМФ-пути распада глюкозы.
Энергия АТФ в нервной ткани используется неравномерно во времени. Резкое повышение энергозатрат происходит при очень быстром переходе от сна к бодрствованию.
Образование креатинфосфата способно удерживать макроэргические связи. Реакция полностью обратима, ее направление зависит от соотношения АТФ/АДФ в клетках нервной ткани. Во время сна накапливается фосфокреатин. Переход к бодрствованию приводит к резкому уменьшению концентрации АТФ — равновесие реакции сдвигается влево, т.е. образуется АТФ.
Метаболизм аминокислот и белков
Ткань мозга интенсивно обменивается аминокислотами с кровью. Для этого существуют специальные транспортные системы: две для незаряженных и еще несколько — для аминокислот, заряженных положительно и отрицательно. Концентрация свободных аминокислот в нервной ткани в 8 раз больше, чем в крови. Белки в головном мозге находятся в динамическом состоянии. Велика активность АсТ и АлТ, переводящих аминокислоты в кето-, для получения субстратов ЦТК. Белки серого вещества и мозжечка характеризуются высокой скоростью обновления особенно возбуждающих агентов (электрический ток, фармсредства), однако под влиянием наркоза, эти процессы затухают.
До 75% от общего количества аминокислот нервной ткани составляют аспартат, глутамат, а также продукты их превращений или вещества, синтезированные с их участием (глутамин, ацетильные производные, ГАМК (гамма-аминомасляная кислота) глутатион).
8. ЭНЕРГООБЕСПЕЧЕНИЕ СПЕЦИФИЧЕСКИХ ФУНКЦИЙ НЕРВНОЙ ТКАНИ
Изучение суммарных процессов окисления и образования энергопродукции в мозге представляет собой одну сторону проблемы; другая сторона – выявление специфических процессов в нервной ткани, требующих энергетических затрат. Характеристика этих процессов остается до настоящего времени во многом загадочной.
Еще в ранних работах, выполненных П. Мак-Ильвейном и другими исследователями на изолированных нервах, ганглиях или срезах мозга, установлено, что электростимуляция препаратов сопровождалась усилением потребления кислорода и глюкозы, причем обнаружена прямая зависимость между частотой электрической импульсации и степенью интенсификации окислительных процессов. Электрическое раздражение вызывает резкое и быстрое снижение уровня АТФ, вслед за которым уменьшается содержание креатинфосфата и накапливается неорганический фосфат; после прекращения электростимуляции препаратов в первую очередь восстанавливается уровень АТФ.
Впоследствии аналогичные данные об изменении уровня основных макроэргических соединений при изменении функционального состояния нервной системы были получены in vivo в экспериментах с мозгом целостных животных. Показано заметное ускорение расходования АТФ и креатинфосфата при возбуждении – условно-рефлекторном или вызванном фармакологическими препаратами; и, напротив, замедление использования этих соединений при торможении или наркозе. В этих экспериментах на интактных животных установлено, что при усилении энергозатрат в мозге сначала уменьшается уровень запасных энергетических субстратов – креатинфосфата и гликогена и лишь после исчерпания этих источников начинает быстро снижаться уровень АТФ; в период восстановления уровень АТФ первым возвращается к исходным значениям, а затем нормализуется содержание креатинфосфата и гликогена.
В последние годы для исследования интенсивности энергетического метаболизма различных структур мозга широко применяется радиоактивный дериват глюкозы – 2-дезоксиглюкоза; теоретическое обоснование использования этого соединения сделано в лаборатории американского нейрохимика Л. Соколова. Метод основан на том, что дезоксиглюкоза поглощается мозгом и вступает в гексокиназную реакцию со скоростью, прямо пропорциональной скорости использования глюкозы. Однако дальнейшие метаболические превращения дезок-сиглюкозо-6-фосфата в мозге практически не происходят.
Использование С- или дезоксиглюкозы с последующей авторадиографией срезов мозга позволило получить более детальное представление о поглощении глюкозы и интенсивности энергетического метаболизма в самых разных структурах мозга. Была установлена тесная корреляция между интенсивностью энергетического обмена и функциональной активностью определенных структур мозга в экспериментах, где контролем служили аналогичные структуры контрлатерального полушария того же самого животного. Например, обнаружено снижение на 35–60% потребления глюкозы структурами слуховой системы или зрительной системы после соответствующей депривации.
Подобные исследования дают представление лишь об итоговых, балансовых изменениях важнейших компонентов энергетического обмена, оставляя неясными количественные характеристики энергозатрат на специфические процессы, присущие только нервной ткани, интенсивность которых меняется при изменении функционального состояния. К сожалению, в настоящее время нет еще исчерпывающего ответа на один из кардинальных вопросов нейрохимии и нейрофизиологии: какие конкретные биохимические реакции лежат в основе целого ряда функций нервной ткани. Многие стороны этой важной проблемы нуждаются в уточнениях и дальнейших углубленных исследованиях. Некоторые специфические энергозависимые функции нервной ткани и биохимические процессы, лежащие в их основе, в общих чертах суммированы в табл. 8.
Одной из основных функций нервной ткани является передача импульсов от одного нейрона к другому. Толчком к расшифровке взаимосвязи между энергетическим метаболизмом и этой функцией послужили работы А. Ходжкина, установившего, что необходимым условием для прохождения импульсов по нервному волокну служит неравномерное распределение ионов натрия и калия по разным сторонам клеточной мембраны. Поддержание ионной асимметрии, восстановление ее после прохождения нервного импульса связано со значительными энергетическими затратами; прежде всего это относится к транспорту ионов натрия против градиента концентрации в момент перехода потенциала действия в потенштал покоя. Особое значение в этом процессе принадлежит К + , Na^-стимулируемой АТФазе.
Таблица 8. Основные энергозависимые процессы, лежащие в основе специфических функций нервной ткани
К + , № + -АТФазная реакция
Детально изучены свойства этого важнейшего фермента, определена роль липидов мембран в его активации. Установлено, что активность К + , фазы в головном мозге заметно выше, чем во многих других тканях, причем максимальная активность фермента обнаружена в коре больших полушарий, меньшая – в коре мозжечка и таламусе, затем – в экстрапирамидальных ядрах; минимальная активность найдена в белом веществе. Активность фермента значительно возрастает в ходе формирования и окончательного созревания мозга; например, у крыс в интервале между 5-м днем до рождения и 60-м днем постнатального развития она увеличивается в 10 раз.
Таким образом, обнаруживается четкий – параллелизм между повышением энергетических потребностей в ходе созревания мозга и увеличением активности фермента, обеспечивающего энергетику усиливающейся импульсавди. Следует добавить, что энергия, требующаяся на прохождение одного нервного импульса в мозге взрослых животных, гораздо выше, чем у новорожденных.
Сопоставление средней частоты прохождения нервных импульсов и объема, требующегося для обеспечения импульсации трансмембранного переноса ионов натрия и калия, со скоростью синтеза макроэргических соединений дает возможность приблизительно оценить затраты энергии на осуществление этой важнейшей функции нервной ткани. По расчетам М.И. Прохоровой, при стационарном состоянии эти затраты составляют около 10–15% от общего количества АТФ, образующегося в мозге за единицу времени; при изменении функционального состояния, особенно при возбуждении, расход АТФ возрастает. В работах других исследователей приводятся более высокие цифры; например, для коры больших полушарий мозга крыс общие затраты на трансмембранный перенос ионов натрия составляют около 40, для гиппокампа – 55%.
К специфическим функциям нервной ткани – относятся также процессы хранения и переработки информации, поступающей в головной мозг. Синтез специфических белков и нейро-пептидов, компонентов липо- и гликопротеидных комплексов, участвующих в реализации отдельных этапов хранения и переработки информации, в процессе консолидации временных связей требует значительных энергетических затрат. Интенсивность синтеза ряда белков и пептидов мозга существенно превышает скорость биосинтеза белков в других тканях и заметно изменяется при нарушении энергетического обмена, вызванного недостатком кислорода.
Процессы, связанные с возникновением долговременного следа памяти, наиболее выражены в областях синаптических контактов; именно в синаптических окончаниях зарегистрирована интенсификация синтеза специфических белков при обучении.
Следовательно, чем интенсивнее протекают в том или ином образовании мозга процессы переработкой запоминания поступающей информации, тем выше потребность в богатых энергией соединениях и субстратах для синтетических реакций.
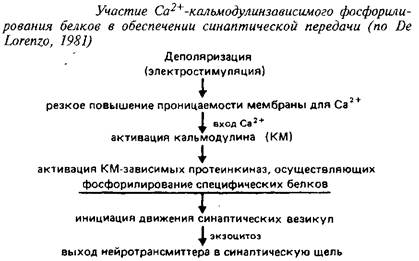
Кроме того, огромную роль в обеспечении функционирования синапсов играют процессы фосфорилирования белков, также связанные с потреблением АТФ или ГТФ. Циклы фосфорилирования – дефосфорилирования белков служат важным регуляторным механизмом, обеспечивающим пластичность на уровне нейронов. Процесс фосфорилирования является Са + -кальмодулинзависимым или циклонуклеотидзависимым; последнее обстоятельство делает понятным высокую t концентрацию циклических нуклеотидов и высокую активность ферментов их метаболизма в синаптических окончаниях. В синапсах процесс фосфофилирования специфических белков включается разнообразными нейромедиаторами. Изменение степени фосфорилирования белков, участвующих в функционировании синаптических окончаний, рецепторов, ферментов синтеза и метаболизма нейромедиаторов может существенно менять проводимость синапса; это доказано на примере фосфорилирования тирозингидроксилазы и ряда других белков.
Фосфорилирование специфических белков служит необходимым этапом синоптической передачи, обеспечивая выход некоторых нейромедиаторов в синаптическую щель. Энергозависимость процесса высвобождения нейромедиаторов в синаптическую щель, участие в нем АТФазных системна также энергозависимость процесса обратного захвата нейротрансмиттера из синаптической щели подтверждена результатами многих исследований. Последовательность протекающих; при этом процессов отражает схема 5.
Таким образом, осуществление такой важной и специфической функции нервной ткани, как синоптическая передача, требует значительных энергетических затрат. – В настоящее время трудно оценить количественно эти затраты, однако их большой объем не вызывает сомнений, поскольку число синоптических контактов на поверхности нейронов необычайно велико.
Еще одним важным обстоятельством, накладывающим определенный отпечаток на энергетический обмен и многие другие стороны метаболизма нервной ткани, является необычное для большинства других типов клеток соотношение между поверхностью клетки и объемом ее центральной части. Так, для мотонейронов коры кошки средние размеры тела клетки составляют около 50 мк, в то время как длина аксона – до 10 – 10 мк; общая поверхность клетки – до 10 мк. Поверхность дендритов клеток Пуркинье мозжечка в 80–100 раз превышает поверхность сомы.
Подобные особенности структуры клеток нервной системы объясняют причины значительных энергетических затрат на транспортные нужды клетки. В первую очередь необходимо упомянуть трансмембранный перенос субстратов, медиаторов, различных предшественников с помощью специфических транслоказ или в результате конформационных перестроек мембран, большинство из которых сопровождаются фосфорилированием-дефосфорилированием мембранах белков. Например, установлено, что не более 5% важнейшего энергетического субстрата – глюкозы – поступает в мозг за счет пассивной диффузии; подавляющее количество глюкозы переносится через ГЭБ с энергетическими затратами и при участии К + , Na – АТФазы. По мнению ряда нейрохимиков, на эти процессы расходуется в нейроне достигает 15 000–20 000, занимая до 38–40% поверхности нейрона.
В последние годы внимание нейрохимиков привлекает изучение аксонального и ретроградного транспорта. Показано, что по аксону переносятся различные белки, синтезированные в перикарионе, ряд аминокислот, некоторые углеводы – глюкозамины, сиаловые кислоты и др. Транспорт последних обусловливает специфические перестройки и функциональные модификации белков синаптических окончаний за счет включения в них углеводных компонентов. Представление о соединениях, транспортируемых по аксону с различной скоростью, дают сведения, приведенные в табл. 9.
Таблица 9. Характеристика аксонального транспорта
Гликопротеиды, гликолипиды, белки, медиаторы, ферменты, ионы Са +
Поддержание определенного конформационного состояния белков важных структурных образований нейрональных отростков – нейрофиламентов – также требует энергетических затрат, поскольку конформационные переходы белков нейрофиламентов осуществляются за счет реакций фосфорилирования-дефосфорилирования. Преимущественная локализация нейрофиламентов в осевом цилиндре аксонов и дендритов обеспечивает определенную пространственную ориентацию нейрональных отростков. Это обстоятельство имеет необычайную важность для осуществления нейрональных контактов, для организации функциональных ансамблей нейронов, т.е. для осуществления интегративной деятельности мозга.
Рассмотренные данные позволяют в некоторой степени конкретизировать общее положение об исключительно высоких энергетических потребностях нервной ткани и понять причину тесной зависимости между функциональной активностью мозга и интенсивностью энергетического обмена.
1. Для мозга характерна высокая интенсивность энергетического метаболизма; мозг взрослого животного или человека потребляет до 20–25% кислорода, поступающего в организм, и до 70% свободной глюкозы, выделяемой из печени в артериальную кровь.
2. Наиболее интенсивно потребление кислорода и глюкозы осуществляется в филогенетически более молодых отделах мозга; максимальная скорость дыхания обнаружена в коре больших полушарий, минимальная – в спинном мозге и периферических нервах. Интенсивность дыхания нейронов, как правило, выше, чем нейроглиальных клеток.
5. Энергетические расходы нервной системы
Сопоставив размеры мозга и размеры тела животных, легко установить закономерность, по которой увеличение размеров тела чётко коррелирует с увеличением размеров мозга (см. табл. 1; табл. 3). Однако мозг является только частью нервной системы и не может рассматриваться отдельно от спинного мозга и периферической нервной системы. Масса одного мозга очень условно отражает энергетические затраты организма. Баланс между затратами на работу и приобретёнными результатами составляет суть работы нервной системы и затрагивает краеугольные принципы организации мозга, которые определили эволюцию нервной системы и стратегий поведения.
Общий баланс энергетических затрат складывается из нескольких компонентов. Рассмотрим компоненты нервной системы, которые постоянно находятся в активном состоянии. К ним относятся все периферические отделы, поддерживающие тонус мускулатуры, контролирующие дыхание, пищеварение, кровообращение и т. д. Понятно, что отключение одной из таких систем приведёт к гибели организма. Нагрузка на эти системы постоянна, но не стабильна. Она меняется в зависимости от поведения. Если животное начинает питаться, то активность пищеварительной системы возрастает и расходы на содержание её нервного аппарата увеличиваются. Аналогично повышаются расходы на иннервацию и контроль за скелетной мускулатурой, если животное активно. Колебания этих энергозатрат относительно невелики, так как тонус мускулатуры или активность кишечника организм вынужден поддерживать и в состоянии покоя. Следовательно, работа периферических систем требует постоянных затрат, которые возрастают с повышением активности организма.
Совершенно иная ситуация складывается с энергетическими затратами головного мозга. Головной мозг всегда активен независимо от состояния организма. Это связано с фундаментальными свойствами нервной системы.
Таблица 3. Абсолютная (средняя) масса тела различных животных



Нейроны только тогда могут хранить информацию, когда могут её передавать. Поддержание как наследуемой (видоспецифической), так и приобрётенной памяти всегда крайне энергозатратно. Многие органы чувств работают по принципу модуляции постоянно проходящего сигнала, что тоже требует постоянных расходов.
Иначе говоря, существуют постоянные базовые затраты на поддержание активности головного мозга. Однако существует и значительная вариабельность потребления энергии мозгом в разных физиологических состояниях. Если животное находится в состоянии относительного покоя, то мозг потребляет минимальное количество энергии, которое расходуется на поддержание необходимых физиологических функций. Если животное активно добывает пищу, избегает опасности или размножается, то затраты организма на содержание мозга существенно увеличиваются.
Для первичноводных позвоночных характерны относительно небольшой головной мозг, но высокоразвитый спинной мозг и периферическая нервная система. У ланцетника головной мозг не имеет чёткой анатомической границы со спинным и идентифицируется только по топологическому положению и цитологическим особенностям строения. У других первичноводных — круглоротых, хрящевых, лопастепёрых, лучепёрых и костистых рыб — головной мозг небольшой по отношению к размерам тела (см. рис. I-7). В этих группах доминирует периферическая нервная система. Она, как правило, в несколько десятков, а то и в сотни раз больше головного и спинного мозга вместе взятого. Более того, если принять массу головного мозга за единицу, то масса спинного мозга будет
в среднем составлять от 1 до 30 таких единиц. Иначе говоря, масса спинного мозга обычно больше, чем головного, или равна ей. Следовательно, большую часть энергетических расходов в нервной системе первичноводных животных следует считать постоянными. Небольшой мозг даже в состоянии высокой активности не может существенно повлиять на изменение энергетических затрат. Примером могут служить акулы-няньки. При массе тела около 20 кг их головной мозг весит только 7–9 г, спинной мозг — 15–20 г, а вся периферическая нервная система по приблизительным оценкам весит около 250–300 г. Понятно, что головной мозг составляет только 3 % массы всей нервной системы. Допустим, что потребление энергии мозгом активного животного увеличилось в 3 раза. Это изменение всё равно будет ничтожно малым на фоне постоянной активности периферической нервной системы. Данный пример показывает, что большая доля энергетических затрат первичноводных животных приходится на периферическую нервную систему.
Таким образом, маленькие первичноводные животные с относительно большой нервной системой легко осуществляют мобилизацию организма при смене форм поведения. Избегание опасности, поиск добычи, преследование конкурирующей особи происходят в любой последовательности, прекращаются и начинаются почти мгновенно. Все, кто содержал аквариумных рыбок, много раз наблюдал подобную активность. Небольшому организму достаточно влияния нервной системы для быстрого изменения поведения. У крупных первичноводных животных головной мозг относительно мал и не может быстро мобилизовывать животное прямым действием нервной системы, тем более что поведение в основном детерминировано. Нервная система практически не может ничего добавить к врождённым формам поведения, поэтому её участие в изменении формы поведения сигнальное. Нервная система только сообщает организму о необходимости изменить форму поведения.
Следовательно, нужен простой и недорогой механизм запуска специфического поведения. С одной стороны, он должен быть зависимым от нервной системы и при этом влиять сразу на весь организм. С другой стороны, он должен продолжать действовать продолжительное время и оказывать вторичное влияние на саму нервную систему. Такой механизм запуска врождённых форм поведения сложился у животных ещё на заре появления хордовых. Это нейроэндокринная система с центральной регуляцией через головной мозг. Действительно, большая часть эндокринных желёз позвоночных контролируется гипофизом и гипоталамусом. Их активность регулирует гормональный статус животных и часто предопределяет выбор конкретной формы поведения. Для
первичноводных энергетические затраты на содержание небольшого мозга невелики.
Конкретное животное с его ограниченным персональным опытом имеет небольшую ценность в масштабах всего вида. Это связано с тем, что отсутствие способов накопления опыта, продолжительной памяти и механизмов
информационных контактов между особями нивелирует слабые следы индивидуальной адаптации.
У наземных позвоночных значение нервной системы существенно больше. Выход на сушу поставил бывших первичноводных в крайне сложную ситуацию. Органы чувств и мозг были прекрасно приспособлены
к работе в водной среде, но она мало походила на наземные условия обитания. Новые требования к нервной системе были продиктованы низким сопротивлением среды, увеличением массы собственного тела,
хорошим распространением запахов, звуков и электромагнитных волн. Эти новые условия стали стимулом для развития соответствующих отделов нервной системы. На границе сред сформировались весьма специфические
органы движения — конечности. Их функционирование невозможно без управления со стороны нервной системы. Изменились требования к вестибулярному аппарату. Если в воде упасть невозможно, то на
поверхности Земли такие неприятности неизбежны. Гравитационное поле предъявило крайне жёсткие требования к системе соматических рецепторов и к вестибулярному аппарату. Резкое повышение требований к
координации работы мускулатуры тела и конечностей привело к интенсивному развитию сенсомоторных отделов спинного, заднего и продолговатого мозга. Дыхание атмосферным воздухом, изменение водно-
солевого баланса и механизмов пищеварения обусловили развитие специфических систем контроля этих функций со стороны мозга и периферической нервной системы.
С появлением теплокровности требования к нервной системе стали возрастать. Любое повышение метаболизма приводит к увеличению потребления пищи и соответственно к её активному поиску. Совершенствование приёмов добывания пищи и постоянная экономия энергии — актуальные условия выживания животного с высоким метаболизмом. Для этого необходим мозг с развитой памятью и механизмами принятия быстрых и адекватных решений.
Таблица 4. Отношение массы головного мозга к массе спинного мозга

Активное регулируется более активным. Мозг должен работать с заметным опережением складывающейся ситуации, от этого зависят выживание и успех конкретного вида. Однако повышение метаболизма мозга приводит к неизбежному возрастанию затрат на его содержание. Возникает замкнутый круг: теплокровность требует усиления обмена, которое может быть достигнуто ещё большим повышением метаболизма нервной системы. Иначе говоря, успех теплокровных напрямую зависит от эффективности работы нервной системы. В реальной ситуации эта связь не столь прямолинейна, но общая тенденция сохраняется практически всегда.
Для теплокровных животных размер тела становится критичным. Небольшие животные вынуждены постоянно решать основную, но не единственную задачу — искать пищу. Мелкие насекомоядные съедают
ежедневно огромное количество пищи. Бурозубка ежедневно потребляет пищи в несколько раз больше массы собственного тела. В похожей ситуации находятся мелкие летучие мыши и птицы. У многих небольших животных возникли механизмы защиты организма от перерасхода энергии — торпидность. В этом состоянии колибри снижают метаболизм, частоту дыхания и температуру тела. На несколько часов животное впадает в своеобразную спячку, которая нужна для экономии энергии. Иначе говоря, мелкие теплокровные могут находиться в двух основных состояниях: гиперактивности либо спячки. Промежуточное состояние малоэффективно, поскольку энергетические расходы не компенсируются поступающей пищей. Даже в случае её избытка промежуточное состояние достигает только долей процента основных форм поведения.
Мозг таких небольших насекомоядных, как американский крот- скалёпус (Scalopus aquaticus) или короткохвостая бурозубка (Blarina brevicauda), имеет массу 1/34 и 1/50 массы всего тела. В этих же пределах колеблется отношение масса мозга/масса тела у луговой полёвки (Arvicola argestis) — 1/47, гудзонской белки (Sciurus hudsonicus) — 1/36, обычной домовой мыши (Mus musculus) — 1/50 и у мыши Вагнера (Mus wahner!) 1/45. Спинной мозг у этих животных обычно составляет 1/4-1/6 массы мозга. Периферическая нервная система мелких млекопитающих, как уже
отмечалось, может составить примерно 2/3 массы головного и спинного мозга. Это позволяет довольно приблизительно определить реальные общие размеры нервной системы у грызунов и насекомоядных. По- видимому, их нервная система составляет 1/17-1/25 массы всего тела. Это означает, что она потребляет очень большую часть всей энергии организма.
Следовательно, нужен простой и недорогой механизм запуска специфического поведения. С одной стороны, он должен быть зависимым от нервной системы и при этом влиять сразу на весь организм. С другой стороны, он должен продолжать действовать продолжительное время и оказывать вторичное влияние на саму нервную систему. Такой механизм запуска врождённых форм поведения сложился у животных ещё на заре появления хордовых. Это нейроэндокринная система с центральной регуляцией через головной мозг. Действительно, большая часть эндокринных желёз позвоночных контролируется гипофизом и гипоталамусом.
У наземных позвоночных значение нервной системы существенно больше. Выход на сушу поставил бывших первичноводных в крайне сложную ситуацию. Органы чувств и мозг были прекрасно приспособлены к работе в водной среде, но она мало походила на наземные условия обитания. Новые требования к нервной системе были продиктованы низким сопротивлением среды, увеличением массы собственного тела, хорошим распространением запахов, звуков и электромагнитных волн. Эти новые условия стали стимулом для развития соответствующих отделов нервной системы. На границе сред сформировались весьма специфические органы движения — конечности. Их функционирование невозможно без управления со стороны нервной системы. Изменились требования к вестибулярному аппарату. Если в воде упасть невозможно, то на поверхности Земли такие неприятности неизбежны. Гравитационное поле предъявило крайне жёсткие требования к системе соматических рецепторов и к вестибулярному аппарату. Резкое повышение требований к координации работы мускулатуры тела и конечностей привело к интенсивному развитию сенсомоторных отделов спинного, заднего и продолговатого мозга. Дыхание атмосферным воздухом, изменение водно- солевого баланса и механизмов пищеварения обусловили развитие специфических систем контроля этих функций со стороны мозга и периферической нервной системы.
Следовательно, роль мозга после выхода на сушу существенно возросла. Эти изменения сказались на массе центральной и периферической нервной системы. У наземных животных её масса всегда больше, чем у первичноводных обитателей идентичных размеров. Увеличение массы нервной системы относительно массы всего тела привело к нескольким последствиям.
Во-первых, возросла общая масса периферической нервной системы. Это увеличение было весьма заметным. Основной прирост пришёлся на иннервацию конечностей, кожную чувствительность, органы дыхания и черепно-мозговые нервы. Такое масштабное увеличение сенсорных входов, по-видимому, привело к многократному росту относительных размеров периферической иннервации. Вполне естественно, что в определённый момент истории произошло смещение метаболического баланса нервной системы в сторону периферических отделов.
Во-вторых, произошло увеличение размеров спинного мозга. Поскольку он выполняет функции интегративного центра периферической нервной системы, его преобладание над головным мозгом было неизбежным следствием выхода животного на сушу. На морфологии спинного мозга особенно сказалось развитие конечностей. Сформировались специальные спинномозговые утолщения и даже специализированные центры управления в заднем и продолговатом мозге. У крупных динозавров эти отделы превысили размеры головного мозга, хотя и выполняли примитивные функции управления работой мышц. Даже если сравнить современных амфибий и рептилий, то станет понятной общая тенденция изменения размеров спинного и головного мозга при освоении сложной системы наземных движений. У геккона-токо головной мозг по массе равен мозгу бурой лягушки и составляет 0,27 г при сходных размерах тела. Однако спинной мозг бурой лягушки имеет массу 0,15 г, а геккона — уже 0,37 г.
В-третьих, увеличился и сам головной мозг. Его рост был вызван увеличением мозгового представительства анализаторов различных типов. В первую очередь это были моторные, сенсомоторные, зрительные, слуховые и обонятельные центры. Дальнейшее развитие получила система связей между различными отделами мозга. Они стали основой для быстрого сравнения активностей специализированных анализаторов. Параллельно развились внутренний рецепторный комплекс и сложный эффекторный аппарат. Синхронизированное управление дистантными рецепторами, сложной мускулатурой и внутренними органами невозможно без ассоциативных центров. Они возникали в процессе эволюции неоднократно и на базе различных отделов головного мозга.
Читайте также: