
Нервных волокнах скорость проведения нервного импульса выше
Потенциалы действия (импульсы возбуждения) обладают способностью распространяться вдоль по нервным и мышечным волокнам. Потенциалы могут быть локальными, они распространяются на небольшие расстояния 1-2 мм с затуханием (декрементом) и импульсными. Импульсные потенциалы распространяются без декремента на значительные расстояния – до нескольких десятков сантиметров. Локальные потенциалы возникают в ответ на действие подпорогового раздражителя, например, на мембране рецепторной клетки. Если локальное возбуждение попадает в участок мембраны, способной генерировать ПД, и амплитуда локального тока достигает критического уровня деполяризации, формируется ПД, который распространяется по всей длине нервного волокна.
Передача информации на большие расстояния в пределах нервной системы осуществляется с помощью нервных импульсов по аксонам нейронов. Обязательным условием проведения нервного импульса является наличие на всем протяжении или в ограниченных, но повторяющихся участках волокна потенциал чувствительных ионных каналов. В зависимости от расположения и концентрации ионных каналов в мембране волокна выделяют два способа проведения нервного импульса.
1. Непрерывное проведение нервного импульсаосуществляется в безмиелиновых волокнах, объясняется равномерным распределением потенциал чувствительных ионных каналов, участвующих в генерации ПД.
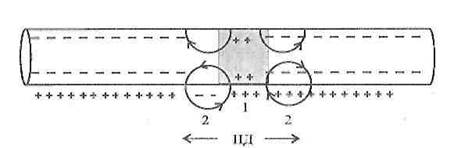
Рисунок 3 - Непрерывное распространение ПД в нервном волокне
Возникший ПД обеспечивает открытие потенциал зависимых Na-каналов на соседнем участке мембраны нервного волокна и движение ионов Na + внутрь волокна, что обеспечивает развитие критического уровня деполяризации на соседнем участке нервного волокна и возникновение нового ПД. Непрерывное распространение нервного импульса идет через генерацию новых импульсов по эстафете, когда каждый возникший импульс является раздражителем для соседнего участка нервного волокна и обеспечивает возникновение нового ПД.
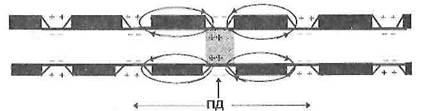
Рисунок 4 – Сальтаторное распространение ПД в нервных волокнах
2. Сальтаторное проведение нервного импульса (ПД) осуществляется в миелиновых волокнах, так как у них потенциал чувствительные ионные каналы локализованы только в участках мембраны перехватов Ранвье, где их плотность достигает 12 000 на 1 мкм 2 . В области межузловых сегментов, обладающих высокими изолирующими свойствами, потенциал чувствительных каналов нет, вследствие чего мембрана осевого цилиндра там практически невозбудима. Поэтому ПД, возникший в одном перехвате Ранвье распространяется через межузловой сегмент до соседнего перехвата, деполяризует мембрану до критического уровня и вызывает возникновение потенциала действия.
Сальтаторное проведение нервных импульсов является эволюционно более поздним механизмом, возникшим впервые у позвоночных в связи с миелинизацией нервных волокон. Оно имеет два важных преимущества по сравнению с непрерывным механизмом проведения возбуждения:
-более экономично по затрате энергии, так как возбуждаются только перехваты Ранвье, площадь которых составляет менее 1% от площади мембраны волокна, следовательно, требуется меньше энергии для восстановления трансмембранных градиентов ионов натрия и калия;
-возбуждение проводится с большей скоростью (до 120 м/с), чем в безмиелиновых волокнах (0,5-2,0 м/с).
В связи с этими приемуществами миелиновые волокна в нервной системе сформировались там, где необходима наиболее быстрая регуляция функций. Все особенности распространения возбуждения в ЦНС объясняются ее нейронным строением: наличием химических синапсов, многократным ветвлением аксонов нейронов, наличием замкнутых нейронных путей.
Проведение нервных импульсов по волокнам нейронов починяется определенным законам:
Закона 1: нервный импульс распространяется в обе стороны от места раздражения.
Закон 2: проведение нервного импульса по волокну происходит изолировано и не распространяется на параллельные волокна. Объяснение этого закона заключается в том, что аксолемма имеет очень высокое сопротивление и не пропускает петли тока на невозбужденные волокна, расположенные рядом. Изолированное проведение обеспечивает высокую точность регуляторной деятельности ЦНС.
Закон 3: скорость проведения возбуждения по нервному волокну определяется его диаметром. Отсюда следствие: чем толще нервное волокно, тем больше скорость проведения нервного импульса по этому волокну.
Закон 4: нерв сохраняет способность к проведению возбуждения в течение 6-8 часов непрерывного раздражения (закон Н.Е. Введенского, 1883).
Закон 5: действие веществ, блокирующих работу ионных каналов, без нарушения целостности нервного волокна вызывает состояние обратимого парабиоза, (закон парабиоза, Введенский, 1901).
Кроме того, выделяют несколько видов распространения возбуждения:
1. Иррадиация (дивергенция) возбуждения в ЦНС, которая объясняется ветвлением аксонов и наличием вставочных нейронов, аксоны которых также ветвятся. Дивергенция расширяет сферу действия каждого нейрона. Один нейрон, посылая импульсы в кору большого мозга, может участвовать в возбуждении до 5000 нейронов.
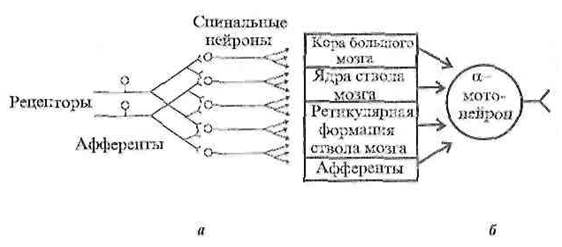
Рисунок 4 – Дивергенция афферентных дорсальных корешков на нейроны головного мозга (а – дивергенция, б – конвергенция)
2. Конвергенция возбуждения представляет собой схождение нескольких нервных импульсов, идущих по разным путям к одному и тому же нейрону. Явление конвергенции распространения возбуждения описал Э. Шеррингтон, поэтому явление было названо принцип шеррингтоновской воронки или принцип общего конечного пути. Примером может служить конвергенция возбуждения на спинальном мотонейроне. Мотонейрон, иннервирующий мышцы глотки, участвует в рефлексах глотания, кашля, сосания, чиханья и дыхания, образуя общий конечный путь для многочисленных рефлекторных дуг.
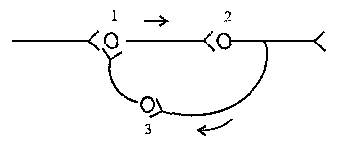
Рисунок 5 – Циркуляция возбуждения в замкнутых нейронных цепях
3. Циркуляция возбуждения по замкнутым нейронным цепям, возникает в результате замыкания группы нейронов в кольцевую структуру. Циркуляция возбуждения – одна из причин явления последействия. Считают, что циркуляция возбуждения в замкнутых нейронных цепях наиболее вероятный механизм феномена кратковременной памяти. Циркуляция возбуждения возможна в цепи нейронов и в пределах одного нейрона в результате контактов разветвлений его аксона с собственными дендритами и телом.
Литература
1. Смирнов, В.М. Физиология сенсорных систем и высшая нервная деятельность / В.М. Смирнов, С.М. Будылина. - М.: Медицина, 2003. - 304 с.
2. Шульговский, В. В. Основы нейрофизиологии: Учебное пособие для студентов вузов. - М.: Аспект Пресс, 2000. - с. 277.
3. Батуев, А.С. Физиология поведения. Нейрофизиологические закономерности / А.С. Батуев. - Л.: Наука, 1986. - 340 с.
4. Александров, Ю.И. Психофизиология / Ю.И.Александров. - М.: Медицина, 2001. - 230 с.
Данилова, Н.Н. Физиология высшей нервной деятельности / Н. Н. Данилова, А.Л. Крылова. - Ростов н/Д: Феникс, 1999. – 480 с.
[1] ионоселективные каналы и ионные насосы
[2] правило действует и в обратном направлении
[3] 1780 год, профессор анатомии Болонского университета
Нервный импульс (лат. nervus нерв; лат. impulsus удар, толчок) — волна возбуждения, распространяющаяся по нервному волокну; единица распространяющегося возбуждения.
Нервный импульс обеспечивает передачу информации от рецепторов к нервным центрам и от них к исполнительным органам — скелетной мускулатуре, гладким мышцам внутренних органов и сосудов, железам внутренней и внешней секреции и т. д.
Распространение Нервных импульсов отождествляется с проведением потенциалов действия (см. Биоэлектрические потенциалы). Возникновение возбуждения может быть результатом раздражения (см.), напр, воздействие света на зрительный рецептор, звука на слуховой рецептор, или процессов, протекающих в тканях (спонтанное возникновение Н. и.). В этих случаях Н. и. обеспечивают согласованную работу органов при протекании какого-либо физиологического процесса (напр., в процессе дыхания Н. и. вызывают сокращение скелетных мышц и диафрагмы, результатом чего являются вдох и выдох, и т. д.).
В живых организмах передача информации может осуществляться и гуморальным путем, посредством выброса в русло крови гормонов, медиаторов и т. п. Однако преимущество информации, передаваемой при помощи Н. и., состоит в том, что она более целенаправленна, передается быстро и может быть точнее закодирована, чем сигналы, посылаемые гуморальной системой.
Факт, что нервные стволы являются путем, по к-рому передаются влияния от мозга к мышцам и в обратном направлении, был известен еще в эпоху античности. В средние века и вплоть до середины 17 в. считалось, что по нервам распространяется некая субстанция, подобная жидкости или пламени. Идея о электрической природе Н. и. возникла в 18 в. Первые исследования электрических явлений в живых тканях, связанных с возникновением и распространением возбуждения, были осуществлены Л. Гальвани. Г. Гельмгольц показал, что скорость распространения Н. и., к-рую ранее считали близкой к скорости света, имеет конечное значение и может быть точно измерена. Германн (L. Hermann) ввел в физиологию понятие потенциала действия. Объяснение механизма возникновения и проведения возбуждения стало возможным после создания С. Аррениусом теории электролитической диссоциации. В соответствии с этой теорией Бернштейн (J. Bernstein) предположил, что возникновение и проведение Н. и. обусловлено перемещением ионов между нервным волокном и окружающей средой. Англ. исследователи А. Ходжкин, Б. Катц и Э. Хаксли детально исследовали трансмембранные ионные токи, лежащие в основе развития потенциала действия. Позже стали интенсивно изучаться механизмы работы ионных каналов, по к-рым происходит обмен ионами между аксоном и окружающей средой, и механизмы, обеспечивающие способность нервных волокон проводить ряды Н. и. разного ритма и продолжительности.
Н. и. распространяется за счет местных токов, возникающих между возбужденным и невозбужденным участками нервного волокна. Ток, выходящий из волокна наружу в покоящемся участке, служит раздражителем. Наступающая после возбуждения в данном участке нервного волокна рефрактерность обусловливает поступательное движение Н. и.
Количественно соотношения разных фаз развития потенциала действия можно охарактеризовать, сопоставляя их по амплитуде и длительности во времени. Так, напр., для миелиновых нервных волокон группы А млекопитающих диаметр волокна находится в пределах 1—22 мк, скорость проведения — 5—120 м/сек, длительность и амплитуда высоковольтной части (пика, или спайка) — 0,4—0,5 мсек и 100—120 мв соответственно, следовой негативный потенциал — 12—20 мсек (3—5% от амплитуды спайка), следовой позитивный потенциал — 40—60 мсек (0,2% от амплитуды спайка).
Возможности передачи разнообразной информации расширяются за счет повышения скорости развития потенциала действия, скорости распространения, а также за счет повышения лабильности (см.) — т. е. способности возбудимого образования воспроизводить в единицу времени высокие ритмы возбуждения.
Возникновение Н. и. в нервных клетках (см.) или рецепторах (см.) связано с деполяризацией мембраны, т. е. со снижением величины электрического потенциала на мембране (потенциала покоя, или мембранного потенциала). Если величина мембранного потенциала снижается на 10—20% (пороговый критический уровень), то местный процесс переходит в распространяющийся — возникает потенциал действия (см. Возбуждение).
Конкретные особенности распространения Н. и. связаны со строением нервных волокон (см.). Сердцевина волокна (аксоплазма) обладает низким сопротивлением и, соответственно, хорошей проводимостью, а окружающая аксоплазму плазматическая мембрана — большим сопротивлением. Особенно велико электрическое сопротивление наружного слоя у миелинизированных волокон, у к-рых свободны от толстой миелиновой оболочки только перехваты Ранвье. В безмиелиновых волокнах Н. и. движется непрерывно, а в миелиновых — скачкообразно (сальтаторное проведение).
Различают декрементное и бездекрементное распространение волны возбуждения. Декрементное проведение, т. е. проведение возбуждения с угасанием, наблюдается в безмиелиновых волокнах. В таких волокнах скорость проведения Н. и. невелика и по мере отдаления от места раздражения раздражающее действие местных токов постепенно уменьшается вплоть до полного угасания. Декрементное проведение свойственно волокнам, иннервирующим внутренние органы, обладающие низкой функц, подвижностью. Без декрементное проведение характерно для миелиновых и тех безмиелиновых волокон, к-рые передают сигналы к органам, обладающим высокой реактивностью (напр., сердечной мышце). При бездекрементном проведении Н. и. проходит весь путь от места раздражения до места реализации информации без затухания.
Передача Н. и. с нервного волокна на мышечное или какой-либо другой эффектор осуществляется через синапсы (см.). У позвоночных животных в подавляющем большинстве случаев передача возбуждения на эффектор происходит при помощи выделения ацетилхолина (нервно-мышечные синапсы скелетной мускулатуры, синаптические соединения в сердце и др.). Для таких синапсов характерно строго одностороннее проведение импульса и наличие временной задержки передачи возбуждения.
В синапсах, в синаптической щели которых сопротивление электрическому току благодаря большой площади контактирующих поверхностей мало, происходит электрическая передача возбуждения. В них нет синаптической задержки проведения и возможно двустороннее проведение. Такие синапсы свойственны беспозвоночным животным.
Регистрация Н. и. нашла широкое применение в биол, исследованиях и клин, практике. Для регистрации используют шлейфные и чаще катодные осциллографы (см. Осциллография). При помощи микроэлектродной техники (см. Микроэлектродный метод исследования) регистрируют Н. и. в одиночных возбудимых образованиях — нейронах и аксонах. Возможности исследования механизма возникновения и распространения Н. и. значительно расширились после разработки метода фиксации потенциала. Этим методом были получены основные данные о ионных токах (см. Биоэлектрические потенциалы).
Нарушение проведения Н. и. происходит при повреждении нервных стволов, напр, при механических травмах, сдавливании в результате разрастания опухоли или при воспалительных процессах. Такие нарушения проведения Н. и. зачастую бывают необратимы. Следствием прекращения иннервации могут быть тяжелые функциональные и трофические расстройства (напр., атрофия скелетных мышц конечностей после прекращения поступления Н. и. вследствие необратимой травмы нервного ствола). Обратимое прекращение проведения Н. и. может быть вызвано специально, в терапевтических целях. Напр., с помощью анестезирующих средств блокируют импульсацию, идущую от болевых рецепторов в ц. н. с. Обратимое прекращение проведения Н. и. вызывает и новокаиновая блокада. Временное прекращение передачи Н. и. по нервным проводникам наблюдается и во время общего наркоза.
Библиография: Бpеже М. А. Электрическая активность нервной системы, пер. с англ., М., 1979; Жуков Е. К. Очерки по нервно-мышечной физиологии, Л., 1969; Коннели К. Восстановительные процессы и обмен веществ в нерве, в кн.: Совр, пробл. биофизики, пер. с англ., под ред. Г. М. Франка и А. Г. Пасынского, т. 2, с. 211, М., 1961; Костюк П. Г. Физиология центральной нервной системы, Киев, 1977; Латманизова Л. В. Очерк физиологии возбуждения, М., 1972; Общая физиология нервной системы, под ред. П. Г. Костюка, Л., 1979; Тасаки И. Нервное возбуждение, пер. с англ., М., 1971; Ходжкин А. Нервный импульс, пер. с англ., М., 1965; Ходоров Б. И. Общая физиология возбудимых мембран, М., 1975.
Здоровье твоей планеты в твоих руках!
Скорость распространения нервных импульсов
В 1830 г. один из крупнейших физиологов XIX века Иоганн Мюллер заявил, что скорость распространения ПД измерить невозможно. По его мнению, поскольку ПД – это электрический импульс, он должен проводиться со скоростью, примерно равной скорости света (3–1010 см/с); учитывая небольшие размеры биологических объектов, даже с помощью лучших инструментов того времени измерить такую скорость было невозможно.
Спустя 15 лет один из студентов Мюллера Герман фон Гельмгольц с помощью простого и изящного эксперимента, который легко воспроизвести на студенческом лабораторном практикуме (рис. 6–8), измерил скорость распространения импульсов в нерве лягушки. В своих опытах Гельмгольц раздражал нерв в двух участках, отстоящих друг от друга на 3 см, и измерял время от момента подачи стимула до максимума мышечного сокращения. Предположим, что при раздражении дистального (расположенного ближе к мышцам) участка это время уменьшается на 1 мс. Тогда скорость распространения импульсов V равна
Эта величина оказалась на семь порядков меньше, чем скорость распространения электрического тока в медном проводнике или в растворе электролита. Отсюда Гельмгольц сделал совершенно правильный вывод, что проведение нервного импульса –это более сложный процесс, чем простое продольное распространение тока в нервном волокне.
Скорость распространения импульсов в различных аксонах варьирует от 120 м/с (в некоторых крупных волокнах) до нескольких сантиметров в секунду (в очень тонких аксонах). Эти различия между скоростью проведения в разных волокнах иллюстрируют табл. 6–1 и рис. 6–9.
Скорость распространения импульса в значительной степени зависит от того, как быстро участок мембраны, расположенный на определенном расстоянии от места подачи стимула, деполяризуется местными токами до порогового уровня. Чем выше постоянная длины волокна, тем дальше могут распространяться эти токи, тем быстрее происходит деполяризация мембраны впереди от места возбуждения и, следовательно, тем выше скорость распространения импульса. Влияние постоянной длины на эту скорость можно продемонстрировать, если поместить аксон в масло или в воздух. При этом на поверхности аксона остается лишь тонкая пленка солевого раствора, и постоянная длины уменьшается из–за увеличения наружного продольного сопротивления [в уравнении (6–2) –r0]. В этих условиях скорость проведения возбуждения будет ниже чем при погружении аксона в солевой раствор.
Таблица 6–1. Классификация нервных волокон лягушки по их диаметру и скорости проведения возбуждения (Erlanger, Gasser, 1937)
В процессе эволюции живые организмы выработали два способа увеличения постоянной длины аксона и тем самым–скорости распространения импульса. Один из них (типичным примером могут быть гигантские аксоны кальмаров, членистоногих кольчатых червей, костистых рыб) – это увеличение диаметра аксона, т. е. уменьшение внутреннего продольного сопротивления [в уравнении (6–2) – ri] Подробнее этот вопрос рассмотрен в дополнении 6–2. Гигантские аксоны развились в процессе эволюции у некоторых видов животных для того, чтобы обеспечивать быструю синхронную активацию двигательных рефлексов, например движений мантии у кальмара и рефлекса отдергивания либо избегания у некоторых членистоногих (раков, тараканов) и кольчатых червей (например, земляных).
Сальтаторное проведение
Второй способ увеличить скорость проведения нервных импульсов, реализовавшийся только у позвоночных животных, состоит в изоляции участков аксона с помощью миелиновой оболочки. При этом постоянная длины соответствующих участков значительно увеличивается, и тем самым существенно облегчается проведение тока в продольном направлении. По мере развития животного миелин откладывается вокруг периферических и центральных аксонов глиальными клетками, расположенными вблизи этих аксонов. В результате вокруг волокон образуется плотная многослойная оболочка из клеточных мембран. К клеткам, синтезирующим миелин, относятся шванновские клетки (в области периферических нервов) и олигодендроциты (рис. 6–10) (в ЦНС). На поперечных срезах миелиновой оболочки видны периодически повторяющиеся промежутки в 12 нм, образующиеся в результате наслоения мембран глиальных клеток. С образованием каждого нового слоя поперечное сопротивление оболочки увеличивается. Поскольку слоев в этой оболочке много, ее емкость гораздо ниже, чем у одиночной мембраны. Многослойная миелиновая оболочка периодически прерывается (так называемые перехваты Ранвье>, и на этих небольших участках возбудимая мембрана аксона контактирует с внеклеточной средой. Между перехватами Ранвье миелиновая оболочка тесно прилегает к мембране аксона, практически вытесняя внеклеточную среду. Кроме того, участки мембраны аксона между перехватами Ранвье, по–видимому, не содержат натриевых каналов.
Благодаря изолирующим свойствам миелиновой оболочки постоянная длины аксона резко возрастает: наличие этой оболочки оказывает такой же эффект, как и увеличение rм [уравнение (6–2)]. Из–за высокого сопротивления миелиновой оболочки местные токи, текущие впереди от волны возбуждения, выходят из аксона почти исключительно в области перехватов Ранвье. Кроме того, поскольку емкость толстой миелиновой оболочки мала, на перезарядку этой емкости в участках между перехватами расходуется лишь очень небольшой ток. Благодаря этим особенностям ПД, возникающий в каком–либо перехвате, электротонически деполяризует лишь мембрану, расположенную в области следующего перехвата, и поэтому импульсы в таких аксонах не распространяются по всей их длине, как в немиелинизированных нервных волокнах (например, в аксоне кальмара). Они возникают лишь в небольших участках мембраны –перехватах Ранвье. Все это обусловливает сальтаторное (скачкообразное) проведение, при котором импульсы распространяются прерывисто от перехвата к перехвату (рис. 6–11). Скорость распространения. ПД при этом резко увеличивается, поскольку электротоническое проведение местных токов между перехватами осуществляется очень быстро. Таким образом, у позвоночных животных Природа решила проблему быстрого распространения нервных импульсов, не прибегая к созданию таких громоздких структур, как гигантские аксоны.
Не́рвный и́мпульс, распространение по нервным волокнам возбуждения (биоэлектрического импульса) в ответ на раздражение нейронов.
Во второй половине 19 века в работах Г. Гельмгольца и Э.
Геринга на нерве лягушки было показано, что биоэлектрический сигнал (ток, или потенциал действия), в отличие от электрического тока в обычном проводнике, распространяется по нервному волокну с конечной скоростью (3-120 м/сек).
Возможность распространения нервных импульсов по нервным волокнам определяется их строением, напоминающим строение электрического кабеля, где роль проводника играют аксоны, а роль изолятора — миелиновая оболочка аксона, представляющая собой мембрану шванновской клетки, намотанную на аксон в несколько слоев.
Скорость распространения нервного импульса по толстым миелинизированным волокнам (диаметром 10-20 микрон) у человека достигает 70-120 м/сек, а по самым тонким немиелинизированным нервным волокнам — на два порядка ниже (менее 2 м/сек).
Способность вырабатывать нервные импульсы — одно из основополагающих свойств нейронов.
Нервные импульсы обеспечивают быстрое проведение однотипных сигналов (потенциалов действия) по аксонам на большие расстояния и поэтому являются важнейшим средством обмена информацией как между нервными клетками, так и между нервными и другими типами клеток. Информация о силе раздражения нервной клетки кодируется и передается другим клеткам путем изменения частоты следования нервных импульсов.
Нервный импульс. М., 1965.
Скорость распространения нервных импульсов может быть различной: меньше 1 метра в секунду в очень тонких аксонах и около 100 метров в секунду в толстых аксонах <например, в аксонах, иннервирующих мышцы).
Дейлом и О. Лёви.
Как известно, центральная нервная система человека (включая, конечно, не только головной мозг, но и спинной) состоит примерно из 10 миллиардов (1010) нервных клеток. Почти все нервные клетки, за редким исключением, получают информацию непосредственно в форме импульсов (смотрите рисунок ниже) сразу от нескольких нервных клеток (нередко от сотен их) и передают ее столь же большому числу клеток.
Возбуждение и торможение нервной клетки

Возбуждение и торможение нервной клетки осуществляют нервные волокна, образующие синапсы на ее поверхности.
Вверху (1) двигательный нейрон в состоянии покоя. Импульсы, приходящие по одному возбуждающему волокну (2), еще не в состоянии вызвать разряд двигательного нейрона. Разряд возникает только тогда, когда импульсы приходят и по второму возбуждающему волокну (3) (пороговое состояние нейрона). Если нейрон получает еще и импульсы по тормозному волокну, то он возвращается в подпороговое состояние (4).
Внизу (б) — импульсы приходят только по тормозному волокну. Электрические импульсы, распространяющиеся по возбуждающим и тормозным нервным волокнам, не отличаются друг от друга. Их противоположное действие объясняется выделением в синаптических окончаниях разных химических передатчиков.
В данной нервной клетке в зависимости от ее порога возбуждения может возникнуть разряд импульсов при раздражении всего лишь нескольких приходящих к ней волокон; в других же случаях разряд импульсов не возникает даже при раздражении многих таких волокон.
Давно известно, что различные факторы способны повышать или понижать порог возбуждения нервной клетки.
Более того, примерно 60 лет назад было высказано предположение, что некоторые волокна должны тормозить разряд импульсов в клетке, к которой они подходят, а не возбуждать ее. Предположение это впоследствии подтвердилось, и в настоящее время механизм торможения выяснен. Двум равноценным процессам — торможению и его антиподу — возбуждению нервной клетки — и посвящена данная статья.
Читайте далее:
Распространение нервного импульса по нервному волокну
Особенность нервного импульса (потенциала действия) является его самораспространение по нервному или мышечному волокну, в результате которого обеспечивается передача информации от периферических рецепторных окончаний к нервным центрам, а от них к эффекторам.
В мышечных клетках нервный импульс оказывает пусковое влияние на процессы, активизирующие сократительный аппарат. Распространение неровного импульса начинается с момента, когда внутренняя часть нервного волокна заряжается положительно, и разность потенциалов между внутренней средой нерва и наружной может достигать 40-50 мВ.
Проведение нервного импульса можно сравнить с распространением пламени по бикфордовому шнуру: импульс возникает при пороговом запале, идет с определенной скоростью без затухания, передаваясь от возбужденного участка к соседнему – невозбужденному.
В основе объяснения этого механизма лежит теория немецкого физика Л.
Германа (1879) и затем А.
Ходжкина (1937). Согласно этой теории возникший в точке раздражения потенциал действия является источником раздражения соседнего невозбужденного участка волокна. Это происходит вследствие возникновения круговых, или местных токов между возбужденным (т.е. отрицательно заряженным) и соседним (положительно заряженным) участками мембраны.
РИС
В результате местного электро-химического сдвига ионной проницаемости мембраны возникает ее деполяризация и достигается критический пороговый потенциал действия. В зоне первоначально возбужденной в это время восстанавливается потенциал покоя. Затем потенциал действия возникает на следующем участке волокна и т.д. Поэтому волна возбуждения проходит вдоль волокна, не затухая и не поворачивая назад, ибо на соседнем пройденном участке находится рефрактерная зона.
Важное свойство возбудимых тканей рефрактерность. Она определяет прерывистости импульсов. В естественных условиях по нервам непрерывно бегут нервные импульсы. Частота этих ритмических зарядов зависит от силы раздражения. Двигательные нейроны могут проводить около 500 импульсов в секунду, промежуточные – 1000.
Таким образом, распространение (проведение) возбуждения заключается в последовательном возникновении и исчезновении потенциала действия на протяжении нервного или мышечного волокна.
Непрерывное проведение импульсов характерно для мышечного волокна и безмякотных, безмиелиновых нервных волокон, имеющих только шванновскую оболочку.
Таких волокон в нервной системе меньшинство. В мякотных нервных волокнах, имеющих миелиновую оболочку (она является хорошим изолятором) круговые токи могут возникать лишь между двумя соседними (возбужденными и невозбужденными) перехватами Ранвье, где миелин отсутствует. Следовательно, возбуждение в миелинизированных нервных волокнах распространяется скачкообразно, сальтаторно (la salto – скачу, прыгаю).
Скорость проведения импульса в нервных волокнах определяется их гистостроением и диаметром. В миелинизированных нервных волокнах она составляет 30-120 м/сек (6 км/мин, 360 км/ч), в безмякотных — 0,5-3 м/сек, в скелетных 5 м/сек. (рис **)
РИС
Особенности проведении нервного импульса:
— возбуждение проводится в обе стороны по нервному волокну от места раздражения;
— проведение возбуждения возможно лишь при целостности волокна;
— более толстые волокна обладают наиболее низким порогом возбуждения;
— волокна, входящие в состав одиночных или смешанных нервов проводят возбуждение изолированно, т.е.
не переходя на другие волокна и адресуются лишь своим клеткам;
Проявлением возбуждения нервных волокон является повышение обменных процессов. Возрастает расход кислорода и выделение СО2 , увеличивается расход АТФ, повышается образование молочной кислоты. В окончаниях аксонов нервных клеток при возбуждении секретируются особые химические вещества – медиаторы, оказывающие возбуждающее или тормозящее влияние на иннервируемые ткани.
Дата добавления: 2016-06-29; просмотров: 2199;
Читайте также: