
Нервный импульс может распространяться только в
Потенциал действия (нервный импульс): распространение
Нервный импульс в основном распространяется только в одном направлении - по нескольким дендритам к телу клетки и от тела по единственному аксону к мышце, органу или дендриту следующего нейрона .
Смысл потенциала действия (нервного импульса) заключается в его распространении. Как это драматическое изменение мембранной полярности распространяется? И снова ответ заключается в локальных токах. Мы уже видели, что деполяризация небольшого участка мембраны ведет к возникновению малых электрических токов, распространяющихся отсюда и деполяризующих следующие участки мембраны на небольшом расстоянии. То же самое происходит и, когда мембрана резко меняет полярность при прохождении потенциала действия, описанного выше. Локальные токи, деполяризуя соседний участок мембраны, так же открывают в нем Na+-каналы и запускают последовательность событий, ответственную за формирование потенциала действия, описанную выше. Можно сказать, что потенциал действия похож на огонь, бегущий по дорожке из пороха. Каждый активированный участок запускает последующий, оставляя за собой дорожку из пепла. В случае нервного волокна инактивация, символизируемая пеплом, представляет собой гиперполяризацию мембраны вслед за прошедшим потенциалом действия ( рис. 2.9 ) и, что более существенно, то, что Na+-воротный механизм остается в закрытом состоянии в течение прибл. 1,5 мс после их потенциал-зависимого открывания. Это состояние мембраны именуется рефрактерностью . Рефрактерный период длительностью 1,5 - 2 мс имеет существенное значение для сенсорной сигнализации. Он ограничивает частоту, с которой импульсы могут следовать по сенсорному нервному волокну. Такая длительность означает, что сенсорное волокно в состоянии проводить импульсы с частотой не выше 500 Гц. Это важно, например, в детектировании частоты звука и частотной дискриминации в слуховой системе (см. гл. РАВНОВЕСИЕ И СЛУХ ). В то время, как рефрактерный период ограничивает частоту, с которой импульсы могут проводиться по нервному волокну, диаметр последнего (при прочих равных) определяет скорость проведения ( табл. 2.2 ). Чем больше диаметр, тем дальше по аксоплазме распространяются локальные токи и на большем расстоянии они могут открыть Na+-воротный механизм. Поэтому у многих беспозвоночных, например, кольчатых червей и головоногих моллюсков, развились гигантские волокна, обеспечивающие быстрое проведение, необходимое для реакции избегания. Позвоночные развили иной механизм увеличения скорости проведения - миелинизацию . Миелин состоит из нескольких слоев мембран шванновских клеток, обвивающихся вокруг аксон, и обеспечивает очень эффективную электрическую изоляцию. Последняя предупреждает утечку внутриклеточного компонента локального тока из аксоплазмы, "удерживая" его внутри волокна, и обеспечивает его распространение. У беспозвоночных такого специфического и эффективного механизма повышения скорости проведения не развилось. Однако когда мы будем рассматривать функцию некоторых сенсилл насекомых в разделе ВКУС , мы увидим, что существуют альтернативные средства изоляции локальных токов, обеспечивающие эффективное электротоническое проведение в длинных внешних сегментах хемочувствительных клеток. Табл. 2.2 показывает скорость проведения импульсов в афферентных волокнах млекопитающих. Афферентные волокна мышц состоят из четырех перекрывающихся классов (I - IV), чья скорость проведения состовляет от 120 м/с в наиболее крупных миелинизированных волокон (диаметр ок. 20 мкм) до прибл. 0,5 м/с в самых тонких немиелинизированных (диаметр ок. 0,5 мкм). В сенсорных нервах кожи отсутствуют наиболее быстрые волокна класса I (или Аальфа ). Кожные нервы , таким образом, подразделяются на три группы A бета , A дельта и C , что соответствует группам II, III и IV мышечных афферентов.
Проведение нервных импульсов по нервным волокнам и через синапсы. Высоковольтный потенциал, возникающий при возбуждении рецептора в нервном волокне, в 5—10 раз больше порога раздражения рецептора. Проведение волны возбуждения по нервному волокну обеспечивается тем, что каждый последующий его участок раздражается высоковольтным потенциалом предыдущего участка. В мякотных нервных волокнах этот потенциал распространяется не непрерывно, а скачкообразно; он перескакивает через один или даже несколько перехватов Ранвье, в которых усиливается. Продолжительность проведения возбуждения между двумя соседними перехватами Ранвье равняется 5—10% длительности высоковольтного потенциала.
Проведение нервного импульса по нервному волокну происходит только при условии его анатомической непрерывности и нормального физиологического его состояния. Нарушение физиологических свойств нервного волокна сильным охлаждением или отравлением ядами и наркотиками прекращает проведение нервного импульса даже при анатомической его непрерывности.
Нервные импульсы проводятся изолированно по отдельным двигательным и чувствительным нервным волокнам, которые входят в состав смешанного нерва, что зависит от изолирующих свойств покрывающих их миелиновых оболочек. В безмякотных нервных волокнах биоток распространяется непрерывно вдоль волокна и благодаря соединительнотканой оболочке не переходит с одного волокна на другое. Нервный импульс может распространяться по нервному волокну в двух направлениях: центростремительном и центробежном. Следовательно, существуют три правила проведения нервного импульса в нервных волокнах: 1) анатомической непрерывности и физиологической целости, 2) изолированного проведения и 3) двустороннего проведения.
Через 2—3 дня после отделения нервных волокон от тела нейрона они начинают перерождаться, или дегенерировать, и проведение нервных импульсов прекращается. Нервные волокна и миелин разрушаются и сохраняется только соединительнотканая оболочка. Если соединить перерезанные концы нервных волокон, или нерва, то после дегенерации тех участков, которые отделены от нервных клеток, начинается восстановление, или регенерация, нервных волокон со стороны тел нейронов, из которых они прорастают в сохранившиеся соединительнотканые оболочки. Регенерация нервных волокон приводит к восстановлению проведения импульсов.
В отличие от нервных волокон через нейроны нервной системы нервные импульсы проводятся только в одном направлении — от рецептора к работающему органу. Это зависит от характера проведения нервного импульса через синапсы. В нервном волокне над пресинаптической мембраной есть множество мельчайших пузырьков ацетилхолина. При достижении биотоком пресинаптической мембраны часть этих пузырьков лопается, и ацетилхолин проходит через мельчайшие отверстия в пресинаптической мембране в синаптическую щель.
В постсинаптической мембране имеются участки, обладающие особым сродством к ацетилхолину, который вызывает временное появление пор в постсинаптической мембране, отчего она становится временно проницаемой для ионов. В результате в постсинаптической мембране возникает возбуждение и высоковольтный потенциал, который распространяется по следующему нейрону или по иннервируемому органу. Следовательно, передача возбуждения через синапсы происходит химическим путем посредством посредника, или медиатора, ацетилхолина, а проведение возбуждения по следующему нейрону снова осуществляется электрическим путем.
Действие ацетилхолина на проведение нервного импульса через синапс кратковременно; он быстро разрушается, гидролизуется ферментом холинэстеразой.
Так как химическая передача нервного импульса в синапсе происходит в течение доли мсек, то в каждом синапсе нервный импульс на это время задерживается.
Нервный импульс
Нервный импульс представляет собой наиболее быстрый способ передачи информации в организме. [1]
Нервный импульс всегда распространяется в одном направлении: по дендриту - к телу клетки, по аксону - от тела. Поскольку дендритов у каждого нейрона много, а аксон всего один, наблюдается следующая закономерность, характерная как для отдельной нервной клетки, так и для всей нервной системы в целом: входов много, выход - один, хотя, понятно, один - не значит, что обеспечивает его единственная клетка, ведь в любом конкретном нерве и нейронов и аксонов много. [2]
Нервные импульсы должны передаваться от одной клетки к другой. Аксон может быть связан с сомой второй клетки через аксо-соматический синапс. Имеются также аксо-дендритные и аксо-аксонные синапсы. [4]
Нервные импульсы распространяются не только внутри клетки, но должны также передаваться соседним клеткам. Специализированная структура для межклеточной передачи называется синапсом. [6]
Нервные импульсы передаются только в направлении от пресинаптической к постсинаптической мембране, и, таким образом, синапсы выполняют роль выпрямителей в процессах проводимости. [7]
Нервный импульс ( возбуждение) передается по этой цепи комплексно - электрическим и химическим способами. Передача по аксону имеет электрическую природу, то есть в первом приближении аналогична передаче электрического тока по проводнику. В межнейрои-ных, нерьно-мыщечных и нервно-репепторных синапсах связь между разобщенными звеньями депи нервной системы осуществляется при помощи химических веществ - передатчиков нервных импульсов или медиаторов. Медиаторы находятся в специальных пузырьках в области окончаний нервных волокон. Под влиянием поступающего по аксону импульса они выделяются в сннаптнческую щель через пресинаптнческую мембрану ( мембрану перед синаптической щелью), возбуждают мембрану дендрита следующей нервной клетки или рецептора ( постсннаптическую мембрану) н обеспечивают таким образом прохождение нервного импульса дальше. [8]
Нервный импульс , прибывающий к нервному окончанию, вызывает поток ионов Са через их мембрану, что способствует соединению синаптических пузырьков с мембраной и выбросу путем экзоцитоза содержащихся в них молекул ацетилхолина в синаптическую щель. Молекулы ацетилхолина диффундируют к мембране мышечного волокна и примерно через 0 3 мс соединяются с рецепторными молекулами, которые открывают натриевые каналы в мембране мышцы. При каждом импульсе открывается около 2000 каналов. [9]
Нервный импульс поступает внутрь мышечного волокна насекомого через систему поперечных трубок ( Г - систему) так же, как в поперечнополосатых мышцах позвоночных животных. Однако если в мышцах скелетных животных трубки Г - сис-темы лежат либо в области Z-пластинок ( см. рис. 56), либо в областях границ А - и / - зон, то в мышцах насекомых трубки Г - сис-темы лежат на половине расстояния между Z-пластинкой и Я-зоной. [10]
Нервный импульс представляет сложнейший физико-химический процесс, связанный с перемещением некоторых минеральных веществ, в частности ионов калия и натрия. В состоянии покоя ионы калия находятся преимущественно внутри нервной клетки, ионы натрия - на ее наружной поверхности. В протоплазме нервных клеток ионов калия примерно в 30 - 40 раз больше, чем в окружающей клетку тканевой жидкости, ионов же натрия в 8 - 10 раз меньше. В соответствии с этим внутри клетки преобладают отрицательные электрические заряды, BSD ее - положительные. В тот момент, когда нервный импульс приходит в окончание аксона ( так называемую синаптическую бляшку), пузырьки, содержащие медиатор, лопаются. Химический передатчик изливается в синаптическую щель и изменяет проницаемость постсин-аптической мембраны. Это ведет к тому, что ионы калия устремляются из клетки и располагаются на ее поверхности, обращенной к щели, а ионы натрия входят в клетку. Электрический заряд мембраны мгновенно изменяется, возникает разница потенциалов, и импульс переходит с аксона одной клетки на дендрит другой. Как только импульс прошел синапс, медиатор разрушается, ионы калия снова поступают в клетку, а ионы натрия выходят из нее. [11]
Нервные импульсы и медленные потенциалы являются двумя видами процессов, которые могут функционировать реципрокно. Естественно было бы предположить, что, чем эффективнее осуществляется процесс превращения структуры афферентных возбуждений в эфферентные, тем менее долговечна композиция, сформированная микроструктурой медленных потенциалов. При озникновении навыка и привыкания поведение становится рефлекторным - при этом более или менее устойчивые композиции узора медленных потенциалов соответствуют осознанию. Из этого следует вывод, что сами по сзбе динамические структуры нервных импульсов и поведение, которое ими вызывается, непосредственно не осознаются. Так, даже процесс речи остается неосозна-оаемым в тот момент, когда слова произносятся. [12]
Нервные импульсы переплетаются с гуморальным, Гуморальные вызывают нервные. [13]
Нервный импульс ( возбуждение) передается по этой цепи комплексно - электрическим и химическим способами. Передача по аксону имеет электрическую природу, то есть в первом приближении аналогична передаче электрического тока по проводнику. В межнейронных, нервно-мышечных и нервно-рецепторных синапсах связь между разобщенными звеньями цепи нервной системы осуществляется при помощи химических веществ - передатчиков нервных импульсов или медиаторов. Медиаторы находятся в специальных пузырьках в области окончаний нервных волокон. Под влиянием поступающего по аксону импульса они выделяются в синаптическую щель через пресинаптическую мембрану ( мембрану перед синаптической щелью), возбуждают мембрану дендрита следующей нервной клетки или рецептора ( постсинаптическую мембрану) и обеспечивают таким образом прохождение нервного импульса дальше. [14]
Нервные импульсы , поступающие по зрительному нерву в мозг, создают в нем определенное представление о размере, форме, объеме и цвете наблюдаемого предмета. [15]
Нервный импульс (лат. nervus нерв; лат. impulsus удар, толчок) — волна возбуждения, распространяющаяся по нервному волокну; единица распространяющегося возбуждения.
Нервный импульс обеспечивает передачу информации от рецепторов к нервным центрам и от них к исполнительным органам — скелетной мускулатуре, гладким мышцам внутренних органов и сосудов, железам внутренней и внешней секреции и т. д.
Распространение Нервных импульсов отождествляется с проведением потенциалов действия (см. Биоэлектрические потенциалы). Возникновение возбуждения может быть результатом раздражения (см.), напр, воздействие света на зрительный рецептор, звука на слуховой рецептор, или процессов, протекающих в тканях (спонтанное возникновение Н. и.). В этих случаях Н. и. обеспечивают согласованную работу органов при протекании какого-либо физиологического процесса (напр., в процессе дыхания Н. и. вызывают сокращение скелетных мышц и диафрагмы, результатом чего являются вдох и выдох, и т. д.).
В живых организмах передача информации может осуществляться и гуморальным путем, посредством выброса в русло крови гормонов, медиаторов и т. п. Однако преимущество информации, передаваемой при помощи Н. и., состоит в том, что она более целенаправленна, передается быстро и может быть точнее закодирована, чем сигналы, посылаемые гуморальной системой.
Факт, что нервные стволы являются путем, по к-рому передаются влияния от мозга к мышцам и в обратном направлении, был известен еще в эпоху античности. В средние века и вплоть до середины 17 в. считалось, что по нервам распространяется некая субстанция, подобная жидкости или пламени. Идея о электрической природе Н. и. возникла в 18 в. Первые исследования электрических явлений в живых тканях, связанных с возникновением и распространением возбуждения, были осуществлены Л. Гальвани. Г. Гельмгольц показал, что скорость распространения Н. и., к-рую ранее считали близкой к скорости света, имеет конечное значение и может быть точно измерена. Германн (L. Hermann) ввел в физиологию понятие потенциала действия. Объяснение механизма возникновения и проведения возбуждения стало возможным после создания С. Аррениусом теории электролитической диссоциации. В соответствии с этой теорией Бернштейн (J. Bernstein) предположил, что возникновение и проведение Н. и. обусловлено перемещением ионов между нервным волокном и окружающей средой. Англ. исследователи А. Ходжкин, Б. Катц и Э. Хаксли детально исследовали трансмембранные ионные токи, лежащие в основе развития потенциала действия. Позже стали интенсивно изучаться механизмы работы ионных каналов, по к-рым происходит обмен ионами между аксоном и окружающей средой, и механизмы, обеспечивающие способность нервных волокон проводить ряды Н. и. разного ритма и продолжительности.
Н. и. распространяется за счет местных токов, возникающих между возбужденным и невозбужденным участками нервного волокна. Ток, выходящий из волокна наружу в покоящемся участке, служит раздражителем. Наступающая после возбуждения в данном участке нервного волокна рефрактерность обусловливает поступательное движение Н. и.
Количественно соотношения разных фаз развития потенциала действия можно охарактеризовать, сопоставляя их по амплитуде и длительности во времени. Так, напр., для миелиновых нервных волокон группы А млекопитающих диаметр волокна находится в пределах 1—22 мк, скорость проведения — 5—120 м/сек, длительность и амплитуда высоковольтной части (пика, или спайка) — 0,4—0,5 мсек и 100—120 мв соответственно, следовой негативный потенциал — 12—20 мсек (3—5% от амплитуды спайка), следовой позитивный потенциал — 40—60 мсек (0,2% от амплитуды спайка).
Возможности передачи разнообразной информации расширяются за счет повышения скорости развития потенциала действия, скорости распространения, а также за счет повышения лабильности (см.) — т. е. способности возбудимого образования воспроизводить в единицу времени высокие ритмы возбуждения.
Возникновение Н. и. в нервных клетках (см.) или рецепторах (см.) связано с деполяризацией мембраны, т. е. со снижением величины электрического потенциала на мембране (потенциала покоя, или мембранного потенциала). Если величина мембранного потенциала снижается на 10—20% (пороговый критический уровень), то местный процесс переходит в распространяющийся — возникает потенциал действия (см. Возбуждение).
Конкретные особенности распространения Н. и. связаны со строением нервных волокон (см.). Сердцевина волокна (аксоплазма) обладает низким сопротивлением и, соответственно, хорошей проводимостью, а окружающая аксоплазму плазматическая мембрана — большим сопротивлением. Особенно велико электрическое сопротивление наружного слоя у миелинизированных волокон, у к-рых свободны от толстой миелиновой оболочки только перехваты Ранвье. В безмиелиновых волокнах Н. и. движется непрерывно, а в миелиновых — скачкообразно (сальтаторное проведение).
Различают декрементное и бездекрементное распространение волны возбуждения. Декрементное проведение, т. е. проведение возбуждения с угасанием, наблюдается в безмиелиновых волокнах. В таких волокнах скорость проведения Н. и. невелика и по мере отдаления от места раздражения раздражающее действие местных токов постепенно уменьшается вплоть до полного угасания. Декрементное проведение свойственно волокнам, иннервирующим внутренние органы, обладающие низкой функц, подвижностью. Без декрементное проведение характерно для миелиновых и тех безмиелиновых волокон, к-рые передают сигналы к органам, обладающим высокой реактивностью (напр., сердечной мышце). При бездекрементном проведении Н. и. проходит весь путь от места раздражения до места реализации информации без затухания.
Передача Н. и. с нервного волокна на мышечное или какой-либо другой эффектор осуществляется через синапсы (см.). У позвоночных животных в подавляющем большинстве случаев передача возбуждения на эффектор происходит при помощи выделения ацетилхолина (нервно-мышечные синапсы скелетной мускулатуры, синаптические соединения в сердце и др.). Для таких синапсов характерно строго одностороннее проведение импульса и наличие временной задержки передачи возбуждения.
В синапсах, в синаптической щели которых сопротивление электрическому току благодаря большой площади контактирующих поверхностей мало, происходит электрическая передача возбуждения. В них нет синаптической задержки проведения и возможно двустороннее проведение. Такие синапсы свойственны беспозвоночным животным.
Регистрация Н. и. нашла широкое применение в биол, исследованиях и клин, практике. Для регистрации используют шлейфные и чаще катодные осциллографы (см. Осциллография). При помощи микроэлектродной техники (см. Микроэлектродный метод исследования) регистрируют Н. и. в одиночных возбудимых образованиях — нейронах и аксонах. Возможности исследования механизма возникновения и распространения Н. и. значительно расширились после разработки метода фиксации потенциала. Этим методом были получены основные данные о ионных токах (см. Биоэлектрические потенциалы).
Нарушение проведения Н. и. происходит при повреждении нервных стволов, напр, при механических травмах, сдавливании в результате разрастания опухоли или при воспалительных процессах. Такие нарушения проведения Н. и. зачастую бывают необратимы. Следствием прекращения иннервации могут быть тяжелые функциональные и трофические расстройства (напр., атрофия скелетных мышц конечностей после прекращения поступления Н. и. вследствие необратимой травмы нервного ствола). Обратимое прекращение проведения Н. и. может быть вызвано специально, в терапевтических целях. Напр., с помощью анестезирующих средств блокируют импульсацию, идущую от болевых рецепторов в ц. н. с. Обратимое прекращение проведения Н. и. вызывает и новокаиновая блокада. Временное прекращение передачи Н. и. по нервным проводникам наблюдается и во время общего наркоза.
Библиография: Бpеже М. А. Электрическая активность нервной системы, пер. с англ., М., 1979; Жуков Е. К. Очерки по нервно-мышечной физиологии, Л., 1969; Коннели К. Восстановительные процессы и обмен веществ в нерве, в кн.: Совр, пробл. биофизики, пер. с англ., под ред. Г. М. Франка и А. Г. Пасынского, т. 2, с. 211, М., 1961; Костюк П. Г. Физиология центральной нервной системы, Киев, 1977; Латманизова Л. В. Очерк физиологии возбуждения, М., 1972; Общая физиология нервной системы, под ред. П. Г. Костюка, Л., 1979; Тасаки И. Нервное возбуждение, пер. с англ., М., 1971; Ходжкин А. Нервный импульс, пер. с англ., М., 1965; Ходоров Б. И. Общая физиология возбудимых мембран, М., 1975.
Нервная система человека выступает своеобразным координатором в нашем организме. Она передаёт команды от мозга мускулатуре, органам, тканям и обрабатывает сигналы, идущие от них. В качестве своеобразного носителя данных используется нервный импульс. Что он собой представляет? С какой скоростью работает? На эти, а также на ряд других вопросов можно будет найти ответ в этой статье.
Чем является нервный импульс?
Исследование строения и работы
Впервые прохождение нервного импульса было продемонстрировано немецкими учеными Э. Герингом и Г. Гельмгольцем на примере лягушки. Тогда же и было установлено, что биоэлектрический сигнал распространяется с указанной ранее скоростью. Вообще, такое является возможным благодаря особенному построению нервных волокон. В некотором роде они напоминают электрический кабель. Так, если проводить параллели с ним, то проводниками являются аксоны, а изоляторами – их миелиновые оболочки (они являют собой мембрану шванновской клетки, которая намотана в несколько слоев). Причем скорость нервного импульса зависит в первую очередь от диаметра волокон. Вторым по важности считается качество электрической изоляции. Кстати, в качестве материала организмом используется липопротеид миелин, который обладает свойствами диэлектрика. При прочих равных условиях, чем больше будет его слой, тем быстрее будут проходить нервные импульсы. Даже на данный момент нельзя сказать, что эта система полноценно исследована. Многое, что относится к нервам и импульсам, ещё остаётся загадкой и предметом исследования.
Особенности строения и функционирования
Где они создаются?
Типы клеток
- Рецепторные (чувствительные). Ими кодируются и превращаются в нервные импульсы все температурные, химические, звуковые, механические и световые раздражители.
- Вставочные (также называются кондукторными или замыкательными). Они служат для того, чтобы перерабатывать и переключать импульсы. Наибольшее их число находится в головном и спинном мозге человека.
- Эффекторные (двигательные). Они получают команды от центральной нервной системы на то, чтобы были совершены определённые действия (при ярком солнце закрыть рукой глаза и так далее).
Каждый нейрон имеет тело клетки и отросток. Путь нервного импульса по телу начинается именно с последнего. Отростки бывают двух типов:
- Дендриты. На них возложена функция восприятия раздражения расположенных на них рецепторов.
- Аксоны. Благодаря им нервные импульсы передаются от клеток к рабочему органу.
Интересный аспект деятельности
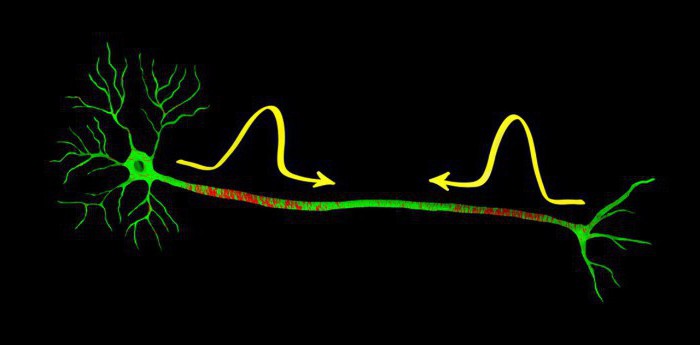
О потенциале действия
Как всё работает в мозгу?
Работа нейромедиаторов
Когда они передают нервные импульсы, то существует несколько вариантов, что произойдёт с ними:
- Они будут диффундированы.
- Подвергнутся химическому расщеплению.
- Вернутся назад в свои пузырьки (это называется обратным захватом).
В конце 20-го века сделали поразительное открытие. Ученые узнали, что лекарства, что влияют на нейромедиаторы (а также их выброс и обратный захват), могут изменять психическое состояние человека коренным образом. Так, к примеру, ряд антидепрессантов вроде "Прозака" блокируют обратный захват серотонина. Есть определённые причины считать, что в болезни Паркинсона виноват дефицит в головном мозге нейромедиатора дофамина.
Если кратко, то они могут работать с тысячами нейромедиаторов, которые посылаются их соседями. Детали относительно обработки и интеграции данного типа импульсов нам почти не известны. Хотя над этим работает много исследовательских групп. На данный момент получилось узнать, что все полученные импульсы интегрируются, а нейрон выносит решение – необходимо ли поддерживать потенциал действия и передавать их дальше. На этом фундаментальном процессе базируется функционирование головного мозга человека. Ну что ж, тогда это неудивительно, что мы не знаем ответа на эту загадку.
Некоторые теоретические особенности
Где же создаются нервные импульсы?
Откуда они начинают свой путь? Ответ на этот вопрос может дать любой студент, который прилежно изучал физиологию возбуждения. Есть четыре варианта:
- Рецепторное окончание дендрита. Если оно есть (что не факт), то возможным является наличие адекватного раздражителя, что создаст сначала генераторный потенциал, а потом уже и нервный импульс. Подобным образом работают болевые рецепторы.
- Мембрана возбуждающего синапса. Как правило, такое возможно только при наличии сильного раздражения или их суммирования.
- Триггерная зона дентрида. В этом случае локальные возбуждающие постсинаптические потенциалы формируются как ответ на раздражитель. Если первый перехват Ранвье миелинизирован, то они на нём суммируются. Благодаря наличию там участка мембраны, которая обладает повышенной чувствительностью, здесь возникает нервный импульс.
- Аксонный холмик. Так называют место, где начинается аксон. Холмик – это наиболее частый создать импульсов на нейроне. Во всех остальных местах, которые рассматривались ранее, их возникновение гораздо менее вероятное. Это происходит из-за того, что здесь мембрана имеет повышенную чувствительность, а также пониженный критический уровень деполяризации. Поэтому, когда начинается суммирование многочисленных возбуждающих постсинаптических потенциалов, то раньше всего на них реагирует холмик.
Пример распространяющегося возбуждения
Вспомните сводки из новостей прошлого лета (также это скоро можно будет услышать опять). Пожар распространяется! При этом деревья и кустарники, которые горят, остаются на своих местах. А вот фронт огня идёт всё дальше от места, где был очаг возгорания. Аналогичным образом работает нервная система.
Часто бывает необходимо успокоить начавшееся возбуждение нервной системы. Но это не так легко сделать, как и в случае с огнем. Для этого совершают искусственное вмешательство в работу нейрона (в лечебных целях) или используют различные физиологические средства. Это можно сравнить с заливанием пожара водой.
Нервные импульсы распространяются при перемещении ионов через мембрану нервной клетки и передаются из одной нервной клетки в другую с помощью нейромедиаторов.
В результате эволюции нервной системы человека и других животных возникли сложные информационные сети, процессы в которых основаны на химических реакциях. Важнейшим элементом нервной системы являются специализированные клетки нейроны. Нейроны состоят из компактного тела клетки, содержащего ядро и другие органеллы. От этого тела отходит несколько разветвленных отростков. Большинство таких отростков, называемых дендритами, служат точками контакта для приема сигналов от других нейронов. Один отросток, как правило самый длинный, называется аксоном и передает сигналы на другие нейроны. Конец аксона может многократно ветвиться, и каждая из этих более мелких ветвей способна соединиться со следующим нейроном.
Во внешнем слое аксона находится сложная структура, образованная множеством молекул, выступающих в роли каналов, по которым могут поступать ионы — как внутрь, так и наружу клетки. Один конец этих молекул, отклоняясь, присоединяется к атому-мишени. После этого энергия других частей клетки используется на то, чтобы вытолкнуть этот атом за пределы клетки, тогда как процесс, действующий в обратном направлении, вводит внутрь клетки другую молекулу. Наибольшее значение имеет молекулярный насос, который выводит из клетки ионы натрия и вводит в нее ионы калия (натрий-калиевый насос).
Когда клетка находится в покое и не проводит нервных импульсов, натрий-калиевый насос перемещает ионы калия внутрь клетки и выводит ионы натрия наружу (представьте себе клетку, содержащую пресную воду и окруженную соленой водой). Из-за такого дисбаланса разность потенциалов на мембране аксона достигает 70 милливольт (приблизительно 5% от напряжения обычной батарейки АА).
Однако при изменении состояния клетки и стимуляции аксона электрическим импульсом равновесие на мембране нарушается, и натрий-калиевый насос на короткое время начинает работать в обратном направлении. Положительно заряженные ионы натрия проникают внутрь аксона, а ионы калия откачиваются наружу. На мгновение внутренняя среда аксона приобретает положительный заряд. При этом каналы натрий-калиевого насоса деформируются, блокируя дальнейший приток натрия, а ионы калия продолжают выходить наружу, и исходная разность потенциалов восстанавливается. Тем временем ионы натрия распространяются внутри аксона, изменяя мембрану в нижней части аксона. При этом состояние расположенных ниже насосов меняется, способствуя дальнейшему распространению импульса. Резкое изменение напряжения, вызванное стремительными перемещения ионов натрия и калия, называют потенциалом действия. При прохождении потенциала действия через определенную точку аксона, насосы включаются и восстанавливают состояние покоя.
Когда импульс достигает конца основной части тела аксона, его необходимо передать либо следующему нижележащему нейрону, либо, если речь идет о нейронах головного мозга, по многочисленным ответвлениям многим другим нейронам. Для такой передачи используется абсолютно иной процесс, нежели для передачи импульса вдоль аксона. Каждый нейрон отделен от своего соседа небольшой щелью, называемой синапсом. Потенциал действия не может перескочить через эту щель, поэтому нужно найти какой-то другой способ для передачи импульса следующему нейрону. В конце каждого отростка имеются крошечные мешочки, называющие (пресинаптическими) пузырьками, в каждом из которых находятся особые соединения — нейромедиаторы. При поступлении потенциала действия из этих пузырьков высвобождаются молекулы нейромедиаторов, пересекающие синапс и присоединяющиеся к специфичным молекулярным рецепторам на мембране нижележащих нейронов. При присоединении нейромедиатора равновесие на мембране нейрона нарушается. Сейчас мы рассмотрим, возникает ли при таком нарушении равновесия новый потенциал действия (нейрофизиологи продолжают искать ответ на этот важный вопрос до сих пор).
После того как нейромедиаторы передадут нервный импульс от одного нейрона на следующий, они могут просто диффундировать, или подвергнуться химическому расщеплению, или вернуться обратно в свои пузырьки (этот процесс нескладно называется обратным захватом). В конце XX века было сделано поразительное научное открытие — оказывается, лекарства, влияющие на выброс и обратный захват нейромедиаторов, могут коренным образом изменять психическое состояние человека. Прозак (Prozac*) и сходные с ним антидепрессанты блокируют обратный захват нейромедиатора серотонина. Складывается впечатление, что болезнь Паркинсона взаимосвязана с дефицитом нейромедиатора допамина в головном мозге. Исследователи, изучающие пограничные состояния в психиатрии, пытаются понять, как эти соединения влияют на человеческий рассудок.
Читайте также: