
Область окончаний эфферентных нервов это
Глава 7. Вещества, действующие преимущественно в области окончаний эфферентных нервов
Настоящий раздел посвящен лекарственным веществам, оказывающим влияние преимущественно в области окончаний эфферентных (центробежных) нервов, иначе говоря, в области эффекторной части рефлекторной дуги.
Изучение физиологической роли нервов, снабжающих внутренние органы, показало, что многие из них несут импульсы от центральной нервной системы к исполнительным органам. Нервные импульсы, доходя до органов, вызывают в них те или иные реакции. Чаще всего это сокращение или расслабление мускулатуры органа либо усиление его секреторной функции. Кроме того, эти нервы осуществляют также и трофическую функцию, так как они оказывают влияние и на обмен веществ в органах и тканях.
Многие из эфферентных нервов принадлежат к вегетативному отделу нервной системы. К ним относятся также и нервы, иннервирующие скелетную мускулатуру (двигательные нервы).
Вегетативный отдел нервной системы принимает большое участие в регуляции функций организма в соответствии с изменяющимися условиями внешней среды. Вегетативную нервную систему принято делить на два отдела: парасимпатический и симпатический. Большинство органов получает иннервацию от обоих систем, т. е. имеет место двойная иннервация органов. Импульсы, идущие к органам по нервным стволам этих систем, часто противоположно влияют на эти органы и системы (табл. 1).
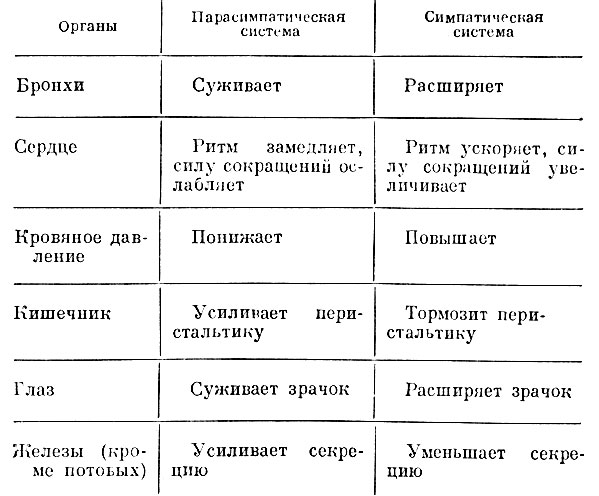
Таблица 1. Сравнительная характеристика влияния парасимпатической и симпатической нервной системы на некоторые органы и системы
Если импульсы одной системы являются пусковыми для данного-органа, импульсы другой системы будут тормозить его деятельность. Такая двойная иннервация органов обеспечивает смену возбудительного и тормозного процессов в органах и тканях в соответствии с постоянно изменяющейся деятельностью целостного организма, что позволяет тонка регулировать его жизнедеятельность, обеспечивая при этом постоянство внутренней среды организма (более подробное освещение вопроса не входит в задачу фармакологии).
В вегетативном отделе нервной системы различают центральную часть, представляющую собой скопления нервных клеток, расположенных на различных уровнях центральной нервной системы. Таким образом, первый нейрон находится в центральной нервной системе. Однако для вегетативного отдела нервной системы характерна двухнейронная система передачи возбуждения от центра на периферию. Нервные волокна первого нейрона заканчиваются у клеток ганглиев. Эти волокна носят название преганглионарных волокон. Вегетативные ганглии или узлы представляют собой скопления нервных клеток (своего рода промежуточную станцию на пути от центра к периферии), отростки которых (аксоны), образуя нервные стволы, осуществляют непосредственно иннервацию органов. Эти волокна называют постганглионарными, т. е. идущими после ганглиев. Двигательные нервы скелетных мышц идут от нейронов, лежащих в центральной нервной системе, до периферии, не прерываясь.
Центральная часть парасимпатического отдела вегетативной нервной системы распадается на краниальную часть (черепную), расположенную в среднем и заднем мозге, и крестцовую.
Парасимпатические нервы выходят в составе трех пар черепных нервов: глазодвигательного (снабжают внутренние мышцы глаза), лицевого (несущего секреторные волокна к подчелюстным слюнным железам) и блуждающего (иннервирует бронхи, сердце и желудочно-кишечный тракт). Крестцовый отдел парасимпатической нервной системы иннервирует нижний отдел кишечника и тазовые органы.
Центры симпатической нервной системы заложены в подкорковых узлах, промежуточном, продолговатом и спинном мозгу. Симпатические нервы выходят из мозга в составе корешков всех грудных и первых четырех поясничных спинномозговых нервов.
Для понимания механизма действия лекарственных веществ, оказывающих влияние в области окончаний эфферентных нервов, большое значение сыграло открытие роли химических веществ в передаче возбуждения с нервных окончаний на иннервируемые ими клетки и органы. До недавнего времени господствовало представление, что лекарственные вещества оказывают непосредственное влияние на окончания эфферентных нервов, возбуждая или парализуя их, что соответственно должно было изменять и функции иннервируемых ими органов. Однако, как показали дальнейшие исследования, передача нервного импульса на рабочий орган осуществляется при участии химических веществ. Под влиянием нервного импульса в области окончаний нервов образуются химические вещества. Последние являются непосредственными передатчиками возбуждения с нерва на клетки исполнительного органа или на ганглионарные клетки. Эти вещества получили название медиаторов (посредников). Роль медиаторов заключается в том, что они вступают в реакцию с биохимическими системами клеток, вызывая возникновение ответной реакции (в виде усиления или торможения функции) в соответствии с той ролью, которую играет нерв в отношении данного органа (теория нейро-гуморальной передачи возбуждения).
По характеру образующихся медиаторов все эфферентные нервы делят на холинергичес кие и адренергичес-кие. У окончаний холинергическихнервов образуется медиатор ацетилхолин, у окончаний адренергических нервов выделяется норадреналин или в некоторых случаях адреналин. Биохимические системы клеток, вступающие в реакцию с ацетилхолином, по терминологии С. В. Аничкова, носят название холинореактивных систем, а реагирующие с норадреналином или адреналином — адренореактивных систем. Необходимыми условиями для образования химического медиатора в организме является наличие нервного импульса и целостность нервных окончаний.
К холинергическим нервам относят все преганглионарные волокна парасимпатической и симпатической нервной системы, постганглионар-ные волокна парасимпатической системы и двигательные нервы скелетной мускулатуры. Многими исследователями показано, что ацетилхолин принимает участие в межнейронной передаче нервного возбуждения и в центральной нервной системе.
К адренергическим нервам относятся все постганглионарные волокна симпатической нервной системы, за исключением волокон, снабжающих потовые железы. Последние являются холинергическими волокнами.
Введенные извне ацетилхолин и норадреналин, будучи по химической структуре идентичными естественным медиаторам, воспроизводят в организме эффекты, присущие действию этих медиаторов.
Кроме ацетилхолина, норадреналина и адреналина, к медиаторам нервных импульсов, но действующим в центральной нервной системе, относится серотонин (5-окситриптамин).
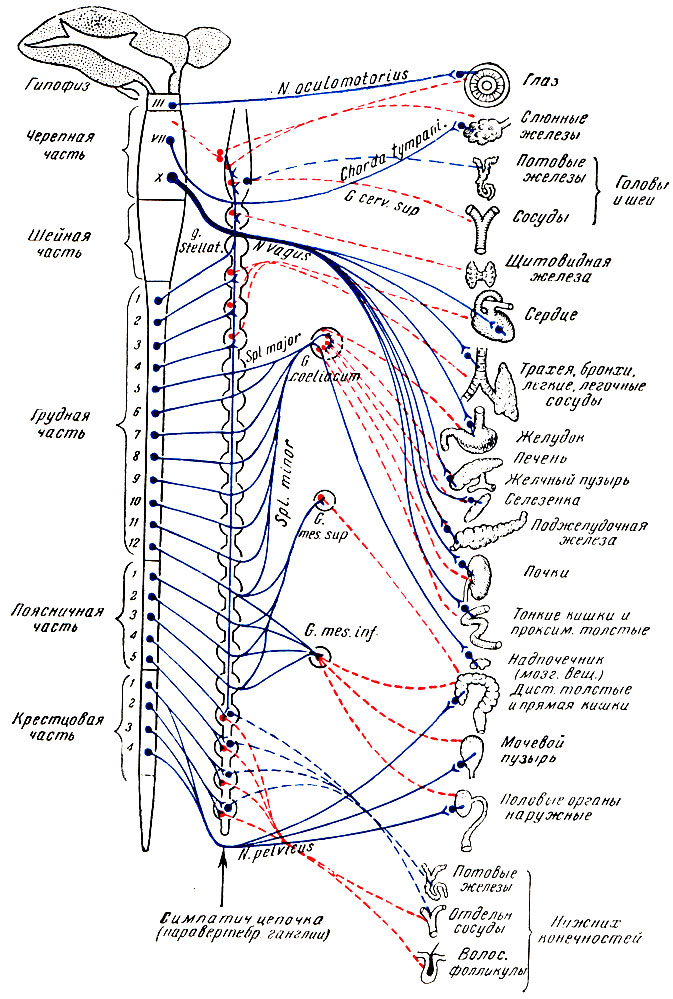
Рис. 23. Схема вегетативной нервной системы. Синим показаны холинергические нервы; красным — адренергические; непрерывными линиями — преганглионарные и прерывистыми — постганглионарные волокна (по М. П. Николаеву).
Медиаторы претерпевают в тканях определенный цикл биохимических превращений. Они возникают в результате ряда ферментативных реакций и разрушаются также при участии ферментов.
Ацетилхолин образуется в результате ацетилирования холина под воздействием фермента холинацетилазы:
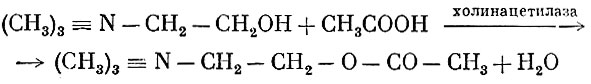
и быстро омыляется в тканях благодаря действию холинэстеразы
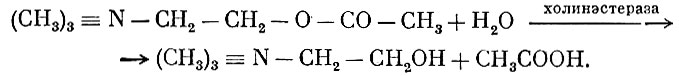
Различают истинную холинэстеразу, действующую только на аце-тилхолин, и ложную холинэстеразу, которая катализирует омыление наряду с ацетилхолином многих других сложных эфиров.
Норадреналин образуется в тканях из тирозина в результате окисления и декарбоксилирования приблизительно по следующей схеме:
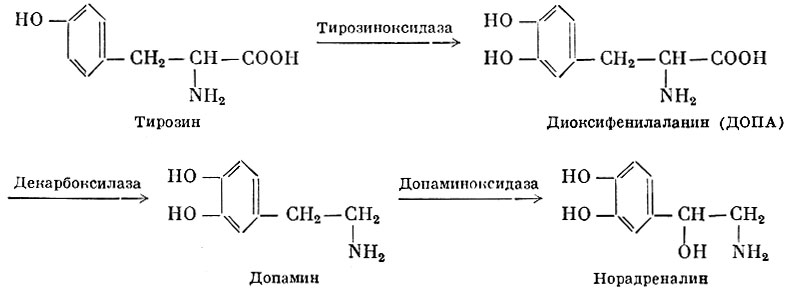
В тканях Норадреналин быстро инактивируется (окисляется) с помощью фермента моноаминооксидазы.
Серотонин образуется в организме из триптофана путем ряда ферментативных превращений:
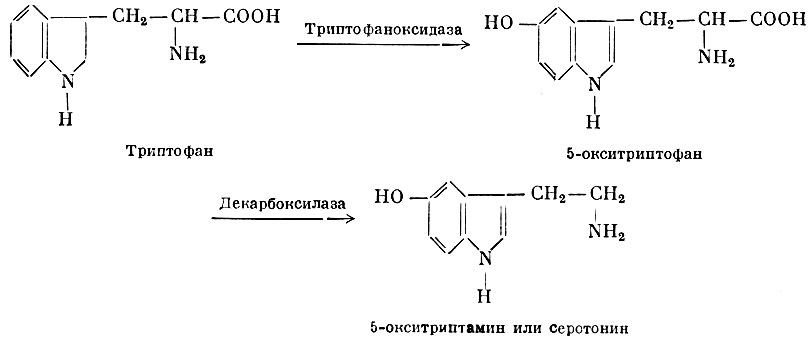
5-окситриптамин или Серотонин
Серотонин разрушается (окисляется) в тканях также с помощью моноаминооксидазы. В качестве лекарственных препаратов используются как сами медиаторы, так и вещества, действующие сходно с ними, имитирующие их действие. Вещества, сходные по действию с ацетил-холином, называют ход и но ми мет и чески ми, а сходные по действию с норадреналином — адреномвметическипи. Наряду с этим имеется большая группа веществ, введение которых в организм препятствует или задерживает проявление действия медиатора. Так, вещества, устраняющие эффект ацетилхолина, называют холинолитическими. Вещества, вызывающие тот же эффект в отношении норадреналина и адреналина, носят название а д р е-нолитических.
Особую группу составляют вещества, обладающие способностью блокировать ферменты, инактивирующие медиаторы (холинэстеразу или моноаминооксидазу). В результате действие естественного медиатора усиливается и становится более длительным (пролонгируется). Рассмотрим упомянутые группы веществ.
Эфферентные (эффекторные) нервные окончания в зависимости от природы иннервируемого органа подразделяются на двигательные и секреторные. Двигательные окончания имеются в поперечнополосатых и гладких мышцах, секреторные - в железах.
Нервно-мышечное окончание (нервно-мышечный синапс, моторная бляшка) - двигательное окончание аксона мотонейрона на волокнах поперечнополосатых соматических мышц - состоит из концевого ветвления аксона, образующего пресинаптическую часть, специализированного участка на мышечном волокне, соответствующего постсинаптйческой части, и разделяющей их синаптической щели (рис. 8.18.).
В крупных мышцах, развивающих значительную силу, один аксон, разветвляясь, иннервирует большое количество (сотни и тысячи) мышечных волокон. Напротив, в мелких мышцах, осуществляющих тонкие движения (например, наружных мышцах глаза), каждое волокно или их небольшая группа иннервируются отдельным аксоном. Один мотонейрон в совокупности с иннервируемыми им мышечными волокнами образует двигательную единицу.
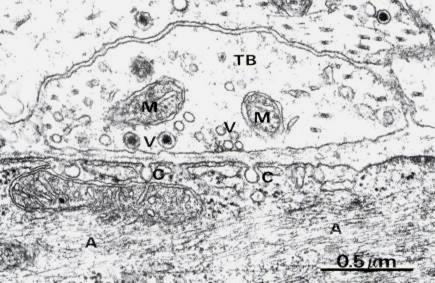
Рис. 8.18. Нервно мышечный синапс между терминалями аксона (ТВ) и гладкомышечной клеткой (х42000). В терминалях много синаптических пузырьков (V), митохондрий (М). В мышечной клетке нити актина (А), плазматическая мембрана имеет впячивания (С). Хорошо видна синаптическая щель.
Пресинаптическая часть. Вблизи мышечного волокна аксон утрачивает миелиновую оболочку и дает несколько веточек, которые сверху покрыты уплощенными леммоцитами и базальной мембраной, переходящей с мышечного волокна. В терминалях аксона имеются митохондрии и синаптические пузырьки, содержащие ацетилхолин.
Синаптическая щель шириной около 50 нм располагается между плазмолеммои ветвлений аксона и мышечного волокна; она содержит материал базальной мембраны и отростки глиальных клеток, разделяющих соседние активные зоны одного окончания.
Постсинаптическая часть представлена мембраной мышечного волокна (сарколеммой), образующей многочисленные складки (вторичные синаптические щели), которые увеличивают общую площадь щели и заполнены материалом, являющимся продолжением базальной мембраны. В области нервно-мышечного окончания мышечное волокно не имеет исчерченности, содержит многочисленные митохондрии, цистерны грЭПС, рибосомы и скопления ядер.
Механизм передачи нервного импульса на мышечное волокно в нервно-мышечном синапсе сходен с таковым в химическом межнейронном синапсе. При деполяризации пресинаптической мембраны происходит выделение ацетилхолина в синаптическую щель; его связывание с холинорецепторами в постсинаптической мембране вызывает ее деполяризацию и последующее сокращение мышечного волокна. Медиатор отщепляется от рецептора и быстро разрушается ферментом ацетилхолинэстеразой, который содержится в синаптической щели.
Понимание механизмов передачи возбуждения в нервно-мышечных окончаниях имеет существенное клиническое значение. Действие некоторых ядов (например, кураре) обусловлено блокированием этой передачи, вызванным их прочным связыванием с холинорецепторами. Получены аналоги таких веществ (миорелаксанты), которые нашли применение в хирургии для расслабления мышц при полостных операциях, приводимых в условиях искусственного дыхания (поскольку нарушается деятельность дыхательных мышц). При миастении (myasthenia gravis) – заболевании, которое характеризуется прогрессирующей мышечной слабостью, в сарколемме снижается содержание рецепторов ацетилхолина (по-видимому, вследствие аутоиммунного процесса).
Двигательные нервные окончания в сердечной и гладких мышцах имеют вид варикозно расширенных (до 0.5-2 мкм) участков тонких (0.1-0.5 мкм) веточек аксонов, которые содержат многочисленные синаптические пузырьки и митохондрии. Обычно они отделены от мышечных клеток широкой (около 100 нм) щелью. Как правило, иннервированы лишь отдельные клетки, возбуждение с которых передается на соседние посредством щелевых соединений.
Секреторные нервные окончания представляют собой конечные участки тонких аксонных веточек. Одни из них, утрачивая оболочку из леммоцитов, проникают сквозь базальную мембрану и располагаются между секреторными клетками, заканчиваясь терминальными варикозными расширениями, содержащими пузырьки и митохондрии (гиполеммольный нейроэффекторный контакт). Другие не проникают сквозь базальную мембрану, образуя варикозные расширения вблизи секреторных клеток (тилеммалъный нейроэффекторный контакт). Секреторные нервные окончания оказывают на железистые клетки несколько видов воздействия: гидрокинетическое (мобилизация воды), протеокинетическое (секреция белка), синтетическое (усиление синтеза) и трофическое (поддержание нормальной структуры и функции).
Дата добавления: 2016-02-16 ; просмотров: 718 ; ЗАКАЗАТЬ НАПИСАНИЕ РАБОТЫ
1 М и Н холинореактивные системы
1. Эфферентная (или вегетативная) нервная система регулирует функцию всех внутренних органов и оказывает сложное влияние на основные биохимические процессы в организме животных. Поэтому лекарственные вещества, изменяющие состояние ее, имеют большое практическое значение для ветеринарии.
В эфферентных нервах различают центры, преганглионарные волокна, ганглии и постганглионарные волокна., Центры этих нервов и преганглионарные волокна реагируют примерно так же (и на те же вещества), как и соответствующие отделы центральной нервной системы. Практически это действие лекарственных веществ мало используется в ветеринарии, так как оно сильно изменяется в ганглиях и постганглионарных волокнах.
Наибольшее практическое значение имеют лекарственные вещества, действующие в области постганглионарных волокон и на ганглии; они и описаны в данном разделе учебника. Влияние постганглионарных эфферентных нервов в организме осуществляется при помощи химических веществ, называемых медиаторами (передатчиками возбуждения), образование и инактивация которых происходят под воздействием соответствующих ферментов.
По современным представлениям различают М- и Н-холинорецепторы (М-мускариночувствительные, Н-никотиночувствительные). Расположенные в постсинаптической мембране эффекторных органов в области окончаний постганглионарных холинергических нервов, холиноре-цепторы называются М-холинорецепторами или М-холинореактивными структурами, а структуры в области ганглиев (симпатических и парасимпатических), в скелетных мышцах, в хромафинных клетках мозгового слоя надпочечников и каротидных клубочков — Н-холинорецепто-
Адренорецепторы также бывают двух видов: Α. Β а-адренорецепторы находятся в гладкой мускулатуре кровеносных сосудов, матки и др. На адреналин и норадреналин они реагируют эффектами возбуждения. в-адрено-рецепторы расположены в мускулату скелетных мышц, а также сердца, линорецептора с ацетилхолином, бронхов и кишечника, они активизируются изопропилнорадреналином, а адреналин и норадреналин активность их понижают.
ВЕЩЕСТВА, ДЕЙСТВУЮЩИЕ НА М-ХОЛИНОРЕАКТИВНЫЕ СИСТЕМЫ (ХОЛИНЕРГИЧЕСКИЕ ВЕЩЕСТВА)
В эту группу входят препараты миметического действия (ацетил-холин, карбахолин), активизирующие постганглионарные холинергиче-ские системы, часто также называемые холиномиметическими (ареколин, пилокарпин, ацеклидин). В этой же группе целесообразно рассматривать антиацетилхолинэстеразные (физостигмин, прозерин, галантамин, ар-мин) и холиноблокирующие (атропин, скополамин, платифилин) вещества. Препараты первых трех видов действия —холиномиметические, активизирующие постганглионарные системы, и антихолинэстеразные— вызывают в основном сходные изменения в организме, типичные для активизации холинергической иннервации.
Резко активизируются секреторная и моторная функции пищеварительного тракта. В 2—8 раз повышается деятельность желез кишечника, несколько больше — желудка (в 3—12) и еще больше — слюнных желез (в 3—20 раз). Из животных разных видов наиболее резко секреция усиливается у травоядных и меньше — у плотоядных. Активизацию секреции желез не представляется возможным использовать для улучшения пищеварения из-за того, что процесс этот длится не дольше часа, сильно снижается активность ферментов сока и, кроме того, бывают резкие изменения функции многих других физиологических систем. Но это влияние очень важно для активизации выделения ядов железами пищеварительного тракта, а также для разжижения и ускорения продвижения содержимого пищеварительного тракта.
Вещества этого действия резко активизируют перистальтику кишечника, а также сокращения желудка и преджелудков.
Нам важно ваше мнение! Был ли полезен опубликованный материал? Да | Нет
Аксоны двигательных нейронов образуют эффекторные нервные окончания двух типов: секреторные (на клетках желез) и двигательные (в поперечно-полосатых и гладких мышцах). В скелетных мышцах это нейро-мышечный синапс или моторная бляшка. По строению как известный вам синапс, но постсинаптическая мембрана представлена участком плазмолеммы мышечного волокна. Один аксон, разветвляясь на конце, образует моторные бляшки сразу на целой группе мышечных волокон. В сердечной и гладкой мышечной тканях веточки аксонов образуют расширения – варикозы, в которых и находятся пузырьки с нейромедиатором. Как правило здесь иннервируются только некоторые клетки, а от них возбуждение передается на соседние с помощью нексусов.
Секреторные нервные окончания оканчиваются варикозными расширениями вблизи секреторных клеток и стимулируют синтез секретов либо процесс экзоцитоза.
8. Нейроглия.
Нейроглия – это группа вспомогательных клеток, которые обеспечивают деятельность нейронов. В ткани головного мозга их число в 5-10 раз больше, чем нейронов.
Выделяют микроглию и макроглию. Микроглия – это мелкие звездчатые клетки, которые образуются из моноцитов и являются специализированными макрофагами ЦНС. Они выполняют защитную, в том числе и антигенпредставляющую функцию. Выяснена ведущая роль этих клеток в поражении нервной системы при СПИДе. Они разносят вирус, а также инициируют усиленный апоптоз нейронов.
9. Характеристика и классификация макроглии.
Макроглия включает разные клетки, относящиеся к трем разновидностям: астроглия, олигодендроглия и эпендимная глия. Клетки эпендимной глии (эпендимоциты) Эпендимоциты.
Образуют выстилку полостей желудочков головного мозга и центрального канала спинного мозга. Они образуют пласт, соединенный межклеточными контактами и лежащий на базальной мембране, поэтому их одновременно относят и к эпителиям. Они разделяют нейроны и спинномозговую жидкость, образуя нейро-ликворный барьер (высокопроницаемый). А в области сосудистых сплетений входят в состав гемато-ликворного барьера (между кровью и спинномозговой жидкостью). Этот барьер включает: эндотелий сосудов, рвст, которая окружает сосуды, базальную мембрану эпендимоцитов и слой эпендимных клеток.
Олигодендроглия – разнообразные мелкие клетки с короткими и малочисленными отростками, которые окружают нейроны. В нервных узлах они охватывают тела нейронов, обеспечивая барьерную функцию. Другая группа образует оболочки вдоль отростков нейронов, вместе с ними формируя нервные волокна. В периферической н.с. их называют леммоциты или шванновские клетки, в ЦНС – олигодендроциты.
Астроглия представлена астроцитами – звездчатые клетки, похожие на нейроны. Протоплазматические астроциты характерны для серого вещества ЦНС имеют короткие толстые отростки, волокнистые - для белого вещества и имеют длинные отростки. Их функции – опорная (заполняют пространства между нейронами), метаболическая и регуляторная (поддерживают постоянным состав ионов и медиаторов), барьерная (входят в состав гемато-энцефалического барьера, который надежно изолирует нейроны от крови, не допуская иммунного конфликта). ГЭБ включает эндотелий капилляров и их базальную мембрану, и плотный футляр из отростков астроцитов, который покрывает сосуды.
10. Безмиелиновые и миелиновые нервные волокна. Образование и особенности строения.
Нервные волокна – отростки нейронов (их называют осевыми цилиндрами), которые покрыты оболочкой из глиальных клеток. Различают миелиновые и безмиелиновые нервные волокна.
Безмиелиновые волокна образуются при погружении осевого цилиндра в углубления леммоцитов, которые лежат цепочкой вдоль всего аксона. Леммоциты прогибаются настолько, что их мембраны соприкасаются над осевым цилиндром. Эта дубликатура называется мезаксон. Если в цепочку леммоцитов погружается сразу несколько аксонов, такое волокно называют кабельным.
Миелиновые нервные волокна. Образуются с участием шванновских клеток, которые сначала формируют над осевым цилиндром мезаксон, а затем начинают многократно закручиваться. Цитоплазма вместе с ядром оттесняется наружу, образуя слой, который называют нейролеммой. Под ней лежит толстый слой тесно прилежащих сдвоенных мембран, который называют миелином. В определенных участках между витками остаются небольшие прослойки – миелиновые насечки. Поскольку шванновские клетки. Аксон длинный и шванновских клеток вдоль него много. На границах двух соседних клеток миелиновая оболочка исчезает. Эти участки называют узловые перехваты Ранвье.
В ЦНС миелиновая оболочка образуется несколько по-иному.
Миелиновые волокна проводят нервный импульс в десятки раз быстрее, чем безмиелиновые.
Большая советская энциклопедия. — М.: Советская энциклопедия . 1969—1978 .
- Эффель Жан
- Эффузивные горные породы
эфферентные нервные волокна — (лат. efferens (eiicrentis) выносящий) центробежные нервные волокна нервные волокна, по которым возбуждение передается от центральной нервной системы (от клетки) к тканям ср. афферентные нервные волокна). Новый словарь иностранных слов. by EdwART … Словарь иностранных слов русского языка
ЭФФЕРЕНТНЫЕ НЕРВНЫЕ ВОЛОКНА — [от лат. effero выносить; efferens (efferentis) выносящий] центробежные двигательные волокна, по которым возбуждение передается от ЦНС ко всем органам тела (ср. афферентные нервные волокна) … Психомоторика: cловарь-справочник
Нервные волокна — Нервные волокна отростки нейронов, покрытые глиальными оболочками. В различных отделах нервной системы оболочки нервных волокон значительно отличаются по своему строению, что лежит в основе деления всех волокон на миелиновые и безмиелиновые … Википедия
афферентные нервные волокна — (лат. afferens (afferentis) приносящий) центростремительные нервные волокна нервные волокна (отростки нервных клеток), по которым возбуждение передается от тканей к центральной нервной системе ср. эфферентные нервные волокна). Новый словарь… … Словарь иностранных слов русского языка
Двигательные нервные волокна — или Эфферентные нервные волокна, отростки нервных клеток, по которым импульсы идут от центральной нервной системы к исполнительным органам мышцам (такие волокна называют моторными), железам и др. Ср. Чувствительные нервные волокна … Большая советская энциклопедия
ПРОЕКЦИОННЫЕ НЕРВНЫЕ ВОЛОКНА — [от лат. projectio бросание вперед] волокна, связывающие полушария головного мозга с нижележащими его отделами стволом и спинным мозгом. По П. н. в. по ступает афферентная (чувствительная) и эфферентная (двигательная) информация (см. Афферентные… … Психомоторика: cловарь-справочник
Центробежные нервные волокна — эфферентные, моторные нервные волокна, отростки нервных клеток (Аксоны) вместе с их оболочками, передающие возбуждение от центральной нервной системы к рабочим органам (мышцам, железам). Различают черепномозговые, спинномозговые и др. Ц.… … Большая советская энциклопедия
Нервные отростки — Нервные волокна отростки нейронов, покрытые глиальными оболочками. В различных отделах нервной системы оболочки нервных волокон значительно отличаются по своему строению, что лежит в основе деления всех волокон на миелиновые и безмиелиновые. Те… … Википедия
Нервные клетки — Не следует путать с нейтроном. Пирамидальные ячейки нейронов в коре головного мозга мыши Нейрон (нервная клетка) – это структурно функциональная единица нервной системы. Эта клетка имеет сложное строение, высоко специализирована и по структуре… … Википедия
волокна парасимпатические постганглионарные — (neurofibrae postganglionares parasympathicae) эфферентные нервные В., идущие от клеток парасимпатических узлов, интрамуральных или параорганных сплетений … Большой медицинский словарь
Афферентные (чувствительные)нервные окончания (рис. 1.5.5, 1.5.6).
Афферентные нервные окончания представляют собой концевые аппараты дендритов чувствительных нейронов, повсеместно располагающихся во всех органах человека и дающие информацию центральной нервной системе об их состоянии. Воспринимают они раздражения, исходящие и из внешней среды, преобразуя их в нервный импульс. Механизм возникновения нервного импульса характеризуется уже описанными явлениями поляризации и деполяризации цитоплазматической мембраны отростка нервной клетки.
Существует ряд классификаций афферентных окончаний — в зависимости от специфичности раздражения (хеморецепторы, бароре-
Глава 1. КЛЕТКА И ТКАНИ
 |
це Мейснера, терморецепторы колбы Краузе, тельца Руффини и др.).
Разнообразно строение рецепторов мышечной ткани, часть которых обнаруживается в наружных мышцах глаза. В этой связи на них мы остановимся более подробно. Наиболее распространенным рецептором мышечной ткани является нервно-мышечное веретено (рис. 1.5.6). Это образование регистрирует растяжение волокон поперечно-полосатых мышц. Представляют они собой сложные инкапсулированные нервные окончания, обладающие как чувствительной, так и двигательной иннервацией. Число веретен в мышце зависит от ее функции и тем выше, чем более точными движениями она обладает. Нервно-мышечное веретено располагается вдоль мышечных волокон. Веретено покрыто тонкой соединительнотканной капсулой (продолжение периневрия), внутри которой находятся тонкие поперечнополосатые интрафу-зальные мышечные волокна двух видов:
— волокна с ядерной сумкой — в расши
ренной центральной части которых содержатся
скопления ядер (1—4 — волокна/веретено);
— волокна с ядерной цепочкой — более тон
кие с расположением ядер в виде цепочки в
центральной части (до 10 волокон/веретено).
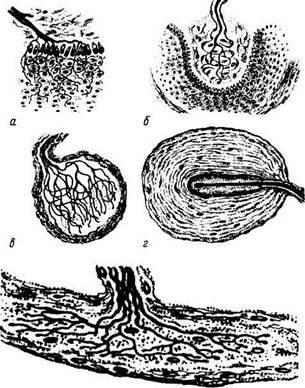
Рис. 1.5.5. Особенности строения различных рецеп-торных окончаний:
а — свободные нервные окончания; б— тельце Мейснера; в — колба Краузе; г — тельце Фатер—Пачини; д — тельце Руффини
цепторы, механорецепторы, терморецепторы и др.), от особенностей строения (свободные нервные окончания и несвободные).
Обонятельные, вкусовые, зрительные и слуховые рецепторы, а также рецепторы, воспринимающие движение частей тела относительно направления силы тяжести, называют специальными органами чувств. В последующих главах этой книги мы подробно остановимся только на зрительных рецепторах.
Рецепторы разнообразны по форме, строению и функциям. В данном разделе нашей задачей не является подробное описание различных рецепторов. Упомянем лишь о некоторых из них в разрезе описания основных принципов строения. При этом необходимо указать на различия свободных и несвободных нервных окончаний. Первые характеризуются тем, что они состоят только из ветвления осевых цилиндров нервного волокна и клетки глии. При этом они контактируют разветвлениями осевого цилиндра с клетками, возбуждающими их (рецепторы эпителиальных тканей). Несвободные нервные окончания отличаются тем, что в своем составе они содержат все компоненты нервного волокна. Если они покрыты соединительнотканной капсулой, они называются инкапсулированными (тельце Фатер—Пачини, осязательное тель-
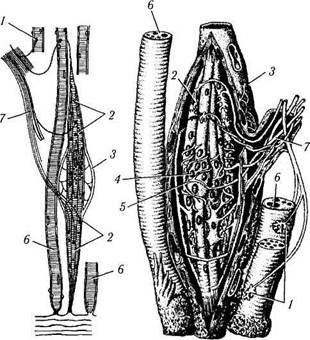
Рис. 1.5.6. Строение нервно-мышечного веретена:
а—моторная иннервация интрафузальных и экстрафузальных мышечных волокон; б — спиральные афферентные нервные окончания вокруг интрафузальных мышечных волокон в области ядерных сумок (/ — нервно-мышечные эффекторные окончания экстрафузальных мышечных волокон; 2 — моторные бляшки интрафузальных мышечных волокон; 3 — соединительнотканная капсула; 4 — ядерная сумка; 5 — чувствительные кольцеспираль-ные нервные окончания вокруг ядерных сумок; 6 — скелетные мышечные волокна; 7 — нерв)
Строение периферической нервной системы.
Двигательные нервные волокна — тонкие, образуют мелкие нервно-мышечные синапсы по краям интрафузальных волокон, обеспечивая их тонус.
Рецепторами растяжения мышцы являются также нервно-сухожильные веретена (сухожильные органы Гольджи). Это веретеновид-ные инкапсулированные структуры длиной около 0,5—1,0 мм. Располагаются они в области соединения волокон поперечнополосатых мышц с коллагеновыми волокнами сухожилий. Каждое веретено образовано капсулой из плоских фиброцитов (продолжение периневрия), которая охватывает группу сухожильных пучков, оплетенных многочисленными терминальными веточками нервных волокон, частично покрытых леммоцитами. Возбуждение рецепторов возникает при растяжении сухожилия во время мышечного сокращения.
Эфферентные нервные окончания несут информацию от центральной нервной системы к исполнительному органу. Это окончания нервных волокон на мышечных клетках, железах и др. Более подробное их описание будет приведено в соответствующих разделах. Здесь мы подробно остановимся лишь на нервно-мышечном синапсе (моторная бляшка). Моторная бляшка располагается на волокнах поперечнополосатых мышц. Состоит она из концевого ветвления аксона, образующего пресинапти-ческую часть, специализированного участка на мышечном волокне, соответствующего постси-наптической части, и разделяющей их синапти-ческой щели. В крупных мышцах один аксон иннервирует большое количество мышечных волокон, а в небольших мышцах (наружные мышцы глаза) каждое мышечное волокно или их небольшая группа иннервируется одним аксоном. Один мотонейрон в совокупности с ин-нервируемыми им мышечными волокнами образует двигательную единицу.
Пресинаптическая часть формируется следующим образом. Вблизи мышечного волокна аксон утрачивает миелиновую оболочку и дает несколько веточек, которые сверху покрыты уплощенными леммоцитами и базальной мембраной, переходящей с мышечного волокна. В терминалах аксона имеются митохондрии и синапти-ческие пузырьки, содержащие ацетилхолин.
Синаптическая щель имеет ширину 50 нм. Располагается она между плазмолеммой ветвлений аксона и мышечного волокна. Содержит она материал базальной мембраны и отростки глиальных клеток, разделяющих соседние активные зоны одного окончания.
Постсинаптическая часть представлена мембраной мышечного волокна (сарколеммой), об-
разующей многочисленные складки (вторичные синаптические щели). Эти складки увеличивают общую площадь щели и заполнены материалом, являющимся продолжением базальной мембраны. В области нервно-мышечного окончания мышечное волокно не имеет исчерченности, содержит многочисленные митохондрии, цистерны шероховатого эндоплазматического ретику-лума и скопление ядер.
Механизм передачи нервного импульса на мышечное волокно сходен с таковым в химическом межнейронном синапсе. При деполяризации пресинаптической мембраны происходит выделение ацетилхолина в синаптическую щель. Связывание ацетилхолина с холинорецеп-торами в постсинаптической мембране вызывает ее деполяризацию и последующее сокращение мышечного волокна. Медиатор отщепляется от рецептора и быстро разрушается ацетил-холинэстеразой.
Читайте также: