
Основные функциональные блоки нервной системы
А.Р. Лурия (1973) обобщил имеющиеся данные об анатомических структурах головного мозга и, показав их физиологическую значимость для развития психических процессов человека, выделил три основные функциональные блока, осуществляющие психическую деятельность. Каждый блок имеет иерархическое строение и состоит из надстроенных друг над Другом уровней нервной системы.
Первый блок — блок регуляции тонуса и бодрствования. Для обеспечения полноценного протекания психических про-
цессов человек должен находиться в состоянии бодрствования. Для их поддержания необходим определенный тонус коры головного мозга, обеспечивающий прием и переработку информации, возникновение в памяти определенных ассоциаций, планирование деятельности. Аппаратом, обеспечивающим и регулирующим тонус коры головного мозга, является ретикулярная (сетевидная) формация. Располагаясь в стволе мозга и поднимаясь вверх до межуточного мозга, она выполняет функцию объединения многих образований, активизируя импульсы, идущие вверх и вниз. Восходящая ретикулярная система выполняет решающую роль в регуляции активности коры. Нисходящая ретикулярная формация контролирует и регулирует структуры среднего мозга, гипоталамуса и мозгового ствола, подчиняя их регулирующей функции коры. Таким образом, ретикулярная формация обеспечивает регуляцию тонуса коры и состояние бодрствования. Однако активизирующая функция ретикулярной формации является неспецифической, отличаясь этим от специфических (сенсорных и моторных) функций систем мозговой коры. Объединяя стволовые образования, средний и межуточный мозг, ретикулярная формация поддерживает регуляцию обменных процессов, обеспечивающих гомеостаз (внутреннее равновесие организма).
Активизировать кору может не только ретикулярная формация, но и раздражения, приходящие от органов чувств. Человек живет в условиях постоянно меняющейся среды, требующей от него обостренного состояния бодрствования, мобилизации организма. И.П. Павлов в свое время называл такую мобилизацию ориентировочным рефлексом, являющимся основой познавательной деятельности.
Таким образом, первый блок объединяет ствол мозга с ядрами черепно-мозговых нервов и системой блуждающего нерва, ретикулярную формацию, средний и межуточный мозг, а также подкорковые образования.
Второй блок — блок приема, переработки и хранения информации располагается в наружных отделах новой коры и включают зрительную (затылочную), слуховую (височную), общечувствительную (теменную) области. Аппараты этого блока приспособлены к приему внешних раздражений, приходящих в головной мозг от периферических рецепторов, к дроблению их на огромное количество компонентов (к анализу сигналов) и к комбинированию их в нужные динамические
функциональные структуры (к синтезу раздражителей). Этот блок головного мозга обладает высокой организацией клеточных структур и их связей, чтобы принимать зрительную, слуховую и общечувствительную информацию. Он включает отделы коры, воспринимающие обонятельные и вкусовые раздражения. Основную функцию принимают на себя первичные и вторичные поля коры головного мозга, центральный аппарат модально-специфического анализатора, построенного по единому принципу иерархической организации. Сложную интеграцию задних отделов коры головного мозга осуществляют третичные поля. По мнению А.Р. Лурия, деятельность третичных зон необходима не только для успешного синтеза наглядной информации, но и для перехода от уровня непосредственного наглядного синтеза к уровню символических процессов, необходимых для оперирования со значениями слов, сложными грамматическими и логическими структурами, системами чисел и отвлеченными соотношениями, участие которых необходимо для превращения наглядного восприятия в отвлеченное мышление, опосредованное внутренними схемами.
Информация, достигающая нервных клеток первичных полей, обрабатывается, дифференцируется, попадает во вторичные поля гностической области, где сохраняется след приходящих сюда раздражений. И.П. Павлов считал, что память — это следы раздражений, достигающих коры головного мозга. Большое значение имеет структура ассоциативных нейронов с короткими аксонами, составляющими вторичные поля, которые обеспечивают объединение различных комбинаций раздражений в блоки, осуществляя синтетическую функцию. В детском возрасте основное значение имеет последовательность включения и надстройки первичных, вторичных и третичных полей. Поражение первичных и вторичных полей затрудняет формирование третичных полей, что сказывается на недоразвитии высоких уровней, задерживает формирование высших психических функций.
Третичные зоны (зоны перекрытия) корковых отделов различных анализаторов имеются только у человека. Они принимают участие в превращении наглядного восприятия в отвлеченное мышление, опосредованное внутренними схемами, и необходимы для сохранения в памяти организованного опыта (А.Р. Лурия, Л.С. Выготский). У взрослого человека, психические функции которого полностью сформировались,
ведущее значение приобретают третичные зоны коры, управляющие подчиненными им вторичными зонами. Принцип латерализации высших психических функций в коре головного мозга актуален только на уровне вторичных и третичных зон, которые играют основную роль в функциональной организации доходящей до коры информации, осуществляемой у человека с помощью речи. Таким образом, второй блок обеспечивает наиболее сложные формы работы мозга, лежащие в основе наиболее высоких видов познавательной деятельности человека, генетически связанных с трудом, а структурно — с участием речи в организации психических процессов.
Третичные зоны коры располагаются в префронтальных отделах мозга, имеют богатую систему связей со всеми отделами коры и с нижележащими отделами мозга, особенно с ретикулярной формацией, которая заряжает и тонизирует все отделы мозга. Связи лобной коры двухсторонние. Кора оказывает регулирующее влияние на образования ретикулярной формации, придавая им дифференцированный характер. Таким образом, префронтальные отделы коры играют важную роль в регуляции состояний активности, приводя их в соответствие с формулируемыми с помощью речи намерениями и замыслами (А.Р. Лурия). Отличительная черта процессов регуляции сознательной деятельности у человека состоит в том, что она совершается при ближайшем участии речи. Именно префронтальные отделы коры обеспечивают сложнейшие формы про-
граммирования, регуляции и контроля сознательной деятельности человека.
Обобщая полученные данные о функционировании трех блоков, необходимо подчеркнуть их функциональное единство при формировании сложной психической деятельности человека (рис. 43).
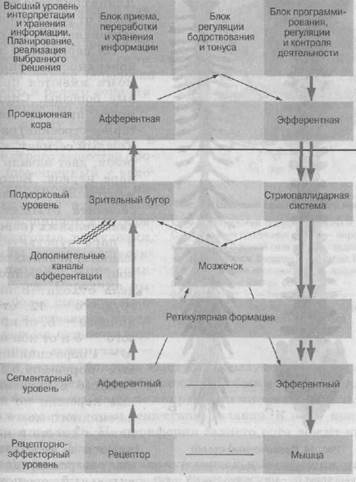
Рис. 43. Общий принцип обработки информации в нервной системе
| | | следующая лекция ==> | |
| Ретикулярная формация | | | Строение и функции спинного мозга |
Дата добавления: 2019-07-26 ; просмотров: 308 ; ЗАКАЗАТЬ НАПИСАНИЕ РАБОТЫ
Именно кора головного мозга осуществляет анализ и синтез поступающих через анализаторы раздражений, поскольку там происходит конечное замыкание всех нервных связей. Однако следует подчеркнуть, что психические процессы человека являются сложными функциональными системами и они не локализованы в узких, ограниченных участках мозга. Они осуществляются при участии многих совместно работающих мозговых аппаратов. А. Р. Лурия выделяет три основных функциональных блока мозга, участие которых необходимо для осуществления любого вида психической деятельности.
Первый — "энергетический блок", или блок регуляции тонуса коры и состояния бодрствования. Анатомически он представляет собой образование в стволе мозга по типу нервной сети, в которую вкраплены тела нервных клеток, соединяющиеся между собой короткими отростками (ретикулярная формация). Одни из волокон ретикулярной формации направляются вверх и оканчиваются в вышележащих нервных образованиях, включая кору мозга (восходящая ретикулярная система). Другие имеют обратное направление: начинаются в коре и направляются к нижележащим структурам среднего мозга, гипоталамуса и мозгового ствола (нисходящая ретикулярная система).
Нервная система всегда находится в состоянии определенной активности, которая, как уже говорилось, опосредована работой ретикулярной формации.
Первый источник активации — обменные процессы в организме, лежащие в основе гомеостаза (внутреннего равновесия организма), и инстинкты. Процессы обмена веществ регулируются главным образом аппаратами гипоталамуса. Инстинктивное поведение (пищевая, половая активация) является функцией более высоко расположенных образований мезэнцефальной, диэнцефальной и лимбической систем.
Второй источник активации связан с поступлением информации от внешней среды. Здесь активация проявляется в виде ориентировочного рефлекса. Человек, лишенный обычного притока информации извне (эксперименты по "сенсорной депривации"), впадает в дремотное состояние, и у него могут возникать галлюцинации и другие психические нарушения.
Третий источник активации обусловлен намерениями, планами, перспективами и программами, которые формируются в процессе сознательной деятельности человека.
Ретикулярная формация имеет обширные связи, прежде всего с лобными отделами коры мозга. Нисходящие аппараты ретикулярной формации облегчают и обеспечивают проведение корковых влияний (как активизирующих, так и тормозных) на нижележащие отделы нервной системы. Аппараты первого функционального блока мозга не только тонизируют кору, но и сами испытывают па себе се регулирующее влияние.
Второй блок — "блок приема, переработки и хранения информации" состоит из трех подблоков, расположенных в конвекситальных (наружных) отделах коры затылочной (зрение), височной (слух и вестибулярная информация) и теменной (общая чувствительность) долей мозга. Сюда включаются и центральные аппараты вкуса и обоняния, но у человека они занимают в коре головного мозга незначительное место.
Каждый подблок имеет иерархическое строение. Условно в них выделяют первичные, вторичные н третичные отделы. Первые дробят воспринимаемый образ мира — слуховой, зрительный, осязательный — на мельчайшие признаки: округлость и угловатость, высоту и звонкость, яркость и контрастность. Вторые синтезируют из этих признаков целые образы. Третьи объединяют информацию, полученную от разных подблоков, т.е. зрения, слуха, обоняния, осязания.
Третий блок — "блок программирования, регуляции и контроля" расположен преимущественно в лобных долях мозга. Первичная двигательная кора (четвертое поле Бродмана) не может работать изолированно. Подготовка программ движения и передача их на пирамидные клетки осуществляется в аппаратах вторичных зон двигательной коры, которые располагаются в верхних слоях коры премоторных отделов лобной области. Третичные зоны лобной коры играют решающую роль в формировании намерений и программ, в регуляции и контроле наиболее сложных форм поведения. Человек, у которого этот блок нарушен, лишается возможности поэтапно организовывать свое поведение, не может перейти от одной операции к другой. В связи с этим личность такого человека как бы "распадается".
Основные подходы к классификации строения нервной системы
1. Центральная - периферическая
2. Модульный Подход (по Г. Шеперду)
Строение нервной системы можно представить как совокупность отдельных блоков (модулей), каждый из которых выполняет свою функцию.
Микросети. Нижний уровень организации нейронных сетей. Его образуют отдельные синапсы с их пре- и постсинаптическими структурами. Синапс – это не только простейший соединительный элемент в сети, но и сложная самостоятельная функциональная единица, в которой действуют регуляторные механизмы, зависимые от времени и интенсивности функционирования.
Локальные сети (коллатерали и интернейроны). Здесь возможно выполнение логических операций, которые выполняет и компьютер.
Локальные модули. Это колонки, клубочки, ядра, ганглии.
Поля и доли коры. Полей, по разным классификациям, 40-50. Долей – 4 (лобная, теменная, затылочная, височная).
3. Атомарный подход
Строение нервной системы можно представить как совокупность слоев, аналогично строению земли.
Три отдела (функциональных блока) нервной системы и психика.
В Н.С. существует 3 отдела: Вегетативная нервная система, Двигательная нервная система и Центральная нервная система.
Вегетативная нервная система. В нее входит лимбическая кора, орбитальная кора, гиппокамп, премоторная кора, миндалевидный комплекс. Делится на симпатическую и парасимпатическую.
Симпатическая состоит из волокон, клетки тела которых лежат в боковых столбах серого вещества спинного мозга. Их аксоны выходят через передние корешки спинномозговых нервов вместе с двигательными волокнами к скелетным мышцам, отделяются и образуют вегетативную ветвь спинномозгового нерва, идущего к симпатическому ганглию (парные). Всего 18 пар: от шеи до обл. живота.
Парасимпатическая состоит из волокон, начинающихся в главном мозге (средний, продолговатый) и выходит в составе 3, 7, 9 и10 черепно-мозговых нервов.
Преобладание симпатической нервной системы – низкое АД, низкая частота пульса и дыхания. Низкая температура конечностей. Видят ситуацию в целом, плохой аппетит, хорошая память, письменная речь.
Преобладание парасимпатической нервной системы – краснеют, подвержены укачиванию. Им легче написать, чем сказать. Стресс, алкоголь – депрессия.
Двигательная нервная система состоит из пирамидальной системы и экстрапирамидальной.
Пирамидальная система организует целенаправленные, точные движения. Состоит: моторная кора, внутренняя капсула, мост, латеральные кортикоспинальные волокна.
Экстрапирамидальная более древняя. Регулирует мышечный тонус. Состоит: поясная и моторная кора, базальные ганглии, часть таламуса и итраламинарные ядра, субталамическое ядро, мозжечок, красное ядро, черная субстанция, ретикулярная формация.
В Центральной нервной системе 3 отдела: проекционные зоны, вторичные и третичные.
В проекционных зонах производится первичная обработка поступающей информации. Поля 3, 4, 17, 41. Связаны с периферическими отделами нервной системы.
Во вторичных осуществляется тончайший анализ раздражений и их упорядочивание. Поля 1, 2, 43, 6, 18, 19, 42.
Третичные поля осуществляют наиболее сложный анализ, синтез деятельности нескольких анализаторов. Поля 5, 7, 39, 40- теменные, 21, 32, 37, 38- височные, 8-12, 44-47 - лобные.
Вопросы к экзамену по учебной дисциплине
для дистантного обучения
и для ленивых студентов различных факультетов и специальностей
Множество ответов на данные вопросы можно найти в этом электронном учебнике:
Часть 2. Физиология ВНД
1. Предмет и задачи физиологии высшей нервной деятельности. Определение ВНД. Методы исследования ВНД. Основные ритмы ЭЭГ и их значение.
2. История развития взглядов на высшую нервную деятельность. Предпосылки возникновения учения И. П. Павлова о ВНД. Рефлекторная теория И. М. Сеченова.
3. Основные принципы рефлекторной теории.
4. Основные функциональные блоки нервной системы, обеспечивающие ВНД. Функционально-блочная схема работы мозга.
6. Модулирующие системы в функциональной организации мозга. Формы активации и источники активации мозга. Пути неспецифического возбуждения для коры головного мозга.
7. Основы функциональной организации двигательных систем мозга. Особенности двигательного анализатора.
8. Третичные (ассоциативные) зоны коры как особый функциональный блок. Особенности их функционирования.
9. Врожденная деятельность организма. Безусловные рефлексы и их классификация. Инстинкты и инстинктивное поведение.
10. Функциональное состояние в структуре поведения. Уровни проявления функционального состояния. Шкалы функциональных состояний.
11. Дуги безусловного и условного рефлекса по Э. А. Асратяну.
12. Концепция нейронной организации рефлекторной дуги по И. П. Павлову и Е. Н. Соколову.
13. Сон. Структура и теории сна. Электрофизиологические параметры. Засыпание.
14. Стресс. Значение, механизмы. Схема развития стрессовой реакции.
15. Концепция драйва и драйв-рефлексы по Ю. Конорскому. Импринтинг: необходимые условия и особенности.
16. Условные рефлексы как эффектзависимое обучение. Условия выработки условных рефлексов.
17. Механизмы образования условного рефлекса. Функциональные основы замыкания временной связи. Концепция Э. А. Асратяна.
18. Динамика условнорефлекторной деятельности.
19. Механизмы торможения условных рефлексов. Виды условного торможения.
20. Виды безусловного торможения условных рефлексов, их характеристика.
21. Доминанта. Динамика существования доминанты.
22. Нейрофизиологические особенности памяти. Временная организация памяти. Структурно-функциональные основы памяти и обучения.
23. Клеточные и молекулярные механизмы памяти и обучения. Роль пластичности синапсов.
24. Структура поведенческого акта. Функциональная система П. К. Анохина. Стадии поведенческого акта.
25. Потребности, их детекция. Детерминанты потребности. Классификация потребностей.
26. Мотивация. Биологическая мотивация. Общие свойства различных видов мотиваций. Мотивация как доминанта.
27. Нейроанатомия и нейрохимия мотиваций.
28. Эмоции. Функции эмоций. Физиологическое выражение эмоций.
29. Нейроанатомиия и нейрохимия эмоций.
30. Особенности ВНД человека. Слово как сигнал сигналов. Речь и её функции. Развитие речи у ребёнка.
31. Первая и вторая сигнальные системы, их взаимодействие. Речевые функции полушарий.
32. Мозг, мышление и сознание.
33. Теория И. П. Павлова о типах ВНД. Общие и человеческие типы ВНД.
34. Индивидуальные различия в высшей нервной деятельности человека. Донервная теория индивидуальности (Гиппократ, Э. Кречмер и др. ).
35. Ориентировочный рефлекс. Его структура и нейронные механизмы. Особенности ориентировочного рефлекса.
36. Определение биологической значимости раздражителя.
37. Асимметрия мозга и межполушарные взаимодействия.
38. Имитационное моделирование нейросети, динамические модели рефлекторных дуг.
1. Основные понятия и принципы функционирования сенсорных систем.
2. Общие принципы работы сенсорных систем. Органы чувств, анализаторы, сенсорные системы.
3. Виды анализаторов. Сенсорные модальности. Свойства анализаторов.
4. Классификация рецепторов и их виды.
5. Механизм трансформации раздражения в возбуждение (трансдукция). Рецепторный и генераторный потенциал. Общая схема рецепторного акта.
6. Низшие нервные центры, их функции и значение для сенсорных систем.
7. Рецепторный отдел зрительной сенсорной системы. Его строение и функционирование.
8. Обработка зрительной информации в таламусе. Рецептивные поля нейронов НКТ.
9. Корковый отдел зрительной сенсорной системы. Анализ зрительных раздражений нейронами зрительной коры. Рецептивные поля корковых нейронов.
10. Колончатая организация коры.
11. Рецепторный отдел слуховой сенсорной системы. Механизмы рецепции звука, трансдукция.
12. Проводниковый отдел слухового анализатора. Движение возбуждения к коре.
13. Восприятие звука.
14. Вестибулярный анализатор. Центральные механизмы чувства равновесия.
15. Кинестетический анализатор. Качества проприоцепции.
16. Тактильный анализатор.
17. Вкусовой анализатор.
18. Обонятельная сенсорная система.
19. Болевая сенсорная система. Нейрофизиологическая основа боли.
20. Антиноцицептивная система.
21. Пути сенсорного возбуждения в нервной системе.
Пользователей онлайн: 0.

Приветствую вас на своем сайте, здесь вы можете найти много полезной информации (или что-то типа того)
САЗОНОВ Вячеслав Фёдорович
доцент кафедры биологии Рязанского государственного университета имени С.А. Есенина, кандидат биологических наук. Преподаватель вуза с 1978 года.
Your browser does not support canvas.
- 13й бодхисаттва
- ВНД
- КСЕ
- антистресс
- возбуждение
- возрастная
- вопросы
- кинезиология
- мат.статистика
- мембрана
- нервная система
- основа
- понятия
- ресурсы
- сексология
- сенсорика
- статобработка
- стресс
- студентам
- физиология
Реклама
Поиск
Притча наудачу:
На сайте введена регистрация через социальные сети, если вы хотите оставлять комментарии без потверждения, пожалуйста, воспользуйтесь именно этим типом аутентификации.
Поддержка сайта
Поддержите сайт материально!
Это очень поможет.
Перечислите любую сумму на карту Сбербанка номер:
Функциональная организация нервной системы
В основе работы самых разных органов чувств лежат те же три рецепторных принципа, известных для растений и простейших: химическая, физическая и электромагнитная чувствительность мембран. Из этих трёх источников внешних сигналов организм животных создал огромное разнообразие органов чувств. Механочувствительность реализуется в виде слуха, органов боковой линии, грави- или терморецептора. Химическая чувствительность может быть представлена дистантным обонянием или контактным органом вкуса, осморецептором или рецептором парциального давления кислорода. Чувствительность к электромагнитным волнам обусловлена рецепторами внешних или собственных полей, светочувствительностью либо способностью воспринимать магнитные поля планеты и Солнца.
Разнообразие сигналов среды крайне велико, но большинство их быстротечны. Они имеют ценность только непродолжительное время. Следовательно, такие сложные сигналы надо сначала воспринять, а затем быстро использовать полученную информацию. Сложная и быстро работающая рецепторная система нужна только организму с высоким уровнем активности, т. е. метаболизма. Для использования информации её надо сравнивать и реагировать на приоритетные (опасные, половые, пищевые) воздействия. Это можно осуществить в таком устройстве, в
котором были бы представлены все используемые органы чувств. Таким образом, нужен некий центр сравнения получаемой информации. Если даже никакой индивидуальной памяти и опыта не накапливать, то всё равно надо хотя бы сравнивать сигналы различной природы. Затем было бы хорошо выбрать главный раздражитель и отвечать в первую очередь на него. Для этого нужно небольшое скопление тел нервных клеток, которые получают информацию от разных источников.
Различные органы чувств и центр сравнения их информации бесполезны без системы реализации полученных сигналов, поэтому нервная система неотделима от эффекторных структур организма. Организм может ответить на внешний сигнал при помощи сокращения или расслабления мышц различных типов, активизации или подавления выброса биохимических сигналов (ферментов, гормонов). При сравнении сигналов из внешней среды должен присутствовать центр управления эффекторными органами. Вполне допустимо объединение этих функций в одном центре. В предельно упрощённом виде нервная система будет представлять собой три относительно разделённых блока (см. рис. I-4, б). Рецепторный блок должен быть приближен к источникам информационных сигналов. Внешние сенсорные системы должны располагаться на границе тела и внешней среды — в покровах. Блок сравнения, как правило, равноудалён от рецепторов и эффекторов. Поскольку этот блок становится важнейшим интегративным центром, он должен быть механически защищён.
Эта идеализированная схема не имеет ничего общего с реальной нервной системой свободноживущих кишечнополостных. Их диффузная нервная сеть распределена почти равномерно по всему телу. Исключение представляют небольшое скопление клеток у подошвы и окологлоточное нервное кольцо.
Такая функционально минимизированная нервная система позволяет быстро, но довольно неспецифически реагировать на внешнее воздействие. Примером могут быть кишечнополостные. Обычная пресноводная гидра мгновенно сжимается, если качнуть лист, на котором она сидит, прикоснуться к ней щетинкой или вызвать движение воды. Сжавшись в серый комочек, гидра становится менее уязвимой.
В таком виде нервная система эффективна, но крайне неспецифична. Она по функциям отличается от растений только тем, что распределена по всему организму. Это обеспечивает быструю согласованную реакцию всего организма, но ответ не дифференцирован. Он столь же неспецифичен, как и реакция простейших на внешние воздействия. Это сходство долго было
основой для идентификации поведения многоклеточных животных и простейших.
Самое первое и простое преимущество, которое даёт нервная система, — это способность многоклеточного организма реагировать на внешние воздействия со скоростью простейших. При этом ответ, как правило, весьма однотипен. Отсутствие специфичности и контроля результатов реакции только подчёркивает наиболее древнее свойство нервной системы — способность распространять информацию о контакте с внешним миром на весь многоклеточный организм.
Таким образом, нервная система нужна тогда, когда многоклеточному организму необходимо реагировать на внешние воздействия как целому. Реакция может быть как угодно неспецифична, но должна быть генерализована, т. е. охватывать весь организм. Эта способность даёт большое преимущество в биологической эволюции. Примеряя данное эволюционное приобретение на себя, легко понять величие события. Если во время урагана ты будешь гордо стоять во весь рост, то тебя скоро заменит памятник.
Если сгруппироваться, как гидра, есть шанс не планировать ритуальные расходы. В связи с этим неспецифическая, но генерализованная реакция организма стала эпохальным началом быстрых адаптаций нейрогенной природы (см. рис. I-4, а).
Уязвимым звеном однотипных реакций организма на все типы воздействий является их неспецифичность. Она очень хороша, когда ничего нет, но при системе интеграции многоклеточного организма её специализации на различные чувствительные компоненты уже недостаточно. Необходимость эволюционной дифференцировки нейронов на несколько чувствительных типов диктуется самой способностью клеток воспринимать механические, химические и электромагнитные сигналы.
Такое упреждение событий (даже в сочетании с неспецифической реакцией организма) стало существенным шагом в быстрых адаптациях многоклеточного организма. Это ещё один мотив для возникновения нервной системы. Она нужна для быстрого ответа многоклеточного организма на внешние воздействия с дифференциацией источника воздействия и упреждением грядущих событий. Организмы, не имеющие нервной системы, о такой адаптивности не могут и мечтать. Однако сразу возникает двойная проблема. Во- первых, сигналы от различных органов чувств должны прийти в одно и то же место, где их можно было бы сравнить, но не просто сравнить, а выбрать самый главный на данный момент. Именно доминирующее возбуждение и будет основным сигналом к действию. Во-вторых, их надо как-то сделать сравнимыми. Одни из них идут от фоторецепторов, другие — от хеморецепторов, а третьи — от рецепторов электромагнитного излучения. Сопоставимость столь разнородных сигналов достигается однотипностью их кодировки. Электрохимический сигнал нейронов, изменяющийся по частоте, амплитуде, модуляции, интенсивности, повторяемости и некоторым другим параметрам, оказался вполне достаточным для сравнимости сигналов из разных органов чувств.
Следующие приобретения для эффективной скоротечной адаптивности вновь связаны с чувствительностью, но уже совершенно другого рода. С одной стороны, это внутренние рецепторы. Активный организм должен знать, чего ему хочется или в чём он остро нуждается. Любому организму обычно хочется поесть и размножиться, а в сложном случае ещё и заранее занять оптимальную (экологическую, социальную и т. п.) нишу для эффективного осуществления двух первых процессов. С другой стороны, необходима информация и о состоянии органов, реализующих ответы на внешние воздействия. Эффекторные органы должны иметь собственную систему рецепторов, которые будут информировать нервный центр о своём состоянии на текущий момент. Нервная система нужна не только для получения дифференциальной информации из внешнего мира, но и для адекватного ответа на неё. Утратив неспецифичность реакций, организм приобретает феноменальную способность приспосабливаться к изменяющейся среде. Чем детальнее информация от органов чувств и точнее адаптирующий ответ, тем приспособленнее оказывается организм.
Однако наша планета — не идеальное место для тех, кто постоянно вынужден кого-то съедать и как-то переносить свой бесценный геном в следующее поколение. Окружающая среда постоянно меняется, и простых адаптивных реакций бывает недостаточно. К счастью, изменения среды подчиняются неким физическим и планетарным законам. Имея возможность сравнивать информацию из внешнего мира во времени, организм получает важное преимущество — опыт предыдущей жизни. Сравнение событий во времени, а не сиюминутных сигналов от различных органов чувств — совершенно новая способность, которая реализуется в нервной системе. Для такого сравнения необходима память (рис. 1–5).
Рис. 1-5. Функциональные блоки сложной нервной системы.


В архитектуре схемы учтены разнообразные органы чувств, системы сравнения одномоментной информации и её сопоставления с предыдущим опытом, который хранится в памяти. Появление памяти и рецепторов эффекторной системы является новым этапом эволюции нервной системы. Эффекторный блок включает в себя железы, изменение концентрации нейрогормонов и мышечные ответы. Контроль за состоянием эффекторных органов достигается рецепторами эффекторной системы.
Энергетическая цена и размеры мозга
Одной из важнейших проблем, с которой сталкивается обладатель нервной системы, — биологическая стоимость этого замечательного органа. Насколько интегративные функции нервной системы окупают затраты на её содержание? Этот вопрос является ключевым в понимании направления и основных путей эволюции нервной системы животных. Абсолютные размеры мозга коррелируют с затратами на его содержание, а относительные — с долей энергетических затрат всего организма. В связи с этим логично рассматривать энергетические затраты организма на нервную систему и её размеры параллельно. По устоявшейся, но необъяснимой традиции под размерами нервной системы понимают массу головного мозга (рис. 1–6). Относительную массу вычисляют как отношение массы мозга к массе тела. Исходя из этих соотношений, определяют уровень обмена и соответствующую долю энергетических затрат на содержание нервной системы. В этих пропорциях, как правило, остаётся неучтённой масса спинного мозга, периферических ганглиев и нервов. Они так же, как и мозг, потребляют кислород и питательные вещества; общая масса спинного мозга и периферической нервной системы может существенно превышать массу головного мозга. Достаточно посмотреть на центральную нервную систему лягушки или змеи (рис. 1–7, а, г). Если добавить к спинному и головному мозгу массу периферической нервной системы, то общее количество нервной ткани будет в несколько раз больше, чем мы привыкли считать.
Читайте также: