
Передача импульсов в нервной системе синапсы и медиаторы








- Физиология
![]()
- История физиологии
![]()
- Методы физиологии



Физиология и строение синапса
Возбуждение с нервного волокна на нервную, мышечную и железистую клетку передается посредством специального структурного образования — синапса.
Синапс — структура, обеспечивающая проведение сигнала от одной клетки к другой. Термин был введен английским физиологом Ч. Шеррингтоном в 1897 г.
Синапсы состоят из трех основных элементов: пресинаптической мембраны, постсинаптической мембраны и синаптической щели (рис. 1).

Рис. 1. Строение синапса: 1 — микротрубочки; 2 — митохондрии; 3 — синаптические пузырьки с медиатором; 4 — пресинаптическая мембрана; 5 — постсинаптическая мембрана; 6 — рецепторы; 7 -синаптическая щель
Некоторые элементы синапсов могут иметь и другие названия. Например, синаптическая бляшка — это синапс между нейронами, концевая пластинка — постсинаптическая мембрана нервно-мышечного синапса, моторная бляшка — пресинаптическое окончание аксона на мышечном волокне.

Пресинаптическая мембрана покрывает расширенное нервное окончание, которое представляет собой нейросекреторный аппарат. В пресинаптической части находятся пузырьки и митохондрии, обеспечивающие синтез медиатора. Медиаторы депонируются в гранулах (пузырьках).
Постсинаптическая мембрана - утолщенная часть мембраны клетки, с которой контактирует пресинаптическая мембрана. Она имеет ионные каналы и способна к генерации потенциала действия. Кроме того, на ней расположены специальные белковые структуры — рецепторы, воспринимающие действие медиаторов.
Синаптическая щель представляет собой пространство между пресинаптической и постсинаптической мембранами, заполненное жидкостью, близкой по составу к плазме крови.

Рис. Строение синапса и процессы, осуществляемые в ходе синаптической передачи сигнала
Синапсы классифицируются по местоположению, характеру действия, способу передачи сигнала.
По месту положения выделяют нервно-мышечные синапсы, нервно-железистые и нейро-нейрональные; последние, в свою очередь, делятся на аксо-аксональные, аксо-дендритические, аксо-соматические, дендро-соматические, дендро-дендротические.
По характеру действия на воспринимающую структуру синапсы могут быть возбуждающими и тормозящими.
По способу передачи сигнала синапсы делятся на электрические, химические, смешанные.
Таблица 1. Классификация и виды синапсов

Синапсы классифицируют следующим образом:
- по местоположению — периферические и центральные;
- по характеру их действия — возбуждающие и тормозящие;
- по способу передачи сигналов — химические, электрические, смешанные;
- по медиатору, с помощью которого осуществляется передача, — холинергические, адренергические, серотонинергические и т.д.
В химических синапсах возбуждение передается с помощью медиаторов (посредников).
Медиаторы — молекулы химических веществ, которые обеспечивают передачу возбуждения в синапсах. Другими словами химические вещества, участвующие в передаче возбуждения или торможения от одной возбудимой клетки к другой.
Свойства медиаторов
- Синтезируются в нейроне
- Накапливаются в окончании клетки
- Выделяются при появлении иона Са2+ в пресинаптическом окончании
- Оказывают специфическое действие на постсинаптическую мембрану
По химическому строению медиаторы можно подразделить на амины (норадреналин, дофамин, серотонин), аминокислоты (глицин, гамма-аминомасляная кислота) и полипептиды (эндорфины, энкефалины). Ацетилхолин известен в основном как возбуждающий медиатор и содержится в различных отделах ЦНС. Медиатор находится в пузырьках пресинаптического утолщения (синаптической бляшки). Медиатор синтезируется в клетках нейрона и может ресинтезироваться из метаболитов его расщепления в синаптической щели.
При возбуждении терминалей аксона происходит деполяризация мембраны синаптической бляшки, вызывающая поступление ионов кальция из внеклеточной среды внутрь нервного окончания через кальциевые каналы. Ионы кальция стимулируют перемещение синаптических пузырьков к пресинаптической мембране, их слияние с ней и последующий выход медиатора в синаптическую щель. После проникновения в щель медиатор диффундирует к постсинаптической мембране, содержащей на своей поверхности рецепторы. Взаимодействие медиатора с рецепторами вызывает открытие натриевых каналов, что способствует деполяризации постсинаптической мембраны и возникновению возбуждающего постсинаптического потенциала. В нервно-мышечном синапсе этот потенциал называется потенциалом концевой пластинки. Между деполяризованной постсинаптической мембраной и соседними с ней поляризованными участками этой же мембраны возникают местные токи, которые деполяризуют мембрану до критического уровня с последующей генерацией потенциала действия. Потенциал действия распространяется по всем мембранам, например, мышечного волокна и вызывает его сокращение.
Выделившийся в синаптическую щель медиатор связывается с рецепторами постсинаптической мембраны и подвергается расщеплению соответствующим ферментом. Так, холинэстераза разрушает медиатор ацетилхолин. После этого некоторое количество продуктов расщепления медиатора поступает в синаптическую бляшку, где из них снова ресинтезируется ацетилхолин.
В организме имеются не только возбуждающие, но и тормозные синапсы. По механизму передачи возбуждения они сходны с синапсами возбуждающего действия. В тормозных синапсах медиатор (например, гамма-аминомасляная кислота) связывается с рецепторами постсинаптической мембраны и способствует открытию в ней каналов для ионов хлора. При этом активизируется проникновение этих ионов внутрь клетки и развивается гиперполяризация постсинаптической мембраны, обусловливающая возникновение тормозного постсинаптического потенциала.
В настоящее время выяснено, что один медиатор может связываться с несколькими различными рецепторами и индуцировать различные реакции.
Химические синапсы
Синапсы с химической передачей возбуждения обладают определенными свойствами:
- возбуждение проводится в одном направлении, так как медиатор выделяется только из синаптической бляшки и взаимодействует с рецепторами на постсинаптической мембраны;
- распространение возбуждения через синапсы происходит медленнее, чем по нервному волокну (синаптическая задержка);
- передача возбуждения осуществляется с помощью специфических медиаторов;
- в синапсах изменяется ритм возбуждения;
- синапсы способны утомляться;
- синапсы обладают высокой чувствительностью к различным химическим веществам и гипоксии.
Одностороннее проведение сигнала. Сигнал передается только от пресинаптической мембраны к постсинаптической. Это вытекает из особенностей строения и свойств синаптических структур.
Замедленная передача сигнала. Обусловлена синаптической задержкой в передаче сигнала с одной клетки на другую. Задержка вызывается временными затратами на процессы выброса медиатора, его диффузии к постсинаптической мембране, связывания с рецепторами постсинаптической мембраны, деполяризации и преобразования постсинаптического потенциала в ПД (потенциал действия). Длительность синаптической задержки колеблется от 0,5 до 2 мс.
Способность к суммации эффекта от приходящих к синапсу сигналов. Такая суммация проявляется, если последующий сигнал приходит к синапсу через короткое время (1- 10 мс) после предыдущего. В таких случаях амплитуда ВПСП возрастает и на постсинаптическом нейроне может генерироваться большая частота ПД.
Трансформация ритма возбуждении. Частота нервных импульсов, приходящих к пресинаптической мембране, обычно не соответствует частоте ПД, генерируемых постсинаптическим нейроном. Исключение составляют синапсы, передающие возбуждение с нервного волокна на скелетную мышцу.
Низкая лабильность и высокая утомляемость синапсов. Синапсы могут проводить 50-100 нервных импульсов в секунду. Это в 5-10 раз меньше, чем максимальная частота ПД, которую могут воспроизводить нервные волокна при их электростимуляции. Если нервные волокна считаются практически неутомляемыми, то в синапсах утомление развивается весьма быстро. Это происходит из-за истощения запасов медиатора, энергетических ресурсов, развития стойкой деполяризации постсинаптической мембраны и т.д.
Высокая чувствительность синапсов к действию биологически активных веществ, лекарственных препаратов и ядов. Например, яд стрихнин блокирует функцию тормозных синапсов ЦНС, связываясь с рецепторами, чувствительными к медиатору глицину. Столбнячный токсин блокирует тормозные синапсы, нарушая выделение медиатора из пресинаптической терминали. В обоих случаях развиваются опасные для жизни организма явления. Примеры действия биологически активных веществ и ядов на передачу сигналов в нервно-мышечных синапсах рассмотрены выше.
Свойства облегчения и депрессии синоптической передачи. Облегчение синаптической передачи имеет место, когда нервные импульсы поступают к синапсу через короткое время (10-50 мс) друг за другом, т.е. достаточно часто. При этом в течение некоторого промежутка времени каждый последующий ПД, приходящий к пресинаптической мембране, вызывает увеличение содержания медиатора в синаптической щели, возрастание амплитуды ВПСП и увеличение эффективности синаптической передачи.
Одним из механизмов облегчения является накопление ионов Са 2 в пресинаптической терминали. Для удаления кальциевым насосом порции кальция, вошедшей в синаптическую терминаль при поступлении ПД, необходимо несколько десятков миллисекунд. Если в это время приходит новый потенциал действия, то новая порция кальция входит в терминаль и ее эффект на высвобождение нейромедиатора складывается с остаточным количеством кальция, которое кальциевый насос не успел удалить из нейроплазмы терминали.
Имеются и другие механизмы развития облегчения. Этот феномен в классических руководствах по физиологии называют также посттетанической потенциацией. Облегчение синаптической передачи имеет значение в функционировании механизмов памяти, для образования условных рефлексов и обучения. Облегчение передачи сигналов лежит в основе развития пластичности синапсов и улучшения их функций при частой активации.
Депрессия (угнетение) передачи сигналов в синапсах развивается при поступлении очень частых (для нервно-мышечного синапса более 100 Гц) нервных импульсов к пресинаптической мембране. В механизмах развития явления депрессии имеют значение истощение запасов медиатора в пресинаптической терминали, снижение чувствительности рецепторов постсинаптической мембраны к медиатору, развитие стойкой деполяризации постсинаптической мембраны, затрудняющих генерацию ПД на мембране постсинаптической клетки.
Электрические синапсы
Кроме синапсов с химической передачей возбуждения в организме есть синапсы с электрической передачей. Эти синапсы имеют очень узкую синаптическую щель и пониженное электрическое сопротивление между двумя мембранами. Благодаря наличию поперечных каналов между мембранами и низкому сопротивлению, электрический импульс легко проходит через мембраны. Электрические синапсы обычно характерны для однотипных клеток.
В результате воздействия раздражителя пресинаптический потенциал действия раздражает постсинаптическую мембрану, где возникает распространяющийся потенциал действия.
Электрические синапсы характеризуются большей скоростью проведения возбуждения по сравнению с химическими синапсами и низкой чувствительностью к воздействию химических веществ.
Электрические синапсы бывают с одно- и двусторонней передачей возбуждения.
В организме встречаются и электрические тормозные синапсы. Тормозное влияние развивается за счет действия тока, который вызывает гиперполяризацию постсинаптической мембраны.
В смешанных синапсах может происходить передача возбуждения с помощью как электрических импульсов, так и медиаторов.
Синапс – это структурно-функциональное образование, обеспечивающее переход возбуждения или торможения с окончания нервного волокна на иннер-вирующую клетку.
1) пресинаптическая мембрана (электрогенная мембрана в терминале аксона, образует синапс на мышечной клетке);
2) постсинаптическая мембрана (электрогенная мембрана иннервируемой клетки, на которой образован синапс);
3) синаптическая щель (пространство между преси-наптической и постсинаптической мембраной, заполнена жидкостью, которая по составу напоминает плазму крови).
Существует несколько классификаций синапсов.
1. По локализации:
1) центральные синапсы;
2) периферические синапсы.
Центральные синапсы лежат в пределах центральной нервной системы, а также находятся в ганглиях вегетативной нервной системы.
Различают несколько видов периферических синапсов:
2. Функциональная классификация синапсов:
1) возбуждающие синапсы;
2) тормозящие синапсы.
3. По механизмам передачи возбуждения в синапсах:
Передача возбуждения осуществляется при помощи медиаторов. Различают несколько видов химических синапсов:
1) холинэргические. В них происходит передача возбуждения при помощи ацетилхолина;
2) адренэргические. В них происходит передача возбуждения при помощи трех катехоламинов;
3) дофаминэргические. В них происходит передача возбуждения при помощи дофамина;
4) гистаминэргические. В них происходит передача возбуждения при помощи гистамина;
5) ГАМКэргические. В них происходит передача возбуждения при помощи гаммааминомасляной кислоты, т. е. развивается процесс торможения.
Синапсы имеют ряд физиологических свойств:
1) клапанное свойство синапсов, т. е. способность передавать возбуждение только в одном направлении с пресинаптической мембраны на постсинап-тическую;
2) свойство синаптической задержки, связанное с тем, что скорость передачи возбуждения снижается;
3) свойство потенциации (каждый последующий импульс будет проводиться с меньшей постсинапти-ческой задержкой);
4) низкая лабильность синапса (100–150 имульсов в секунду).
При деполяризации пресинаптической терминали открываются потенциал-чувствительные кальциевые каналы, ионы кальция входят в пресинаптическую терминаль и запускают механизм слияния синаптических пузырьков с мембраной. В результате медиатор выходит в синаптическую щель и присоединяется к белкам-рецепторам постсинаптической мембраны, которые делятся на метаботропные и ионотропные. Первые связаны с G-белком и запускают каскад реакций внутриклеточной передачи сигнала. Вторые связаны с ионными каналами, которые открываются при связывании с ними нейромедиатора, что приводит к изменению мембранного потенциала. Медиатор действует в течение очень короткого времени, после чего разрушается специфическим ферментом. Например, в холинэргических синапсах фермент, разрушающий медиатор в синаптической щели — ацетилхолинэстераза. Одновременно часть медиатора может перемещаться с помощью белков-переносчиков через постсинаптическую мембрану (прямой захват) и в обратном направлении через пресинаптическую мембрану (обратный захват). В ряде случаев медиатор также поглощается соседними клетками нейроглии.
Открыты два механизма высвобождения: 1 везикула соединяется с мембраной, и из неё в синаптическую щель выходят небольшие молекулы, а крупные остаются в везикуле. Второй механизм, предположительно, быстрее первого, с помощью него происходит синаптическая передача при высоком содержании ионов кальция в синаптической бляшке.
Понятие о нервном центре. Особенности проведения возбуждения через нервные центры (одностороннее проведение, замедленное проведение, суммация возбуждения, трансформация и усвоение ритма).
Нервный центр - сложное сочетание, “ ансамбль” нейронов, согласованно включающийся в регуляцию определенной функции или в осуществление рефлекторного акта. Клетки нервного центра связаны между собой синаптическими контактами и отличаются огромным разнообразием и сложностью внешних и внутренних связей. В соответствии с выполняемой функцией выделяют чувствительные центры, центры вегетативных функций, двигательные центры и др. Различные нервные центры характеризуются определенной топографией в пределах ЦНС.
в физиологическом смысле нервный центр - это функциональное объединение группировок нервных элементов с целью выполнения сложных рефлекторных актов.
Нервные центры состоят из множества нейронов, связанных между собой еще большим множеством синаптических связей. Это обилие синапсов определяют основные, свойства нервных центров: односторонность проведения возбуждения, замедление проведения возбуждения, сум-мацию возбуждений, усвоение и трансформацию ритма возбуждений, следовые процессы и легкую утомляемость.
а) Ганглионарная передача нервных импульсов. Преганглионарные нейроны симпатической и парасимпатической систем — холинергические: при образовании аксодендритических синапсов с ганглионарными клетками из этих нейронов высвобождается ацетилхолин (АХ). Рецепторы на поверхности ганглионарных клеток называют никотиновыми, так как их возбуждение может происходить при местном действии никотина.
б) Передача нервных импульсов в нейроэффекторном синапсе. Постганглионарные нервные волокна симпатической и парасимпатической систем образуют нейроэффекторные соединения (синапсы)с эффекторными тканями (тканями-мишенями). За высвобождение нейромедиаторов отвечают многочисленные пресинаптические утолщения, расположенные по ходу нервных волокон.
Главный нейромедиатор в симпатических нейроэффекторных соединениях — норадреналин (норэпинефрин), который высвобождается из гранулярных везикул. Постганглионарные симпатические волокна — преимущественно адренергические; исключение составляют холинергические волокна, отвечающие за симпатическую иннервацию расположенных по всему телу эккриновых потовых желез. Главный нейромедиатор в парасимпатических нейроэффекторных соединениях — ацетилхолин (АХ). Постганглионарные парасимпатические волокна преимущественно холинергические.
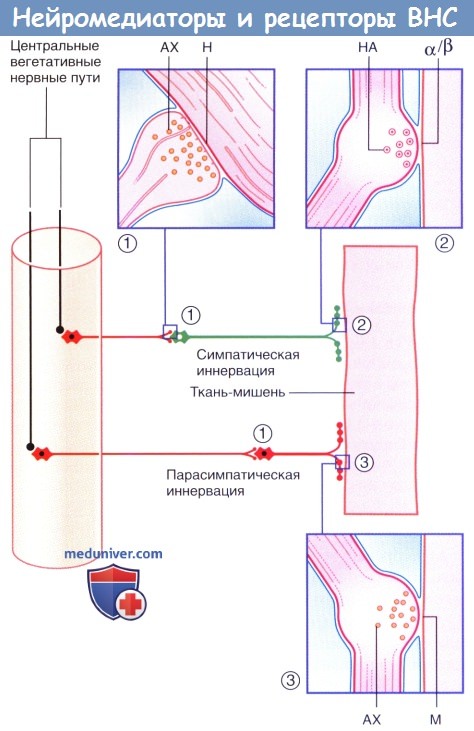
Нейромедиаторы и рецепторы вегетативной нервной системы.
(1) Аксодендритические синапсы с никотиновыми рецепторами.
(2) Нейроэффекторные синапсы с адренергическими рецепторами.
(3) Нейроэффекторные синапсы с мускариновыми рецепторами. Ганглионарные нейроны и постганглионарные волокна выделены красным цветом.
АХ — ацетилхолин; М — мускариновые рецепторы; Н — никотиновые рецепторы; НА — норадреналин.
в) Рецепторы в нейрозффекторном синапсе. Физиологические эффекты вегетативной нервной системы (ВНС) зависят от вида рецепторов на постсинаптической мембране (плазматической мембране эффекторных клеток). На высвобождение нейромедиаторов также влияют рецепторы пресинаптической мембраны (аксолеммы).
- Синаптические рецепторы симпатической системы (адренорецепторы). Для норадреналина существуют два вида а-адренорецепторов и два вида β-адренорецепторов.
1. Постсинаптические α1-адренорецеторы, активация которых вызывает сокращение гладких мышц мелких периферических артерий и крупных артериол, дилататора зрачка, а также мышц семявыносящего протока, сфинктеров ЖКТ и шейки мочевого пузыря.
2. Пресинаптические α2-адренорецеторы локализуются как на симпатических, так и на парасимпатических нервных окончаниях и ингибируют в них высвобождение нейромедиаторов. Пресинаптические α2-адренорецеторы симпатической системы называют ауторецепторами.
3. Постсинаптические адренорецепторы, возбуждение которых приводит к увеличению пейсмекерной активности клеток сердца, а также к повышению силы сердечных (желудочковых) сокращений. При резком падении артериального давления за счет активации симпатических β1-адренорецепторов юкстагломерулярных клеток почек происходит секреция ренина, который, в свою очередь, вызывает секрецию мощного вазоконстриктора ангиотензина II.
4. β2-Адренорецепторы, которые реагируют как на норадреналин, так и на адреналин (эпинефрин).
Активация постсинаптических β2-адренорецепторов вызывает расслабление гладких мышц, наиболее ярко выраженное в трахеобронхиальном дереве и мышцах глаза, участвующих в аккомодации. Часть постсинаптических β2-адренорецепторов располагается на поверхности гепатоцитов. При возрастании энергетических затрат организма их активация вызывает распад гликогена, необходимый для поддержания уровня глюкозы в крови.
Пресинаптические β2-адренорецепторы адренергических нервных окончаний отвечают за высвобождение норадреналина.
В симпатических нервных окончаниях большая часть высвобождаемого норадреналина подвергается обратному захвату моноаминным ферментным насосом. После обратного захвата часть норадреналина разрушается митохондриальным ферментом — моноаминоксидазой (МАО). Влияние лекарственных средств на симпатическую систему подробно рассмотрено в блоке клинической информации ниже.
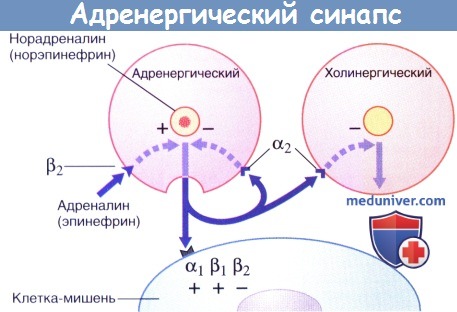
Передача возбуждения в адренергическом нейроэффекторном синапсе.
Секреция норадреналина стимулируется адреналином и ингибируется путем активации пресинаптических α2-рецепторов (данные рецепторы активируются и на пресинаптической мембране близлежащих парасимпатических синапсов, где также ингибируют секрецию).
- Синаптические рецепторы парасимпатической системы. Парасимпатические синаптические рецепторы называют мускариновыми, так как их активацию, помимо АХ, может вызывать другое химическое вещество — мускарин. Стимуляция парасимпатической системы приводит к следующим М-холинергическим эффектам.
• Замедление (в ответ на стимуляцию блуждающего нерва) частоты сердечных сокращений, а также уменьшение силы желудочковых сокращений.
• Сокращение гладких мышц, которое обусловливает такие процессы, как перистальтика кишечника, опустошение мочевого пузыря, а также рефлекс аккомодации при взгляде на близкое расстояние.
• Секреция желез.
Помимо представленных выше эффектов, пресинаптические мускариновые рецепторы симпатических нервных окончаний также отвечают за ингибирование высвобождения норадреналина.
Влияние лекарственных средств на парасимпатическую систему подробно рассмотрено в Блоке клинической информации. Лекарственные средства, обладающие мускариноподобным действием, называют холинергическими (холиномиметическими). Лекарственные средства, блокирующие доступ АХ к постсинаптической мембране, называют антихолинергическими (холинолитическими).
Главный фактор, который необходимо учитывать при назначении препаратов, активирующих или подавляющих деятельность симпатической и парасимпатической систем,— наличие рецепторов к этим лекарственным веществам α-, β- и мускариновых) в ЦНС. Так, в частности, применение данных лекарственных средств в психиатрии обусловлено их действием на центральные, а не на периферические рецепторы.
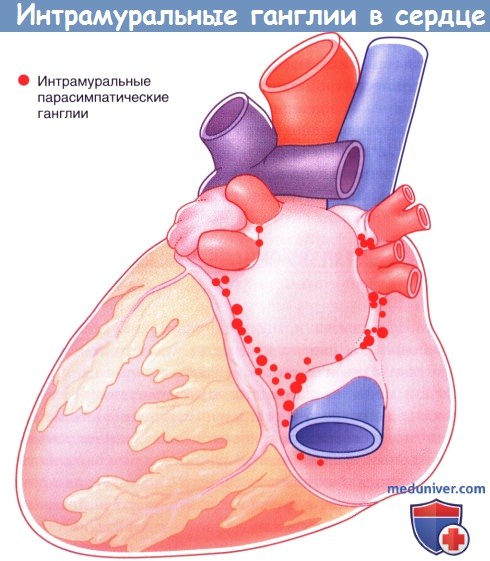
Расположение парасимпатических интрамуральных ганглиев в сердце. 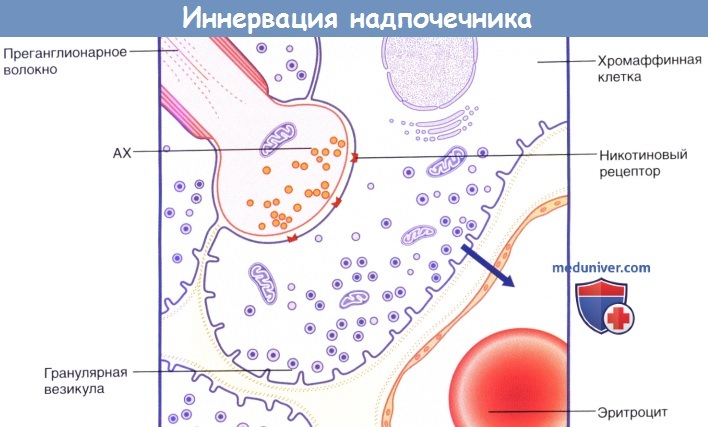
Синаптическое соединение преганглионарного волокна грудного внутренностного нерва и хромаффинной клетки мозгового вещества надпочечников.
Активация никотиновых рецепторов ацетилхолином (АХ).
8% клеток содержат крупные гранулярные везикулы (представлены на рисунке), высвобождающие адреналин; его секрекция в капиллярное русло обозначена стрелкой.
20% клеток содержат маленькие гранулярные везикулы, высвобождающие норадреналин. 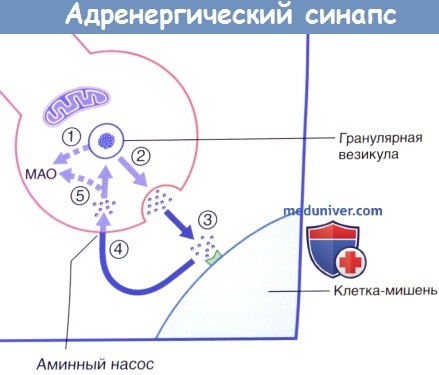
Высвобождение и обратный захват нейромедиатора в адренергическом нервном окончании.
МАО — моноаминоксидаза. 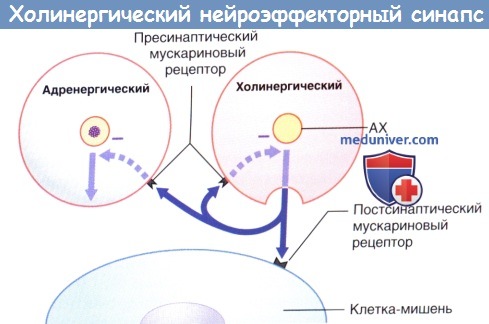
Передача возбуждения в холинергическом нейроэффекторном синапсе.
Избыточная секреция ацетилхолина (АХ) ингибируется путем активации пресинаптических мускариновых рецепторов (данные рецепторы активируются и на пресинаптической мембране близлежащих симпатических синапсов, где также ингибируют секрецию).
г) Другие типы нейронов. И в симпатическом, и в парасимпатическом отделах ВНС часть нейронов представлена неадренергическими, нехолинергическими нейронами (NANC-нейронами). Находящиеся в симпатических ганглиях небольшие вставочные нейроны отвечают за высвобождение дофамина — предшественника норадреналина. Часть дофамина выделяется в капиллярное русло, другая часть связывается с дофаминовыми рецепторами на основных (адренергических) нейронах, оказывая на них слабое тормозное действие.
Самое большое количество NANC-нейронов находится в области ганглионарных клеток стенки ЖКТ и в тазовых ганглиях. Эти нейроны содержат более 50 разнообразных белковых субстанций, находящихся в несвязанном состоянии или в различных комбинациях. Функция большей части этих веществ — регуляторная: действуя на пресинаптическую или постсинаптическую мембрану, они влияют на продолжительность работы основных нейромедиаторов. Другие белковые субстанции (вещества, выделяющиеся совместно с ацетилхолином (АХ)) представляют собой комедиаторы (сопутствующие медиаторы).
Редактор: Искандер Милевски. Дата публикации: 14.11.2018
Медиаторами вегетативной нервной системы являются ацетилхолин, адреналин и норадреналин. Медиаторы вегетативной нервной системы действуют на клетки органов значительно дольше, чем медиаторы, вырабатываемые в окончаниях соматических нервов. Это объясняется меньшей активностью ферментов, разрушающих медиатор. В зависимости от того, какой медиатор выделяется в окончаниях вегетативных нейронов, их делят на холинергические и адренергические. Холинергическими являются все пре- и постганглионарные нейроны парасимпатической нервной системы, а также преганглионарные симпатические нейроны и постганглионарные симпатические нейроны, идущие к потовым железам и обеспечивающие расширение сосудов работающих мышц. Адренергическими являются все остальные постганглионарные нейроны симпатической нервной системы.
Освобождающийся в аксонах медиатор взаимодействует со специфическим белком постсинаптической мембраны, образуя при этом комплексное соединения. Белок, с которым взаимодействует ацетилхолин, получил название холинорецептора. Белок, с которым взаимодействует адреналин, называется адренорецептором. Имеется два вида адренорецепторов: альфа- и бета. В различных органах и тканях может находиться один из видов рецепторов или оба одновременно. Например, в кровеносных сосудах находятся альфа- и бета-рецепторы, а в сердце и бронхах только бетa-рецепторы. Они могут вызывать однонаправленный (в кишечнике) и разнонаправленный эффекты (в стенке артериол). Среди холинорецепторов существует два вида структур: М-холинорецепторы, которые теряют чувствительность к ацетилхолину под действием атропина и Н-холинорецепторы, которые теряют чувствительность к ацетилхолину под влиянием никотина и других веществ, получивших название ганглиоблокаторов.
Проведение возбуждения осуществляется в вегетативных ганглиях и под действием других веществ. В окончаниях симпатических нервов выделяется дофамин, он вступает во взаимодействие с альфа-адренорецепторами, расположенными на самих пресинаптических окончаниях и тем самым тормозит выделение норадреналина. Серотонин действует на гладкую мускулатуру кишечника, матки и кровеносных сосудов. Эффект его напоминает действие ацетилхолина, но сохраняется после блокады М-холинорецепторов. В желудке и кишечнике обнаружены нейроны, возбуждение которых тормозит активность гладкой мускулатуры. Эти нейроны получили название пуринергических и в них выделяется пуриновый нуклеотид АТФ. Также передача осуществляется с помощью тормозных медиаторов (глицин и ГАМК), тканевых гормонов и трансдукторов. Для выполнения своей функции по поддержанию гомеостаза вегетативной нервной системы наряду с обыкновенными клетками содержит клетки, которые принимают информацию обычные способом, а отвечают эндокринной секрецией. К ним относятся хромаффинные клетки мозгового слоя надпочечников, которые в ответ на электрический сигнал выделяют адреналин и норадреналин; юкстрагломерулярные клетки почки, выделяющие ренин в ответ на активацию постганглионарного симпатического волокна; нейроны гипоталамуса, выделяющие вазопрессин и окситоцин; нейроны аденогипофиза, выделяющие в сосудистое русло факторы регуляции. Аксоны этих клеток не образуют синапсы с другими нейронами. Они свободно заканчивается вокруг сосудов, с которым образуют так называемые гемальные органы. Хромаффинные клетки находятся в надпочечниках в виде скоплений на поверхности аорты, в области каротидного синуса, среди клеток симпатических ганглий. На этих клетках заканчиваются симпатические преганглионары. При их активации в кровь выделяется смесь, состоящая из 80% адреналина и 20% норадреналина.
Экстремальные ситуации типа гипотермии, гипоксии, чрезмерного физического напряжения приводят к активации симпатоадреналовой системы. При этом активируется сама симпатическая нервная система, кроме того, увеличивается выброс катехоламинов из систем хромаффинных клеток.
Вегетативная нервная система осуществляет следующие рефлексы:
1. Висцеро-висцеральный рефлекс возникает во внутренних органах, где и заканчивается. Основой для этих процессов являются местные рефлекторные дуги. Например, механическое раздражение кишечника изменяет частоту сердечных сокращений.
2. Висцеросоматический рефлекс, при нем возбуждение возникает во внутренних органах, а в результате происходит возбуждение или торможение активности скелетных мышц. Например, раздражение рецепторов пищеварительного тракта вызывает сокращение мышц брюшного пресса или движение конечностей.
В результате сегментарной организации автономной и соматической иннервации при заболеваниях внутренние органов в ограниченных участках кожи возникает повышение тактильной и болевой чувствительности. Эти боли получили название отраженных, а области, в которых они появляются - зоны Захарьина-Геда. В этом случае висцеральные и кожные чувствительные волокна конвергируют к одним и тем же нейронам спиноталамического пути. В этих путях происходит потеря информации, в результате чего кора головного мозга приписывает возникающее возбуждение раздражению кожной области. Висцеро-соматические рефлексы бывают пусковые и корригирующие.
3. Соматовисцеральные рефлексы начинаются с активации экстерорецепторов, а затем происходит изменение функционального состояния внутренних органов, например, сосудистая реакция при термическом воздействии на рецепторы кожи.
4. Висцеросенсорный рефлекс заключается в том, что при раздражении внутренних органов происходит не только изменения в мышечной системе, но и изменяется соматическая чувствительность. Эта зона ограничивается определенным участком кожи, и для вызова такого рефлекса необходимо длительное и сильное воздействие.
5. Для автономной нервной системы характерно такое явление как аксон-рефлекс. Открыт он был Н.М.Соковниным в 1877 г. и далее изучен Дж. Ленгли, который объяснял это явление как местный рефлекс, осуществляемый в ганглии без участия центральной нервной системы. Сейчас аксон-рефлексом называют такие рефлекторные процессы, которые осуществляются по разветвлениям аксона без участия тела нейрона. В этом случае возбуждение идет по одной ветви аксона, затем переходит на другую ветвь и по ней эфферентно направляется к эффекторному органу. Например, при раздражении волокон от кожных болевых рецепторов возникает расширение сосудов и покраснение области, иннервируемой этими волокнами. Этот эффект удается получить и после разрушения спинного мозга и после удаления симпатических ганглиев. Существует и другое объяснение понятия аксон-рефлекс. Считают, что при раздражении кожных болевых рецепторов из них выделяется биологически активные вещества типа АТФ или вещества Р, обладающих сосудорасширяющим действием.
Влияние автономной нервной системы на деятельность
Внутренних органов
Симпатические нервы оказывают влияние не только на гладкую мускулатуру и железы, но и на скелетные мышцы. Это феномен Орбели-Гинецинского (при раздражении симпатических нервов работоспособность утомленной мышцы повышается). Симпатические нервы не вызывают сокращения мышцы, но изменяют состояние мышечной ткани, повышая ее восприимчивость к импульсам, которые передаются по соматическим нервам. В мышце в этом случае растет потребление кислорода и увеличивается содержание энергосодержаших соединений. Также было установлено, что стимуляция симпатического нерва изменяет возбудимость рецепторов и функциональные свойства центральной нервной системы. Например, при раздражении симпатических волокон языка возрастает вкусовая чувствительность. Эти факты изучил и обобщил Л.А. Орбели и создал теорию об адаптационно- трофической функции симпатической нервной системы.
Таким образом, симпатическая нервная система активирует деятельность нервной системы в целом, повышает защитные свойства организма (иммунитет, свертывание крови, терморегуляция). Следовательно, в процессе эволюции симпатическая нервная система превратилась в особый инструмент мобилизации всех ресурсов организма в тех случаях, когда возникает угроза самому организму. В результате такого действия симпатическая нервная система приводит к изменению гомеостатических констант организма, что выражается в повышении кровяного давления, выбросе крови из депо, поступлении в кровь ферментов, глюкозы, снижении мочеобразования, угнетении функции пищеварительной системы. Удаление симпатической нервной системы не сопровождается значительными расстройствами висцеральный функций, но организм не в состоянии осуществить физические усилия, медленно восстанавливается после кровотечения, плохо переносит повышение и понижение температуры, страдает отсутствием аппетита.
Парасимпатическая нервная система обладает ограниченными областями влияния. Во внутренних органах преганглионарное волокно оканчивается не на эффекторе – мышечных или железистых клетках –, а на интернейроне или эфферентном нейроне метасимпатической нервной системы. Таким образом, влияния парасимпатической нервной системы оказываются не прямыми, а опосредованными, поэтому эффект раздражения не бывает однозначным и зависит от текущих внутриорганных процессов. Но в общем действие этой системы направлено на поддержание гомеостаза организма, который может быть нарушен в результате действия симпатической нервной системы.
Последнее изменение этой страницы: 2016-08-12; Нарушение авторского права страницы
Читайте также: