
Периферические синапсы вегетативной нервной системы
Синапсы вегетативной нервной системы имеют в целом такое же строение, что и центральные.
Передача нервных импульсов с преганглионарных волокон на нейроны всех вегетативных ганглиев осуществляется Н-холинергическими синапсами, т.е. синапсами, на постсинаптической мембране которых расположены никотинчувствительные холинорецепторы.
Постганглионарные холинергические волокна образуют на клетках исполнительных органов М-холинергические синапсы. Их постсинаптическая мембрана содержит мускаринчувствительные рецепторы (блокатор - атропин).
И в тех, и в других синапсах передача возбуждения осуществляется ацетилхолином. М-холинергические синапсы оказывают возбуждающее влияние на гладкие мышцы пищеварительного канала, мочевыводящей системы (кроме сфинктеров), железы ЖКТ. Однако они уменьшают возбудимость, проводимость и сократимость сердечной мышцы и вызывают расслабление некоторых сосудов головы и таза.
Постганглионарные синаптические волокна образуют 2 типа адренергических синапсов на эффекторах: альфа-адренегрические и бета-адренергические. Постсинаптическая мембрана первых содержит бета1- и бета2- адренорецепторы.
постгаплионарными волокнами иннервируются клетки подкожно-жировой клетчатки и печени и, возможно, канальцы почек и лимфатические образования (например, вилочковая железа, селезенка, пейеро-вы бляшки и лимфатические узлы). Постганглионарные симпатические волокна, иннервирующие потовые железы и сосуды скелетных мышц, выделяют ацетилхолин, являются холинергическими.
Норадреналин — нейромедиатор адренергической системы. Другая важная составляющая ВНС — адренергическая система. До сих пор неизвестно, какой нейромедиатор использовался в этой системе первоначально — эпинефрин или норэпинефрин. Сейчас известно, что за исключением надпочечников, которые секретируют эпинефрин (адреналин), нейроме-диатором в адренергической системе является норэпинефрин. Ацетилхолин — ганглионарный медиатор для холинергической и адренергической систем. Эфферентные нервы и для холинергической, и для адренергической систем происходят из соответствующих частей ствола мозга и спинного мозга. Эфферентные нервы образуют синапс в ганглии, расположенном вне органа.
32. Холинэргические и адренэргические нервы. Биохимический механизм передачи возбуждения в холинэргических и адренэргических нервах. М- и Н-холинореактивные системы.
Симпатические и парасимпатические нервные волокна секретируют в основном один из двух синаптических медиаторов — ацетилхолин или норадреналин. Волокна, секретирующие ацетилхолин, называют холинергическими, волокна, секретирующие норадреналин, называют адренергическими (термин, происходящий от адреналина, — альтернатива эпинефрину).
Все преганглионарные нейроны (и симпатической, и парасимпатической нервных систем) являются холинергическими. Ацетилхолин или подобные ему вещества при действии их на ганглии возбуждают симпатические и парасимпатические постганглионарные нейроны. Все или почти все постганглионарные нейроны парасимпатической системы — также холинергические.
С другой стороны, большинство постганглионарных симпатических нейронов являются адренергическими. Однако постганглионарные симпатические нервные волокна, идущие к потовым железам, мышцам, поднимающим волосы, и к очень небольшому числу кровеносных сосудов, являются холинергическими.
Холинергические механизмы нервной системы - это вещества, которые обеспечивают передачу возбуждения в холинергическом синапсе.
Медиатор ацетилхолин (эфир холина и уксусной кислоты) образуется из аминокислоты холина и ацетил-СоА на пресинаптическом окончании нервноего волокна. Образующийся медиатор поступает в везикулы, а частично может остаться в свободном состоянии. При возбуждении медиатор выделяется из везикул. Процесс выделения медиатора С-зависим. Для нормальной работы синапса необходим запас медиатора, поэтому на пресинаптической мембране идёт ресинтез ацетилхолина. Для этого аминокислота холин выделяется из постсинаптической мембраны, частично из синаптической щели (возврат медиатора). Для образования медиатора необходима энергия метехондрий.
Фермент, способствующий синтезу ацетилхолина - ацетилхолинтрансфераза или холинацетилаза. Этот фермент образуется в теле нейрона и поступает в нервные окончания. Для нормального образования медиатора необходима целостность тела нейрона. Изолированное нервное волокно не может долго выделять медиатор.
Фермент, расщепляющий ацетилхолин - ацетилхолинэстераза. Этот фермент обладает высоким сродстворм к ацетилхолину, который находится в виде комплекса и Х-рецептором. Различают истинную ацетилхолинэстеразу (находится в синапсах и эритроцитах), которая расщепляет ацетилхолин в физиологических концентрациях и ложную ацетилхолинэстеразу (в жидкостях организма - слюне, плазме и т. д.), которая расщепляет ацетилхолин в высоких концентрациях и разрушает еще и различные производные ацетилхолина (курарекодовые препараты). Освобождённый холин с помощью переносчиков поступает на пресимпатическую мембрану, а уксусная кислота и глюкоза поступают в кровь через межтканевую жидкость.
Адренергические механизмы нервной системы осуществляются за счет норадреналина - составляет 90 % и других катехоламинов - 10 %.
Предшественник норадреналина - изопропилнораденалин, дофамин. Для синтеза необходимы аминокислоты тиронин, фениламин, которые поступают с постсинапсической мембраны и из тела нейрона. Любые структуры могут образовывать норадреналин, но 95 % его образуется на пресимпатической мембране.
Ферменты синтеза норадреналина - трансаминазы.
Ферменты разрушения ноадреналина - группа катехоламинтрансфераз, часто моноаминоуксусная кислота и моноаминооксидант.
Адренорецепторы - белковые молекулы, обладающие сродством к норадреналину и его производным. Эти рецепторы - наружная субъединица крайней белковой молекулы, внутренняя субъединица может быть ферментом (адемилат- и гуанилатциклазы). При взаимодействии с рецептором изменяется структура молекулы белка и, как следствие, изменяется активность фермента.
Существует 2 вида холинорецепторов - М и Н.
М-холинорецепторы - чувствительны к мускалину (яду мухомора) - расположены в основном во внутренних органах, эндокринных железах, сердце, сосудах, дыхательных путях, желудочнокишечном тракте. Они обладают медленным, но продолжительным действием, могут суммировать возбуждение. Существуют 2 вида М-холинорецепторов: одна - во внутренних органах, другая - в эндокринных железах. При возбуждении М-холинорецепторв происходит торможение сердечной деятельности, раширение сосудов, активация деятельности желудочно-кишечного тракта, изменяется секреция некоторых эндокринных желёз.
Н-холинорецепторы - чувствительны к никотину. Располагаются в вегетативных ганглиях, мионевральных синапсах, в хлорофильной ткани надпочечников. Эти рецепторы обладают быстрым, кратковременным действием, не могут суммировать возбуждение. Существует 3 разновидности. За счёт наличия разновидностей рецепторы могут блокироваться различными веществами. В центральной нервной системе больше Н-холинорецепторов. М-холинорецепторы преобладают в области ствола мозга, подкорковых узлах, лимбической системе, ретикулярной формации, гипоталамусе.
33. Химическая передача возбуждения в ганглиях симпатической нервной системы. Ацетинхолин как передатчик возбуждения в ганглиях. Роль холиностеразы. Ганглиоблокирующие вещества и их роль в лекарственной терапии.
Основным же способом передачи возбуждения в автономной нервной системе является химический. Он осуществляется по определенным закономерностям, среди которых выделяют два принципа. Первый (принцип Дейла) заключается в том, что нейрон со всеми отростками выделяет один медиатор. Как стало теперь известно, наряду с основным в этом нейроне могут присутствовать также другие передатчики и участвующие в их синтезе вещества. Согласно второму принципу, действие каждого медиатора на нейрон или эффектор зависит от природы рецептора постсинаптической мембраны.В автономной нервной системе насчитывают более десяти видов нервных клеток, которые продуцируют в качестве основных разные медиаторы: ацетилхолин, норадреналин, серотонин и другие биогенные амины, аминокислоты, АТФ.Каждый из медиаторов выполняет передаточную функцию, как правило, в определенных звеньях дуги автономного рефлекса.Так, ацетилхолин выделяетсяв окончаниях всех преганглионарных симпатических и парасимпатических нейронов, а также большинства постганглионарных парасимпатических окончаний. Кроме того, часть постганглионарных симпатических волокон, иннервирующих потовые железы и, по-видимому, вазодилататоры скелетных мышц, также осуществляют передачу с помощью ацетилхолина.Медиатор, освобождающийся в пресинаптических терминалах под влиянием приходящих нервных импульсов, взаимодействует со специфическим белком-рецептором постсинаптической мембраны и образует с ним комплексное соединение. Белок, с которым взаимодействует ацетилхолин, носит название холинорецептора, адреналин или норадреналин — адренорецептора и т. д. Местом локализации рецепторов различных медиаторов является не только постсинаптическая мембрана. Обнаружено существование и специальных пресинаптических рецепторов, которые участвуют в механизме обратной связи регуляции медиаторного процесса в синапсе.Ацетилхолинэстераза играет ключевую роль в процессах нейрогуморальной и синаптической передачи: в холинэргических синапсах катализирует гидролиз ацетилхолина, и, как следствие, прекращает влияние данного медиатора на холинорецептор, отвечающий за возбуждение нервного волокна. При ингибировании АХЭ освобождение рецепторов от ацетилхолина происходит очень медленно (только посредством диффузии), и передача нервных импульсов заблокирована на уровне (нейротрансмиттер постсинаптическая мембрана). Это вызывает дезорганизацию процессов организма, а при тяжелых отравлениях (в частности фосфорорганическими боевыми отравляющими веществами) может привести к летальному исходу.Ганглиоблокирующие вещества обладают способностью блокировать н-холинорецепторы вегетативных нервных узлов и в связи с этим тормозить передачу нервного возбуждения с преганглионарных на постганглионарные волокна вегетативных нервов. Современные ганглиоблокаторы угнетают или полностью выключают проведение нервного импульса в симпатических и парасимпатических узлах, синокаротидном клубочке и хромафинной ткани надпочечников, что приводит к временной искусственной денервации внутренних органов и изменению их функции. Однако разные препараты могут обладать различной активностью по отношению к разным группам ганглиев. Первым ганглиоблокатором, получившим практическое применение в медицине в начале 50-х годов, был гексаметоний (гексоний). Затем был получен целый ряд других ганглиоблокаторов; некоторые из них, подобно гексаметонию, являются четвертичными аммониевыми соединениями, а часть является третичными аминами.
34. Значение вегетативной нервной системы в деятельности целого организма. Адаптационно-трофическое значение вегетативной нервной системы организма.
Главной функцией автономной нервной системы является регулирование процессов жизнедеятельности органов тела, согласование и приспособление их работы к общим нуждам и потребностям организма в условиях окружающей среды. Выражением этой функции служит регуляция метаболизма, возбудимости и других сторон деятельности органов и самой ЦНС. В этом случае управление работой тканей, органов и систем осуществляется посредством двух типов влияний — пусковых и корригирующих.Влияние автономной нервной системы на висцеральные функции. Все структуры и системы организма иннервируются волокнами автономной нервной системы. Многие из них имеют двойную, а полые висцеральные органы даже тройную (симпатическую, парасимпатическую и метасимпатическую) иннервацию. Изучение роли каждой из них обычно осуществляют с помощью электрического раздражения, хирургического или фармакологического выключения, химической стимуляции и т. д.Основная функциональная роль метасимпатической части автономной нервной системы состоит в осуществлении механизмов, обеспечивающих гомеостаз — относительное динамическое постоянство внутренней среды и устойчивость основных физиологических функций. В отличие от нее симпатическая часть автономной нервной системы рассматривается как система тревоги, мобилизации защитных сил и ресурсов для активного взаимодействия с факторами среды. Задачу восстановления и поддержания этого постоянства, нарушенного в результате возбуждения симпатической части автономной нервной системы, берет на себя метасимпатическая и отчасти парасимпатическая части автономной нервной системы.Адаптационно-трофическая функция симпатической части автономной нервной системы.Эффекты адаптационно-трофического влияния, полученные сначала при раздражении симпатических волокон, полностью воспроизводятся раздражением гипоталамической области. Следовательно, в целом организме адаптационно-трофические влияния могут осуществляться рефлекторно (посредством стимуляции рецепторов чувствительных путей), а также и путем непосредственного раздражения гипоталамических центров, нейроны которых могут возбуждаться образуемыми местно или приносимыми с кровью биологически активными веществами. Таким образом, адаптационно-трофическое влияние симпатической части автономной нервной системы, не являясь пусковым, модулирует функциональную активность того или иного органа — рецепцию, проведение возбуждения, медиацию, сокращение, секрецию и др. и приспосабливает его к потребностям организма.
35. Участие вегетативной нервной системы в формировании целостных поведенческих реакций.
а) Ганглионарная передача нервных импульсов. Преганглионарные нейроны симпатической и парасимпатической систем — холинергические: при образовании аксодендритических синапсов с ганглионарными клетками из этих нейронов высвобождается ацетилхолин (АХ). Рецепторы на поверхности ганглионарных клеток называют никотиновыми, так как их возбуждение может происходить при местном действии никотина.
б) Передача нервных импульсов в нейроэффекторном синапсе. Постганглионарные нервные волокна симпатической и парасимпатической систем образуют нейроэффекторные соединения (синапсы)с эффекторными тканями (тканями-мишенями). За высвобождение нейромедиаторов отвечают многочисленные пресинаптические утолщения, расположенные по ходу нервных волокон.
Главный нейромедиатор в симпатических нейроэффекторных соединениях — норадреналин (норэпинефрин), который высвобождается из гранулярных везикул. Постганглионарные симпатические волокна — преимущественно адренергические; исключение составляют холинергические волокна, отвечающие за симпатическую иннервацию расположенных по всему телу эккриновых потовых желез. Главный нейромедиатор в парасимпатических нейроэффекторных соединениях — ацетилхолин (АХ). Постганглионарные парасимпатические волокна преимущественно холинергические.
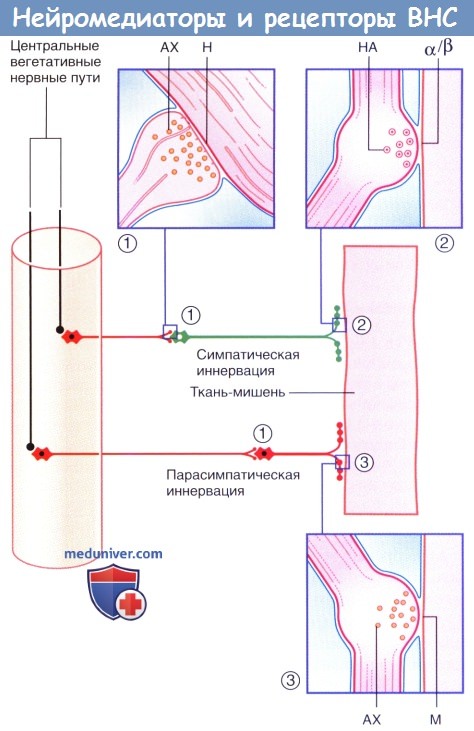
Нейромедиаторы и рецепторы вегетативной нервной системы.
(1) Аксодендритические синапсы с никотиновыми рецепторами.
(2) Нейроэффекторные синапсы с адренергическими рецепторами.
(3) Нейроэффекторные синапсы с мускариновыми рецепторами. Ганглионарные нейроны и постганглионарные волокна выделены красным цветом.
АХ — ацетилхолин; М — мускариновые рецепторы; Н — никотиновые рецепторы; НА — норадреналин.
в) Рецепторы в нейрозффекторном синапсе. Физиологические эффекты вегетативной нервной системы (ВНС) зависят от вида рецепторов на постсинаптической мембране (плазматической мембране эффекторных клеток). На высвобождение нейромедиаторов также влияют рецепторы пресинаптической мембраны (аксолеммы).
- Синаптические рецепторы симпатической системы (адренорецепторы). Для норадреналина существуют два вида а-адренорецепторов и два вида β-адренорецепторов.
1. Постсинаптические α1-адренорецеторы, активация которых вызывает сокращение гладких мышц мелких периферических артерий и крупных артериол, дилататора зрачка, а также мышц семявыносящего протока, сфинктеров ЖКТ и шейки мочевого пузыря.
2. Пресинаптические α2-адренорецеторы локализуются как на симпатических, так и на парасимпатических нервных окончаниях и ингибируют в них высвобождение нейромедиаторов. Пресинаптические α2-адренорецеторы симпатической системы называют ауторецепторами.
3. Постсинаптические адренорецепторы, возбуждение которых приводит к увеличению пейсмекерной активности клеток сердца, а также к повышению силы сердечных (желудочковых) сокращений. При резком падении артериального давления за счет активации симпатических β1-адренорецепторов юкстагломерулярных клеток почек происходит секреция ренина, который, в свою очередь, вызывает секрецию мощного вазоконстриктора ангиотензина II.
4. β2-Адренорецепторы, которые реагируют как на норадреналин, так и на адреналин (эпинефрин).
Активация постсинаптических β2-адренорецепторов вызывает расслабление гладких мышц, наиболее ярко выраженное в трахеобронхиальном дереве и мышцах глаза, участвующих в аккомодации. Часть постсинаптических β2-адренорецепторов располагается на поверхности гепатоцитов. При возрастании энергетических затрат организма их активация вызывает распад гликогена, необходимый для поддержания уровня глюкозы в крови.
Пресинаптические β2-адренорецепторы адренергических нервных окончаний отвечают за высвобождение норадреналина.
В симпатических нервных окончаниях большая часть высвобождаемого норадреналина подвергается обратному захвату моноаминным ферментным насосом. После обратного захвата часть норадреналина разрушается митохондриальным ферментом — моноаминоксидазой (МАО). Влияние лекарственных средств на симпатическую систему подробно рассмотрено в блоке клинической информации ниже.
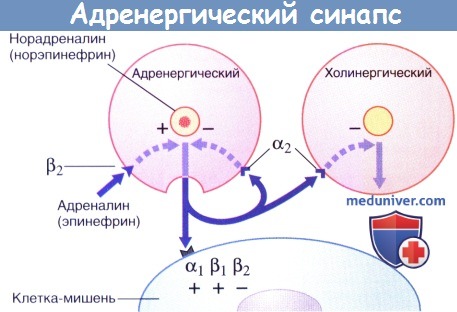
Передача возбуждения в адренергическом нейроэффекторном синапсе.
Секреция норадреналина стимулируется адреналином и ингибируется путем активации пресинаптических α2-рецепторов (данные рецепторы активируются и на пресинаптической мембране близлежащих парасимпатических синапсов, где также ингибируют секрецию).
- Синаптические рецепторы парасимпатической системы. Парасимпатические синаптические рецепторы называют мускариновыми, так как их активацию, помимо АХ, может вызывать другое химическое вещество — мускарин. Стимуляция парасимпатической системы приводит к следующим М-холинергическим эффектам.
• Замедление (в ответ на стимуляцию блуждающего нерва) частоты сердечных сокращений, а также уменьшение силы желудочковых сокращений.
• Сокращение гладких мышц, которое обусловливает такие процессы, как перистальтика кишечника, опустошение мочевого пузыря, а также рефлекс аккомодации при взгляде на близкое расстояние.
• Секреция желез.
Помимо представленных выше эффектов, пресинаптические мускариновые рецепторы симпатических нервных окончаний также отвечают за ингибирование высвобождения норадреналина.
Влияние лекарственных средств на парасимпатическую систему подробно рассмотрено в Блоке клинической информации. Лекарственные средства, обладающие мускариноподобным действием, называют холинергическими (холиномиметическими). Лекарственные средства, блокирующие доступ АХ к постсинаптической мембране, называют антихолинергическими (холинолитическими).
Главный фактор, который необходимо учитывать при назначении препаратов, активирующих или подавляющих деятельность симпатической и парасимпатической систем,— наличие рецепторов к этим лекарственным веществам α-, β- и мускариновых) в ЦНС. Так, в частности, применение данных лекарственных средств в психиатрии обусловлено их действием на центральные, а не на периферические рецепторы.
Расположение парасимпатических интрамуральных ганглиев в сердце. 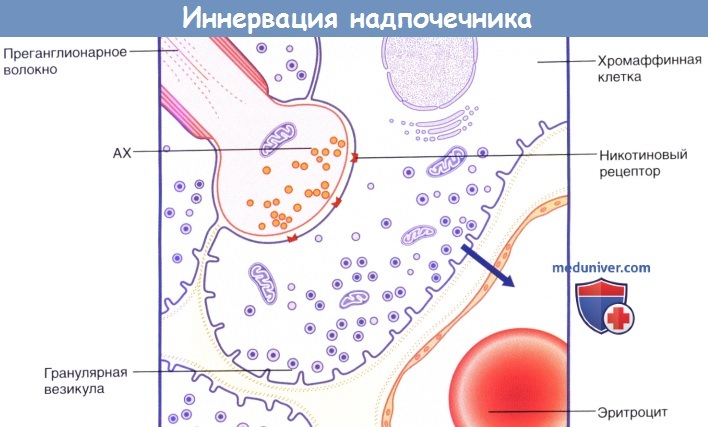
Синаптическое соединение преганглионарного волокна грудного внутренностного нерва и хромаффинной клетки мозгового вещества надпочечников.
Активация никотиновых рецепторов ацетилхолином (АХ).
8% клеток содержат крупные гранулярные везикулы (представлены на рисунке), высвобождающие адреналин; его секрекция в капиллярное русло обозначена стрелкой.
20% клеток содержат маленькие гранулярные везикулы, высвобождающие норадреналин. 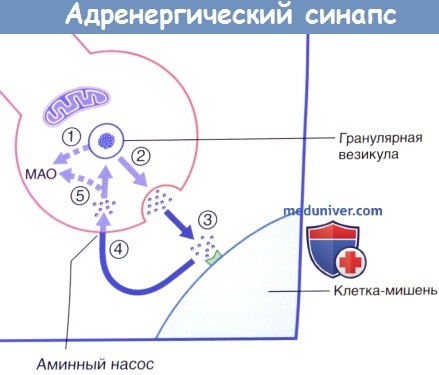
Высвобождение и обратный захват нейромедиатора в адренергическом нервном окончании.
МАО — моноаминоксидаза. 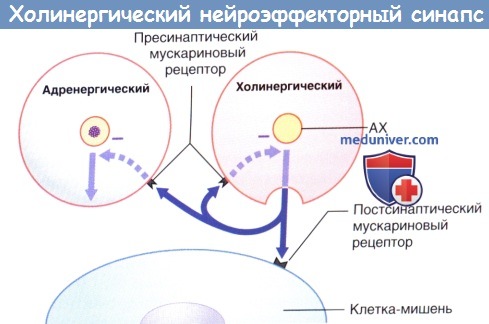
Передача возбуждения в холинергическом нейроэффекторном синапсе.
Избыточная секреция ацетилхолина (АХ) ингибируется путем активации пресинаптических мускариновых рецепторов (данные рецепторы активируются и на пресинаптической мембране близлежащих симпатических синапсов, где также ингибируют секрецию).
г) Другие типы нейронов. И в симпатическом, и в парасимпатическом отделах ВНС часть нейронов представлена неадренергическими, нехолинергическими нейронами (NANC-нейронами). Находящиеся в симпатических ганглиях небольшие вставочные нейроны отвечают за высвобождение дофамина — предшественника норадреналина. Часть дофамина выделяется в капиллярное русло, другая часть связывается с дофаминовыми рецепторами на основных (адренергических) нейронах, оказывая на них слабое тормозное действие.
Самое большое количество NANC-нейронов находится в области ганглионарных клеток стенки ЖКТ и в тазовых ганглиях. Эти нейроны содержат более 50 разнообразных белковых субстанций, находящихся в несвязанном состоянии или в различных комбинациях. Функция большей части этих веществ — регуляторная: действуя на пресинаптическую или постсинаптическую мембрану, они влияют на продолжительность работы основных нейромедиаторов. Другие белковые субстанции (вещества, выделяющиеся совместно с ацетилхолином (АХ)) представляют собой комедиаторы (сопутствующие медиаторы).
Редактор: Искандер Милевски. Дата публикации: 14.11.2018
Преганглионарные синапсы
Преганглионарные синапсы образуются нервными отростками вставочных нейронов вегетативных центров на нервных клетках вегетативных ганглиев. Число нейронов в вегетативном ганглии очень велико и в 2-30 раз превышает количество входящих в ганглий преганглионарных проводников. Поэтому каждое преганглионарное волокно ветвится и образует синапсы на нескольких нейронах ганглия. В то же время, на каждом нейроне ганглия имеются многочисленные синапсы разных преганглионарных волокон. Эти особенности обеспечивают нейронам ганглиев высокую способность к пространственной и временной суммации возбуждений.
Преганглионарные синапсы отличают три особенности:
1) значительная синаптическая задержка проведения, примерно в 5 раз продолжительнее, чем в центральных синапсах,
2) существенно большая длительность ВПСП,
3) наличие выраженной и продолжительной следовой гиперполяризации нейронов ганглия. Благодаря этим особенностям преганглионарные синапсы обладают невысокой лабильностью и обеспечивают трансформацию ритма возбуждений с частотой импульсации в постганглионарном волокне не более 15/с.
Медиатором во всех преганглионарных синапсах и симпатического, и парасимпатического отделов вегетативной нервной системы является ацетилхолин. Химические клеточные рецепторы постсинаптической мембраны, связывающие ацетилхолин, называют холинорецепторами и относят в преганглионарных синапсах к никотиночувствительным, так как они активируются никотином (Н-холинорецепторы). Специфическими блокаторами этих рецепторов являются кураре и курареподобные вещества (бензогексоний, дитилин и др.), входящие В группу ганглиоблокаторов. Кроме основных, участвующих в передаче возбуждения Н-холинорецепторов, преганглионарные синапсы имеют и М-холинорецепторы (активируются алкалоидом мускарином — мускариночувствительные), роль которых, повидимому, сводится к регуляции освобождения медиатора и чувствительности Н-холинорецепторов
Постганглионарные или периферические синапсы
Постганглионарные или периферические синапсы, образуемые эфферентным проводником на эффекторе, отличаются у двух описываемых отделов вегетативной нервной системы.
Симпатические синапсы образуются не только в области многочисленных концевых ветвлений симпатического нерва, как у всех других нервных волокон, но и у мембран варикозов — многочисленных расширений периферических участков симпатических волокон в области иннервируемых тканей. Варикозы также содержат синаптические пузырьки с медиатором, хотя и в меньших концентрациях, чем терминальные окончания.
Основным медиатором симпатических синапсов является норадреналин и такие синапсы называют адренергическими. Рецепторы, связывающие адренергический медиатор получили название адренорецепторов. Различают два типа адренорецепторов — альфа и бета, каждый из которых делят на два подтипа — 1 и 2. Небольшая часть симпатических синапсов использует медиатор ацетилхолин и такие синапсы называют холинергическими, а рецепторы — холинорецепторами. Холинергические синапсы симпатической нервной системы обнаружены в потовых железах. В адренергических синапсах кроме норадреналина в существенно меньших количествах содержатся адреналин и дофамин, также относящиеся к катехоламинам, поэтому медиаторное вещество в виде смеси трех соединений раньше называли симпатином.
Синтез норадреналина из аминокислоты тирозина с помощью трех ферментов — тирозингидроксилазы, ДОФА-декарбоксилазы и дофамин-бета-гидроксилазы — происходит во всех частях постганглионарного нейрона: его теле, аксоне, варикозах и терминальных синаптических окончаниях. Однако из тела с током аксоплазмы поступает меньше 1% норадреналиш, основная же часть медиатора синтезируется в периферических отделах аксона и хранится и гранулах синаптических пузырьков. Содержащийся в гранулах норадреналин находится в двух фондах или пулах (запасных формах) — стабильном или резервном (85-90%) и лабильном, мобилизуемом в синаптическую щель при передаче возбуждения. Норадреналин лабильного фонда в случае необходимости медленно пополняется из стабильного пула. Пополнение запасов норадреналина, кроме процессов синтеза, осуществляется мощным обратным его захватом из синаптической щели пресинаптической мембраной (до 50% выделенного в синаптическую щель количества), после чего захваченный медиатор частично поступает в пузырьки, а не попавший в пузырьки — разрушается ферментом моноаминоксидазой (МАО).
Освобождение медиатора в синаптическую щель происходит квантами под влиянием импульса возбуждения, при этом число квантов пропорционально частоте нервных импульсов. Процесс высвобождения медиатора протекает с помощью экзоцитоза и является Са-зависимым.
Выделение норадреналина в синаптическую щель регулируется несколькими специальными механизмами:
1) связывание норадреналина в синаптической щели с альфа-2-адренорецепторами пресинаптической мембраны (рис.3.10), что играет роль отрицательной обратной связи и угнетает освобождение медиатора;
2) связывание норадреналина с пресинаптическими бета-адренорецепторами, что играет роль положительной обратной связи и усиливает освобождение медиатора.
При этом, если порции освобождающегося норадреналина небольшие, то медиатор взаимодействует с бета-адренорецепторами, что повышает его освобождение, а при высоких концентрациях медиатор связывается с альфа-2
адренорецептором, что подавляет его дальнейшее освобождение;
3) образование клетками эффектора и выделение в синаптическую щель простагландинов группы Е, подавляющих освобождение медиатора через пресинаптическую мембрану;
4) поступление в синаптическую щель адренергического синапса из рядом расположенного холинергического синапса ацетилхолина, связывающегося с М-холинорецептором пресинаптической мембраны и вызывающего подавление высвобождение норадреналина.
Судьба выделившегося в синаптическую щель медиатора зависит от четырех процессов:
1) связывания с рецепторами пост- и пре-синаптических мембран,
2) обратного захвата пресинаптической мембраной,
3) разрушения в области рецепторов постсинаптической мембраны с помощью фермента катехол- О-метилтрансферазы (КОМТ),
4) диффузии из синаптической щели в кровоток, откуда норадреналин активно захватывается клетками многих тканей.
Диффундируя к постсинаптической мембране, норадреналин связывается с находящимися на ней адренорецепторами двух типов — альфа-1 и бета, образуя медиатор-рецепторный комплекс (рис.3.10).

Рис.3.10. Симпатический синапс и его регуляция.
Количество альфа-1 и бета-адренорецепторов в различных тканях неодинаково, например, в гладких мышцах артериальных сосудов внутренних органов преобладают альфа-адренорецепторы, а клетках миокарда — бета-адренорецепторы. Активация медиатором альфа- 1-адренорецепторов приводит к деполяризации и формированию ВПСП, более полого, низкоамплитудного и длительного, чем ВПСП нервных клеток и ПКП скелетных мышц. Стимуляция альфа-адренорецепторов вызывает также сдвиг метаболизма в мембране клеток и образование специфических молекул, называемых вторичными посредниками медиаторного эффекта. Вторичными посредниками стимуляции альфа-адренорецепторов являются инозитол-3-фосфат и ионизированный кальций. Более подробно системы вторичных посредников будут рассмотрены в разделе, посвященном гуморальной регуляции функций.
Бета-адренорецепторы, также как и альфа-, делят на 2 подтипа: бета-1 и бета-2.
Бета-1-адренорецепторы находятся в сердечной мышце и их стимуляция обеспечивает активацию основных физиологических свойств миокарда (автоматии, возбудимости, проводимости и сократимости).
Бета-2-адренорецепторы расположены в гладких мышцах артериальных сосудов, особенно скелетных мышц, коронарных артерий, бронхов, матки, мочевого пузыря и их стимуляция вызывает тормозной эффект в виде расслабления гладких мышц.
Хотя при этом и происходит гиперполяризация постсинаптической мембраны, выявить ТПСП не удается из-за очень медленного процесса и крайне низких амплитуд гиперполяризации. Стимуляция бета-адренорецепторов приводит в действие другую систему вторичных посредников — аденилатциклаза-цАМФ, причем считается, что бета-адренорецептор либо связан с аденилатциклазой, либо вообще является этим белком-ферментом.
Симпатическая нервная система является важнейшим регулятором обмена веществ в организме. С метаболическими эффектами симпатической нервной системы связано ее трофическое действие на ткани. Классическим экспериментальным подтверждением трофического влияния симпатической нервной системы является феномен Орбели — Гинецинского, суть которого состоит в следующем. Регистрируется амплитуда сокращений икроножной мышцы лягушки при раздражении иннервирующих ее передних корешков спинного мозга. Постепенно развивается утомление и амплитуда сокращений падает. Если в этот момент произвести раздражение симпатического пограничного ствола в этой области, то амплитуда сокращений восстанавливается, т.е. утомление исчезает
Парасимпатические постганглионарные или периферические синапсы используют в качестве медиатора ацетилхолин, который находится в аксоплазме и синаптических пузырьках пресинаптических терминалей в трех основных пулах или фондах. Это,
во-первых, стабильный, прочно связанный с белком, не готовый к освобождению пул медиатора;
во-вторых, мобилизационный, менее прочно связанный и пригодный к освобождению, пул;
в-третьих, готовый к освобождению спонтанно или активно выделяемый пул. В пресинаптическом окончании постоянно происходит перемещение пулов с целью пополнения активного пула, причем этот процесс осуществляется и путем продвижения синаптических пузырьков к пресинаптической мембране, так как медиатор активного пула содержится в тех пузырьках, которые непосредственно прилежат к мембране. Освобождение медиатора происходит квантами, спонтанное выделение единичных квантов сменяется активным при поступлении импульсов возбуждения, деполяризующих пресинаптическую мембрану. Процесс освобождения квантов медиатора, также как и в других синапсах, является кальций-зависимым.
Регуляция освобождения ацетилхолина в синаптическую щель обеспечивается следующими механизмами:
1) Связыванием ацетилхолина с М-холинорецепторами пресинаптической мембраны, что оказывает тормозящее влияние на выход ацетилхолина — отрицательная обратная связь;
2) Связыванием ацетилхолина с Н-холинорецептором, что усиливает освобождение медиатора — положительная обратная связь;
3) Поступлением в синаптическую щель парасимпатического синапса норадреналина из рядом располагающегося симпатического синапса, что оказывает тормозной эффект на освобождение ацетилхолина;
4) Выделением в синаптическую шель под влиянием ацетилхолина из постсинаптической клетки большого числа молекул АТФ, которые связываются с пуринергическими рецепторами пресинаптической мембраны и подавляют освобождение медиатора — механизм, получивший название ретро-ингибирование. (рис.3.11)

Рис.3.11. Парасимпатический синапс и его регуляция.
Выделившийся в синаптическую щель ацетилхолин удаляется из нее несколькими путями:
Во-первых, часть медиатора связывается с холинорецепторами пост- и пресинаптической мембраны;
во-вторых, медиатор разрушается ацетилхолинэстеразой с образованием холина и уксусной кислоты, которые подвергаются обратному захвату пресинаптической мембраной и вновь используются для синтеза ацетилхолина;
в-третьих, медиатор путем диффузии выносится в межклеточное пространство и кровь, причем этот процесс происходит после связывания медиатора с рецептором. При удалении медиатора последним путем инактивируется почти половина выделившегося ацетилхолина.
На постсинаптической мембране ацетилхолин связывается с холинорецепторами, относящимися к М (мускариночувствительному) типу.
Образование на мембране медиатор-реиепторного комплекса приводит к общим для разных видов клеток реакциям:
во-первых, к активации рецепторуправляемых ионных каналов и изменению заряда мембраны;
во-вторых, к активации систем вторичных посредников в клетках.
В гладкомышечных и секреторных клетках желудочно-кишечного тракта, мочевого пузыря и мочеточника, бронхов, коронарных и легочных сосудов комплекс ацетилхолин-М-холино-рецептор активирует Na-каналы, приводит к деполяризации и формированию ВПСП, вследствие чего клетки возбуждаются и происходит сокращение гладких мышц или секреция пищеварительных соков. Этому же эффекту способствует активация вторичных посредников — инозитол-три-фосфата и ионизированного кальция. В то же время в клетках проводящей системы сердца, гладких мышцах сосудов половых органов комплекс ацетилхолин-М-холинорецептор активирует К-каналы и выходящий ток К+, приводя к гиперполяризации и тормозным эффектам — снижению автоматии, проводимости и возбудимости в миокарде, расширению артерий половых органов. Одновременно в этих клетках активируется система вторичных посредников — гуанилатциклаза-циклический гуанозинмонофосфат. Разнонаправленность эффектов парасимпатической регуляции при образовании на мембранах разных клеток комплекса ацетилхолин-М-холинорецептор дает основание предполагать наличие на постсинаптической мембране постганглионарных синапсов двух типов М-холинорецепторов, подобно типам адренорецепторов описанным выше. Вместе с тем, все М-холинорецепторы блокируются атропином, что снимает как парасимпатическую стимуляцию сокращения гладких мышц, так и парасимпатическое торможение деятельности сердца.
Эффективность синаптической передачи зависит от количества активных рецепторов на постсинаптической мембране, что отражает функции эффекторной клетки, синтезирующей мембранные рецепторы. Клетка эффектора регулирует число мембранных рецепторов в зависимости от интенсивности работы синапса, т.е. выделения в нем медиатора. Так, при перерезке вегетативного нерва (прекращении выделения медиатора) чувствительность иннервируемой им ткани к соответствующему медиатору возрастает из-за увеличения числа мембранных рецепторов, способных связывать-медиатор. Повышение чувствительности денервированных структур или сенситизация ткани является примером саморегуляции на уровне эффектора.
Взаимосвязи симпатической и парасимпатической регуляции функций
Поскольку большинство эффектов симпатической и парасимпатической нервной регуляции являются противоположными, их взаимоотношения характеризуют иногда как антагонистические. Вместе с тем, существующие взаимосвязи между высшими вегетативными центрами и даже на уровне постганглионарных синапсов в тканях, получающих двойную иннервацию, позволяют применять понятие о реципрокной регуляции. Примером реципрокных взаимоотношений на уровне эффектора является акцентированный антагонизм или взаимоусиливающее противодействие.
Читайте также: