
Поперечно полосатые мышцы нервные клетки
Поперечно- полосатые мышцы скелета состоят из множества отдельных мышечных волокон, которые расположены в общем соединительнотканном футляре и крепятся к сухожилиям, связанным со скелетом. В некоторых мышцах все волокна параллельны длинной оси мышцы – параллельно-волокнистый тип; в других мышцах они расположены косо, прикрепляясь с одной стороны к центральному сухожильному тяжу, а с другой – к наружному сухожильному футляру – перистый тип.
Основной особенностью поперечно-полосатого мышечного волокна является наличие в его саркоплазме массы тонких (диаметром 1 мкм) нитей – миофибрилл, расположенных вдоль длинной оси волокна. Миофибриллы состоят из чередующихся светлых (изотропных – I-дисков) и темных (анизотропных – А дисков) участков, причем в массе соседних миофибрилл у поперечно-полосатых волокон одноименные диски расположены на одном уровне (поперечном сечении). Последнее придает регулярную поперечную исчерченность (полосатость) всему мышечному волокну. Комплекс из одного темного (А) и двух прилежащих светлых (I) дисков, ограниченный тонкими Z-линиями, называется саркомером. Миофибриллы, точнее – их саркомеры – являются сократительным аппаратом мышцы.
Таким образом, сократительное мышечное волокно, или миом, включает следующие компоненты:
сократительный аппарат – систему миофибрилл;
трофический аппарат с типичными для мышечного волокна органеллами – митохондриями (или саркосомами), пластинчатым комплексом Гольджи и слаборазвитой эндоплазматической сетью;
специфический мембранный аппарат – соединительнотканная сумка – саркоплазматическая сеть и трубчал=тый элемент, составляющий Т-систему;
опорный аппарат – соединительнотканная сумка волокна и поперечные перегородки – линии или Z-полоски;
нервный аппарат – мионевральные синапсы и чувствительные (рецепторные) элементы мышцы – тельца Гольджи, мышечные веретена, тела Паччини.
Расчленение мышечного волокна на пять структурно-функциональных подразделений условно, поскольку мышечное волокно представляет собой целостную структурно-функциональную единицу мышечной ткани.
Множественный тип иннервации волокон представлен во внешних глазных мышцах, где имеются также и одиночно иннервированные мышечные волокна. Потенциалы действия в них не генерируются из-за отсутствия в их мембране электровозбудимых натриевых каналов. В них происходит электротоническое распространение деполяризации из синаптических окончаний по всему волокну, что является необходимым для повсеместного запуска сократительного акта. Сократительный акт здесь более медленный, чем в волокнах с одиночной иннервацией, поэтому такие волокна называют медленными.
Структура миофибрилл и её изменения при сокращении.
Современные представления о структуре миофибриллярного (сократительного) аппарата основываются на исследованиях структуры мышечного волокна при помощи электронной микроскопии, рентгеноструктурного анализа в сочетании с гистохимическими методами.
Установлено: каждая миофибрилла мышечного волокна диаметром 1 мкм состоит в среднем из 2500 протофибрилл, представляющих собой удлиненные полимеризованные молекулы сократительных белков – миозина и актина, которые называются протофибриллами. Миозиновые протофибриллы, или как их ещё называют, нити, вдвое толще актиновых (их диаметр около 10 нм). В состоянии покоя в мышце нити расположены так, что тонкие актиновые нити входят своими концами в промежутки между короткими и толстыми миозиновыми протофибриллами. Благодаря этому І-диски состоят только из актиновых нитей, а А-диски – из миозиновых и небольшого количества актиновых нитей (в местах захода актиновых нитей в промежутки между миозиновыми).
Светлая полоса Н представляет собой узкую зону в диске А, свободную от актиновых нитей. Мембрана Z, проходя через середину диска І, скрепляет между собой эти нити.
МЕХАНИЗМ СОКРАЩЕНИЯ МЫШЕЧНОГО ВОЛОКНА.
Основное положение теории скользящих нитей – во время скольжения (сокращения) сами актиновые и миозиновые нити не укорачиваются, так как ширина диска А остается при сокращении постоянной, а І-диски и Н-зоны становятся более узкими или совсем исчезают. Длина протофибрилл не изменяется и при растяжении мышцы. Вместо этого пучки тонких нитей, скользя, выходят из промежутков между толстыми нитями, так что степень их перекрытия уменьшается
Во время сокращения каждая головка миозина, или поперечный мостик, связывает миозиновую протофибриллу с актиновой. Наклоны головок создают объединенное усилие, и происходит скольжение (гребок), продвигающий актиновую нить к середине саркомера. Биполярная организация молекул миозина в двух половинах саркомера уже обеспечивает возможность скольжения актиновых нитей в противоположном направлении в левой и правой половине саркомера.
Когда мышца расслабляется, головки миозина отходят от актиновых нитей. Поскольку актиновые и миозиновые нити могут легко скользить относительно друг друга, сопротивление растяжению в расслабленных мышцах оказывается низким, поэтому удлинение мышцы во время расслабления является пассивным.
ЭТАПЫ ГЕНЕРАЦИИ СОКРАЩЕНИЯ.
Хранение и высвобождение ионов кальция. В состоянии расслабления мышца содержит более 1 мкмоль Са на 1 г сырого веса. Если бы соли Са не были изолированы в особых внутриклеточных хранилищах, обогащенные кальцием мышечные волокна находились бы в состоянии непрерывного сокращения. Структура внутриклеточных систем хранения кальция следующая: во многих участках мембрана мышечной клетки углубляется внутрь волокна, перпендикулярно его продольной оси, образуя трубки; эта система поперечных трубочек (Т-система) соединяется с внеклеточной средой. Перпендикулярно Т-системе, т.е. параллельно миофибриллам, расположена система продольных трубочек (истинный саркоплазматический ретикулум). Пузырьки на концах этих трубочек, терминальные цистерны, находятся очень близко к мембранам поперечной системы, образуя триады. В этих пузырьках и хранится внутриклеточный Са 2+ . В отличие от поперечной системы продольная система не соединяется с окружающей средой.
Таким образом, электромеханическое сопряжение происходит посредством распространения потенциала действия по мембранам поперечной системы внутрь клетки. При этом возбуждение быстро проникает во внутрь волокна, переходит к продольной системе и, в конечном счете, вызывает высвобождение ионов Са 2+ , которые хранятся в терминальных цистернах, во внутриклеточную жидкость около миофибрилл, что ведет к сокращению.
РОЛЬ АТФ В МЕХАНИЗМЕ МЫШЕЧНОГО СОКРАЩЕНИЯ
В процессе взаимодействия миозиновых и актиновых нитей в присутствии Са 2+ важную роль играет АТФ.
Энергия АТФ используется во время деятельности скелетной мышцы для трех основных процессов:
работы натрий-калиевого насоса, обеспечивающего поддержание постоянства градиента концентрации ионов натрия и калия по обе стороны мембраны;
процесса скольжения актиновых и миозиновых нитей, ведущего к укорочению миофибрилл;
работы кальциевого насоса, необходимого для расслабления волокна.
В соответствии с этим фермент АТФаза локализован в трех различных структурах мышечного волокна: клеточной мембране, миозиновых нитях, мембранах саркоплазматического ретикулума. АТФ гидролитически арсщепляется и, таким образом, энергетически утилизируется с помощью АТФазы – фермента миозина; причем, процесс активируется актином.
Циклическая активность поперечных мостиков, т.е. ритмическое прикрепление и отсоединение мостиков, которое обеспечивает мышечное сокращение, возможна только до тех пор, пока продолжается гидролиз АТФ, т.е. пока происходит активация АТФазы. Если расщепление АТФ заблокировать, мостики не могут повторно прикрепляться, мышца расслабляется.
После смерти содержание АТФ в мышечных клетках снижается, когда оно переходит критический уровень, поперечные мостики оказываются устойчиво прикрепленными к актиновой нити (пока не произойдет аутолиз). В таком состоянии актиновые и миозиновые нити прочно связаны друг с другом, мышца находится в состоянии трупного окоченения.
Ресинтез АТФ осуществляется двумя основными путями:
ферментативный перенос фосфатной группы от креатинфосфата на АДФ. Ресинтез обеспечивается по этому пути в течение тысячных долей секунды, т.к. запасы КФ значительно больше в клетке, чем АТФ;
гликолитические и окислительные процессы в покоящейся и деятельной мышце – медленный ресинтез АТФ через окисление молочной и пировиноградной кислот.
Нарушение ресинтеза АТФ ядами ведет к полному исчезновению АТФ и креатинфосфата, вследствие чего кальциевый насос перестает работать. Концентрация Са 2+ в области миофибрилл значительно возрастает и мышца приходит в состояние длительного необратимого укорочения. Это состояние называется контрактурой.
Содержание
- 1 Иннервация поперечнополосатых мышц
- 1.1 Двигательные волокна
- 1.2 Чувствительные волокна
- 1.3 Вегетативные волокна
- 2 Иннервация мышц
- 3 Читайте также
Иннервация поперечнополосатых мышц [ править | править код ]
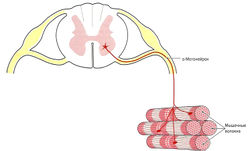
Скелетные мышцы иннервируются одним или несколькими нервами, которые проникают в мышцу в области ворот. За несколькими исключениями (лицевой нерв — n. facialis, подъязычный нерв — n. hypoglossus) все нервы являются смешанными, т. е. имеют двигательные, чувствительные и вегетативные волокна.
Различают тонкие немиелинизированные у-волокна (Ау-волокна) и толстые миелинизированные а-волокна (Аа-волокна).

Запомните: у-Волокна и а-волокна (аксоны) отходят от тела (сомы) у- и а-мотонейронов. Тела клеток расположены в ядрах черепных нервов ствола головного мозга и в передних рогах спинного мозга.
В области брюшка мышцы нерв отдает многочисленные ветви. Каждая ветвь разделяется на терминальные ветви, которые идут к эндомизию мышечных волокон (экстрафузальные волокна), причем количество иннервируемых мышечных волокон может быть различным. Терминальные ветви соединяются с сарколеммой мышечных волокон с помощью моторной концевой пластинки. Один мотонейрон может иннервировать различное количество мышечных волокон, и к каждому волокну подходит только одна концевая пластинка. Мотонейрон, его аксон со всеми ветвями и иннервируемые им волокна образуют так называемую моторную единицу (рис. 1.5).
Мышечные волокна одной моторной единицы располагаются в мышце в различных пучках. Таким образом, при возбуждении мотонейронов обеспечивается сокращение всей мышцы, а не отдельных фасцикул. Величина моторной единицы может сильно различаться: чем меньше моторная единица, тем точнее регуляция движений. Самые маленькие моторные единицы находятся в мышцах глазного яблока и пальцев (10-20 волокон), благодаря чему возможны очень точные движения. Мышцы, которым необходимо развивать большую силу (например, мышцы бедра), наоборот, имеют большие моторные единицы (несколько тысяч мышечных волокон). Внутри одной моторной единицы находятся волокна только одного типа.
Тонкие у-волокна иннервируют мышечные веретена и расположенные внутри мышечных веретен интрафузальные мышечные волокна.
Запомните: Моторная единица состоит из мотонейрона и всех иннервируемых им мышечных волокон. Чем меньше моторная единица, тем точнее и дозированнее выполняются движения в суставах.
Афферентные волокна могут либо заканчиваться свободными нервными окончаниями, либо соединяться со специфическими рецепторами — мышечными веретенами и сухожильными органами Гольджи (нервно-сухожильными веретенами) — проприорецепторами. В соединительной ткани мышц находятся и другие рецепторы — тельца Пачини, тельца Руффини и рецепторы Догеля. Свободные нервные окончания (ноцицепторы) диффузно расположены между мышечными волокнами и в соединительной ткани мышц, и в области мышечно-сухожильного соединения. Они проводят болевые импульсы от мышцы через Аδ-волокна и С-волокна к коре больших полушарий, где повышение импульсной активности от ноцицепторов воспринимается в виде чувства боли (Briigger, 2000). Мышечные веретена, лежащие вдоль волокон, регистрируют длину мышцы и скорость ее изменения. Данные проприоцептивные импульсы затем поступают через афферентные 1а-волокна в задние рога спинного мозга. Сухожильные органы Гольджи вплетаются в коллагеновые волокна сухожилий поперечнополосатых мышц. Они измеряют изменения натяжения сухожилия и проводят афферентные импульсы через lb-волокна в центральную нервную систему. Другие чувствительные волокна проводят афферентные импульсы от соединительнотканных оболочек в ЦНС.
В соединительной ткани мышц также расположено большое количество вегетативных симпатических волокон. Эти немиелинизированные эфферентные волокна иннервируют гладкие мышцы стенок кровеносных сосудов скелетной мускулатуры.





- Физиология
![]()
- История физиологии


Общий обзор мышечной системы человека
У позвоночных животных и человека различают три разных по строению группы мышц:
- поперечно-полосатые мышцы скелета;
- поперечно-полосатая мышца сердца;
- гладкие мышцы внутренних органов, сосудов и кожи.

Рис. 1. Виды мышц человека
Из двух видов мышечной ткани (поперечно-полосатой и гладкой) гладкая мышечная ткань находится на более низкой ступени развития и присуща низшим животным.
Гладкие мышцы образуют мышечный слой стенок желудка, кишечника, мочеточников, бронхов, кровеносных сосудов и других полых органов. Они состоят из веретенообразных мышечных волокон и не имеют поперечной исчерченности, так как миофибриллы в них расположены менее упорядоченно. В гладких мышцах отдельные клетки соединяются между собой специальными участками наружных мембран - нексусами. За счет этих контактов потенциалы действия распространяются с одного мышечного волокна на другое. Поэтому в реакцию возбуждения быстро вовлекается вся мышца.
Гладкие мышцы осуществляют движения внутренних органов, кровеносных и лимфатических сосудов. В стенках внутренних органов они, как правило, располагаются в виде двух слоев: внутреннего кольцевого и наружного продольного. В стенках артерии они формируют спиралевидные структуры.
Характерной особенностью гладких мышц является их способность к спонтанной автоматической деятельности (мышцы желудка, кишечника, желчного пузыря, мочеточников). Это свойство регулируется нервными окончаниями. Гладкие мышцы пластичны, т.е. способны сохранять приданную растяжением длину без изменения напряжения. Скелетная мышца, наоборот, обладает малой пластичностью и эту разницу легко установить в следующем опыте: если растянуть с помощью грузов и гладкую и поперечно-полосатую мышцы и снять груз, то скелетная мышца сразу же после этого укорачивается до первоначальной длины, а гладкая мышца долгое время может находиться в растянутом состоянии.
Такое свойство гладких мышц имеет большое значение для функционирования внутренних органов. Именно пластичность гладких мышц обеспечивает лишь небольшое изменение давления внутри мочевого пузыря при его наполнении.

Рис. 2. А. Волокно скелетной мышцы, клетка сердечной мышцы, гладкая мышечная клетка. Б. Саркомер скелетной мышцы. В. Строение гладкой мышцы. Г. Механограмма скелетной мышцы и мышцы сердца.
Гладким мышцам присущи те же основные свойства, что и поперечнополосатым скелетным мышцам, но и некоторые особые свойства:
- автоматия, т.е. способность сокращаться и расслабляться без внешних раздражений, а за счет возбуждений, возникающих в них самих;
- высокая чувствительность к химическим раздражителям;
- выраженная пластичность;
- сокращение в ответ на быстрое растяжение.
Сокращение и расслабление гладких мышц происходит медленно. Это способствует наступлению перестальтических и маятникообразных движений органов пищеварительного тракта, что приводит к перемещению пищевого комка. Длительное сокращение гладких мышц необходимо в сфинктерах полых органов и препятствует выходу содержимого: желчи в желчном пузыре, мочи в мочевом пузыре. Сокращение гладкомышечных волокон совершается независимо от нашего желания, под воздействием внутренних, не подчиненных сознанию причин.
Поперечно-полосатые мышцы располагаются на костях скелета и сокращением приводят в движение отдельные суставы и все тело. Скелетные мышцы образуют тело, или сому, поэтому их еще называют соматическими, а иннервирующую их систему — соматической нервной системой.
Благодаря деятельности скелетной мускулатуры осуществляется передвижение тела в пространстве, разнообразная работа конечностей, расширение грудной клетки при дыхании, движение головы и позвоночника, жевание, мимика лица. Насчитывается более 400 мышц. Общая масса мышц составляет 40% веса. Обычно средняя часть мышцы состоит из мышечной ткани и образует брюшко. Концы мышц — сухожилия построены из плотной соединительной ткани; они соединяются с костями при помощи надкостницы, но могут прикрепляться и к другой мышце, и к соединительному слою кожи. В мышце мышечные и сухожильные волокна объединяются в пучки при помощи рыхлой соединительной ткани. Между пучками располагаются нервы и кровеносные сосуды. Сила мышцы пропорциональна количеству волокон, составляющих брюшко мышцы.

Рис. 3. Функции мышечной ткани
Некоторые мышцы проходят только через один сустав и при сокращении приводят его в движение — односуставные мышцы. Другие мышцы проходят через два или несколько суставов — многосуставные, они производят движение в нескольких суставах.
При сокращении концы мышцы, прикрепленные к костям, приближаются друг к другу, а размеры мышцы (длина) уменьшается. Кости, соединенные суставами, действуют как рычаги.
Изменяя положение костных рычагов, мышцы действуют на суставы. При этом каждая мышца влияет на сустав только в одном направлении. У одноосного сустава (цилиндрический, блоковидный) имеются две действующие на него мышцы или группы мышц, являющиеся антагонистами: одна мышца — сгибатель, другая — разгибатель. В то же время на каждый сустав в одном направлении действует, как правило, две мышцы и более, являющиеся синергистами (синергизм — совместное действие).
У двуосного сустава (эллипсоидный, мышелковый, седловидный) мышцы группируются соответственно двум его осям, вокруг которых совершаются движения. К шаровидному суставу, имеющему три оси движения (многоосный сустав), мышцы прилежат со всех сторон. Так, например, в плечевом суставе имеются мышцы-сгибатели и разгибатели (движения вокруг фронтальной оси), отводящие и приводящие (сагиттальная ось) и вращатели вокруг продольной оси, кнутри и кнаружи. Различают три вида работы мышц: преодолевающую, уступающую и удерживающую.
Если благодаря сокращению мышцы меняется положение части тела, то преодолевается сила сопротивления, т.е. выполняется преодолевающая работа. Работа, при которой сила мышцы уступает действию силы тяжести и удерживаемого груза, называется уступающей. В этом случае мышца функционирует, однако она не укорачивается, а удлиняется, например, когда невозможно поднять или удержать на весу тело, имеющее большую массу. При большом усилии мышц приходится опустить это тело на какую-нибудь поверхность.
Удерживающая работа выполняется благодаря сокращению мышц, тело или груз удерживается в определенном положении без перемещения в пространстве, например человек держит груз, не двигаясь. При этом мышцы сокращаются без изменения длины. Сила сокращения мышц уравновешивает массу тела и груза.
Когда мышца, сокращаясь, перемешает тело или его части в пространстве, они выполняют преодолевающую или уступающую работу, которая является динамической. Статистической является удерживающая работа, при которой не происходит движений всего тела или его части. Режим, при котором мышца может свободно укорачиваться, называется изотоническим (не происходит изменения напряжения мышцы и меняется только ее длина). Режим, при котором мышца не может укоротиться, называется изометрическим — меняется только напряжение мышечных волокон.

Рис. 4. Мышцы человека
Скелетные мышцы состоят из большого числа мышечных волокон, которые объединяются в мышечные пучки.
В одном пучке содержится 20-60 волокон. Мышечные волокна представляют собой клетки цилиндрической формы длиной 10-12 см и диаметром 10-100 мкм.
Каждое мышечное волокно имеет оболочку (сарколемму) и цитоплазму (саркоплазму). В саркоплазме находятся все компоненты животной клетки и вдоль оси мышечного волокна располагаются тонкие нити - миофибриллы, Каждая миофибрилла состоит из протофибрилл, в состав которых вкючены нити белков миозина и актина, являющихся сократительным аппаратом мышечного волокна. Миофибриллы разделены между собой перегородками, которые называются Z-мембранами, на участки - саркомеры. На обоих концах саркомеров к Z-мембране прикреплены тонкие актиновые нити, а в середине расположены толстые миозиновые нити. Нити актина своими концами частично входят между миозиновыми нитями. В световом микроскопе нити миозина выглядят в виде светлой полоски в темном диске. При электронной микроскопии скелетные мышцы выглядят исчерченными (поперечно-полосатыми).

Рис. 5. Поперечные мостики: Ак — актин; Мз — миозин; Гл — головка; Ш — шейка
На боковых сторонах миозиновой нити имеются выступы, получившие название поперечных мостиков (рис. 5), которые расположены под углом 120° по отношению к оси миозиновой нити. Актиновые филаменты выглядят в виде двойной нити, закрученной в двойную спираль. В продольных бороздках актиновой спирали находятся нити белка тропомиозина, к которым присоединен белок тропонин. В состоянии покоя молекулы белка тропомиозина расположены таким образом, чтобы предотвращать прикрепление поперечных мостиков миозина к актиновым нитям.

Рис. 6. А — организация цилиндрических волокон в скелетной мышце, прикрепленной к костям сухожилиями. Б — структурная организация филаментов в волокне скелетной мышцы, создающая картину поперечных полос.

Рис. 7. Строение актина и миозина
Во многих местах поверхностная мембрана углубляется в виде микротрубок внутрь волокна, перпендикулярно его продольной оси, образуя систему поперечных трубочек (Т-система). Параллельно миофибриллам и перпендикулярно поперечным трубочкам между миофибрилл расположена система продольных трубочек (саркоплазматический ретикулум). Концевые расширения этих трубочек - терминальные цистерны - подходят очень близко к поперечным трубочкам, образуя совместно с ними так называемые триады. В цистернах сосредоточено основное количество внутриклеточного кальция.
Мышечная ткань состоит из клеток, называемых мышечными волокнами. Снаружи волокно окружено оболочкой — сарколеммой. Внутри сарколеммы содержится цитоплазма (саркоплазма), содержащая ядра и митохондрии. В ней содержится огромное количество сократительных элементов, называемых миофибриллами. Миофибриллы проходят от одного конца мышечного волокна до другого. Они существуют сравнительно короткий срок — около 30 суток, после чего и происходит их полная смена. В мышцах идет интенсивный синтез белка, необходимый для образования новых миофибрилл.
Мышечное волокно содержит большое количество ядер, которые располагаются непосредственно под сарколеммой, поскольку основная часть мышечного волокна занята миофибриллами. Именно наличие большого числа ядер обеспечивает синтез новых миофибрилл. Такая быстрая смена миофибрилл обеспечивает высокую надежность физиологических функций мышечной ткани.

Рис. 7. А — схема организации саркоплазматического ретикулума, поперечных трубочек и миофибрилл. Б — схема анатомической структуры поперечных трубочек и саркоплазматического ретикулума в индивидуальном волокне скелетной мышцы. В — роль саркоплазматического ретикулума в механизме сокращения скелетной мышцы
Каждая миофибрилла состоит из правильно чередующихся светлых и темных участков. Эти участки, обладая разными оптическими свойствами, создают поперечную исчерченность мышечной ткани.
В скелетной мышце сокращение вызывается поступлением к ней импульса по нерву. Передача нервного импульса с нерва на мышцу осуществляется через нервно-мышечный синапс (контакт).
Одиночный нервный импульс, или однократное раздражение, приводит к элементарному сократительному акту — одиночному сокращению. Начало сокращения не совпадает с моментом нанесения раздражения, поскольку существует скрытый, или латентный, период (интервал между нанесением раздражения и началом сокращения мышцы). В этот период происходит развитие потенциала действия, активация ферментных процессов и распад АТФ. После этого начинается сокращение. Распад АТФ в мышце приводит к превращению химической энергии в механическую. Энергетические процессы всегда сопровождаются выделением тепла и тепловая энергия обычно является промежуточной между химической и механическими энергиями. В мышце же химическая энергия превращается непосредственно в механическую. Но тепло в мышце образуется и за счет укорочения мышцы, и во время ее расслабления. Тепло, образующееся в мышцах, играет большую роль в поддержании температуры тела.
В отличие от сердечной мышцы, которая обладает свойством автоматики, т.е. она способна сокращаться под влиянием импульсов, возникающих в ней самой, и в отличие от гладкой мускулатуры, также способной к сокращению без поступления сигналов извне, скелетная мышца сокращается только при поступлении к ней сигналов из ЦНС. Непосредственно сигналы к мышечным волокнам поступают по аксонам двигательных клеток, расположенным в передних рогах серого вещества спинного мозга (мотонейронам).
Скелетные мышцы в отличие от гладких способны совершать произвольные быстрые сокращения и производить этим значительную работу. Рабочим элементом мышцы является мышечное волокно. Типичное мышечное волокно представляет собой структуры с несколькими ядрами, отодвинутыми на периферию массой сократительных миофибрилл.
Мышечные волокна обладают тремя основными свойствами:
- возбудимостью — способностью отвечать на действия раздражителя генерацией потенциала действия;
- проводимостью — способностью проводить волну возбуждения вдоль всего волокна в обе стороны от точки раздражения;
- сократимостью — способностью сокращаться или изменять напряжение при возбуждении.
Для скелетной мышцы характерны два основных режима сокращения — изометрический и изотонический.
Изометрический режим проявляется в том, что в мышце во время ее активности нарастает напряжение (генерируется сила), но из-за того, что оба конца мышцы фиксированы (например, при попытке поднять очень большой груз), — она не укорачивается.
Изотонический режим проявляется в том, что мышца первоначально развивает напряжение (силу), способное поднять данный груз, а потом мышца укорачивается — меняет свою длину, сохраняя напряжение, равное весу удерживаемого груза. Чисто изометрического или изотонического сокращения практически наблюдать нельзя, но существуют приемы так называемой изометрической гимнастики, когда спортсмен напрягает мышцы без изменения длины. Эти упражнения в большей мере развивают силу мышц, чем упражнения с изотоническими элементами.
Сократительный аппарат скелетной мышцы представлен миофибриллами. Каждая миофибрилла диаметром 1 мкм состоит из нескольких тысяч протофибрилл — тонких, удлиненных полимеризированных молекул белков миозина и актина. Миозиновые нити в два раза тоньше актиновых, и в состоянии покоя мышечного волокна актиновые нити свободными кольцами входят между миозиновыми нитями.
В передаче возбуждения большую роль играют ионы кальция, которые входят в межфибриллярное пространство и запускают механизм сокращения: взаимное втягивание относительно друг друга актиновых и миозиновых нитей. Втягивание нитей происходит при обязательном участии АТФ. В активных центрах, расположенных на одном из концов миозиновых нитей, АТФ расщепляется. Энергия, выделяемая при расщеплении АТФ, преобразуется в движение. В скелетных мышцах запас АТФ невелик — всего на 10 одиночных сокращений. Поэтому необходим постоянный ре- синтез АТФ, который идет тремя путями: первый — за счет запасов креатинфосфата, которые ограничены; второй — гликолитический путь при анаэробном расщеплении глюкозы, когда на одну молекулу глюкозы образуется две молекулы АТФ, но одновременно образуется молочная кислота, которая тормозит активность гликолитических ферментов, и наконец третий — аэробное окисление глюкозы и жирных кислот в цикле Кребса, совершающееся в митохондриях и образующее 38 молекул АТФ на 1 молекулу глюкозы. Последний процесс наиболее экономичный, но очень медленный. Постоянная тренировка активизирует третий путь окисления, в результате чего повышается выносливость мышц к длительным нагрузкам.
Читайте также: