
Принцип окклюзии в цнс
1. Принцип доминанты был сформулирован А. А. Ухтомским как основной принцип работы нервных центров. Согласно этому принципу для деятельности нервной системы характерно наличие в ЦНС доминирующих (господствующих) в данный период времени очагов возбуждения, в нервных центрах, которые и определяют направленность и характер функций организма в этот период. Доминантный очаг возбуждения характеризуется следующими свойствами:
• стойкостью возбуждения (инертностью), т. к. трудно подавить другим возбуждением;
• способностью к суммации субдоминантных возбуждений;
• способностью тормозить субдоминантные очаги возбуждения, в функционально различных нервных центрах.
2. Принцип пространственного облегчения. Он проявляется в том, что суммарный ответ организма при одновременном действии двух относительно слабых раздражителей будет больше суммы ответов, полученных при их раздельном действии. Причина облегчения связана с тем, что аксон афферентного нейрона в ЦНС синаптирует с группой нервных клеток, в которой выделяют центральную (пороговую) зону и периферическую (подпороговую) "кайму". Нейроны, находящиеся в центральной зоне, получают от каждого афферентного нейрона достаточное количество синаптических окончаний (например, по 2) (рис. 13) , чтобы сформировать потенциал действия. Нейрон подпороговой зоны получает от тех же нейронов меньшее число окончаний (по 1), поэтому их афферентные импульсы будут недостаточны, чтобы вызвать в нейронах "каймы" генерацию потенциалов действия, а возникает лишь подпороговое возбуждение. Вследствие этого, при раздельном раздражении афферентных нейронов 1 и 2 возникают рефлекторные реакции, суммарная выраженность которых определяется только нейронами центральной зоны (3) . Но при одновременном раздражении афферентных нейронов потенциалы действия генерируются и нейронами подпороговой зоны. Поэтому выраженность такого суммарного рефлекторного ответа будет больше. Это явление получило название центрального облегчения. Оно чаще наблюдается при действии на организм слабых раздражителей.
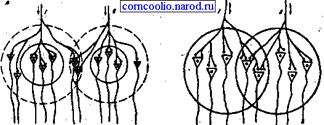
Рис. 13. Схема явления облегчения (А) и окклюзии (Б). Кругами обозначены центральные зоны (сплошная линия) и подпороговая "кайма" (пунктирная линия) популяции нейронов.
3. Принцип окклюзии. Этот принцип противоположен пространственному облегчению и он заключается в том, что два афферентных входа совместно возбуждают меньшую группу мотонейронов по сравнению с эффектами при раздельной их активации. Причина окклюзии состоит в том, что афферентные входы в силу конвергенции отчасти адресуются к одним и тем же мотонейронам, которые затормаживаются при активации обоих входов одновременно (рис. 13). Явление окклюзии проявляется в случаях применения сильных афферентных раздражении.
4. Принцип обратной связи. Процессы саморегуляции в организме аналогичны техническим, предполагающим автоматическую регуляцию процесса с использованием обратной связи. Наличие обратной связи позволяет соотнести выраженность изменений параметров системы с ее работой в целом. Связь выхода системы с ее входом с положительным коэффициентом усиления называется положительной обратной связью, а с отрицательным коэффициентом - отрицательной обратной связью. В биологических системах положительная обратная связь реализуется в основном в патологических ситуациях. Отрицательная обратная связь улучшает устойчивость системы, т. е. ее способность возвращаться к первоначальному состоянию после прекращения влияния возмущающих факторов.
Обратные связи можно подразделять по различным признакам. Например, по скорости действия - быстрая (нервная) и медленная (гуморальная) и т. д.
Можно привести множество примеров проявления эффектов обратной связи. Например, в нервной системе так осуществляется регулирование активности мотонейронов. Суть процесса заключается в том, что импульсы возбуждения, распространяющиеся по аксонам мотонейронов, достигают не только мышц, но и специализированных промежуточных нейронов (клеток Реншоу), возбуждение которых тормозит активность мотонейронов. Данный эффект известен как процесс возвратного торможения.
В качестве примера с положительной обратной связью можно привести процесс возникновения потенциала действия. Так при формировании восходящей части ПД деполяризация мембраны увеличивает ее натриевую проницаемость, которая, в свою очередь, увеличивая натриевый ток, увеличивает деполяризацию мембраны.
Велико значение механизмов обратной связи в поддержании гомеостаза. Так, например, поддержание константного уровня кровяного давления осуществляется за счет изменения импульсной активности барорецепторов сосудистых рефлексогенных зон, которые измененяют тонус вазомоторных симпатических нервов и таким образом нормализуют кровяное давление.
5. Принцип реципрокности (сочетанности, сопряженности, взаимоисключения). Он отражает характер отношений между центрами ответственными за осуществление противоположных функций (вдоха и выдоха, сгибание и разгибание конечности и т. д.). Например, активация проприорецепторов мышцы-сгибателя одновременно возбуждает мотонейроны мышцы-сгибателя и тормозит через вставочные тормозные нейроны мотонейроны мышцы-разгибателя (рис. 18). Реципрокное торможение играет важную роль в автоматической координации двигательных актов.
6. Принцип общего конечного пути. Эффекторные нейроны ЦНС (прежде всего мотонейроны спинного мозга), являясь конечными в цепочке состоящей из афферентных, промежуточных и эффекторных нейронов, могут вовлекаться в осуществление различных реакций организма возбуждениями, приходящими к ним от большого числа афферентных и промежуточных нейронов, для которых они являются конечным путем (путем от ЦНС к эффектору). Например, на мотонейронах передних рогов спинного мозга, иннервирующих мускулатуру конечности, оканчиваются волокна афферентных нейронов, нейронов пирамидного тракта и экстрапирамидной системы (ядер мозжечка, ретикулярной формации и многих других структур). Поэтому эти мотонейроны, обеспечивающие рефлекторную деятельность конечности, рассматриваются как конечный путь для общей реализации на конечность многих нервных влияний.
Благодаря конвергенции координация рефлекторной деятельности осуществляется с реализацией таких принципов, как облегчение и окклюзии.
Облегчение - превышение эффекта одновременного действия двух относительно слабых афферентных возбуждающих входов в ЦНС над суммой их раздельных эффектов. Объясняется это суммацией ВПСП с увеличением амплитуды в КРД в группе мотонейронов, в которых ВПСП достигают только значений подпороговых при разрешении активации входов.
Окклюзия - уменьшение эффекта одновременного возбуждения двух сильных афферентных входов, по сравнению с суммой эффектов при раздельном их раздражении.
Причина окклюзии заключается в том, что эти афферентные входы в результате конвергенции адресуются к общим "пороговых" мотонейронов и каждый может создать в них такое же надпороговая возбуждения, как и оба входа вместе.
Доминанта, по А. А. Ухтомским, - господствующий очаг возбуждения, который усиливается импульсами, поступающими из других центров, тормозя при этом остальные.
Доминанта формируется под влиянием адекватных стимулов извне или из внутренней среды, а также под влиянием гуморальных агентов, например, гормонов. Доминантными становятся те центры, которые обеспечивают удовлетворение жизненно необходимых потребностей. Доминантой может быть пищевая, половая, защитная потребность, определяет поведение в определенный момент. Для доминантного центра характерны следующие особенности: повышенная возбудимость, инертность возбуждения, пространственная суммация, устойчивость и продолжительность возбуждения, затормаживание других центров.
Доминанта в своем формировании проходит несколько стадий:
■ начальная - иррадиация (генерализация) возбуждения, которая характеризуется распространением возбуждения на многие другие центров;
■ концентрация возбуждения - происходит в центре биологически значимой в настоящее время реакции.
Отрицательная сторона доминанты - ее консервативность. Разрушить доминанту трудно: для этого необходимо создать новую, еще более сильную доминанту.
Пластичность - способность изменять реактивность под влиянием последовательных раздражений. Это свойство является необходимым условием для обеспечения нервных механизмов обучения.
Процессы обучения сопровождаются микроструктурными изменениями в нервных синапсах, особенно в области аксонного бугорка и на дендритах. Происходят и медиаторные сдвиги. Проявлением пластичности также способность нервных центров принимать на себя функцию утраченного или поврежденного центра. Именно благодаря этому свойству при повреждении или исключении частей мозга выполнения функций утраченных центров обеспечивают другие центры.
Структурной основой пластичности после исключения центра, во-первых, может быть восстановление нервных связей (на замену утраченным) благодаря формированию спраутинг - разветвления аксонных коллатералей, отходящие от противоположной стороны (от другой, неповрежденной полушария). Во-вторых, определенное значение имеет переобучение нейронов, в частности синапсов с явлениями "проторения" нервного пути по типу посттетанического потенциации путем изменений в пресинаптических мембранах и мобилизации большего количества медиатора, а также изменений аксотранспорту, следовательно и трофических процессов.
Кора полушарий - самый молодой и самый отдел ЦНС - обладает максимальной пластичностью и обеспечивает пластичность других мозговых центров. Нейрофизиологические эксперименты вооружают современную восстановительную неврологию и нейрохирургию новыми подходами к реабилитации (обучение языку после поражения моторного центра Брока, переобучение после протезирования).
СОВРЕМЕННЫЕ ПРЕДСТАВЛЕНИЯ ОБ ИНТЕГРАТИВНОЙ ДЕЯТЕЛЬНОСТИ МОЗГА
Интегративная деятельность ЦНС сводится к подчинению и объединения всех функциональных элементов организма в целостную систему, которая обладает определенной направленностью деятельности.
В осуществлении интегративной функции участвуют разные уровни ЦНС. Первый уровень интеграции осуществляется в результате взаимодействия ВПСП и ТПСП, генерируемые при активации синаптических входов нейрона. В том случае, когда возбуждающие и тормозные входы активируются одновременно, возникает суммирование синаптических потенциалов противоположной полярности и мембранный потенциал может отдаляться от того КРД, при котором в клетках низкопороговых зоны возникает ПД. Конвергенция возбуждающих и тормозных входов на мембране нейрона определяет частоту импульсов, генерируемых и выступает в качестве универсального фактора интегративной деятельности нервной клетки.
Второй уровень интеграции заключается в координационном деятельности нейронов элементарных нервных сетей и зависит от специфики морфологических отношений в ЦНС. Имеются в виду рассмотрены дивергенция, конвергенция, принцип общего конечного пути и принцип обратной связи.
Сложность задач, выполняемых нервной системой, приводит к высокой специализации нейронных объединений - нервных центров (третий уровень), деятельность которых связана с осуществлением различных функций. Функционирование ЦНС в целом осуществляется благодаря деятельности значительного количества таких центров, объединенных с помощью синаптических контактов и отличаются сложностью внутренних и внешних связей.
Нейрогуморальная регуляция функций обеспечивает адаптационные реакции от субклеточных до поведенческих. Можно выделить два вида механизмов, которые заложены в ее основе: жесткие (фиксированные) и гибкие (нефиксированные) (Бехтерева Η. П.).
В основе работы жестких механизмов регуляции лежат генетические программы, которые зафиксированы в процессе эволюции и осуществляют регуляцию достижения постоянно присутствующей цели. Например, регуляция функционирования дыхательного центра; чередование вдоха и выдоха обусловливается генотипическими механизмами; фенотипически же могут меняться продолжительность каждой фазы и амплитуда этих процессов в зависимости от потребности организма.
Гибкие механизмы обеспечивают выполнение организмом сегодняшних задач благодаря организации определенной функциональной системы и тут же прекращают функционирование. Осуществляются они срочно создаваемыми нейронными ансамблями. При этом главным принципом объединения нейронов является доминанта, которая обеспечивает синхронизацию работы нервных образований, входящих в ансамблей.
Программа нейрогуморальной регуляции функций реализуется осуществлением эфферентных влияний на исполнительные органы, работа которых обеспечивает адекватные изменения параметров, которые регулируются. Выделяют три вида таких воздействий: пусковые - вызывают или причиняют деятельность структуры, которую регулируют; адаптационные - влияют на величину реакции и соотношения отдельных ее компонентов; влияние готовности - формирует состояние готовности к ответам на пусковые и адаптационные воздействия.
Взаимодействие возбуждающих и тормозных нервных процессов обеспечивает быстрое и точное приспособление организма к условиям существования. Такое взаимодействие процессов возбуждения и торможения в ЦНС, обеспечивающих ее согласованную деятельность, называют координацией.
Выделяют семь основных принципов координации рефлекторной деятельности:
принцип общего конечного пути,
принцип реципрокного торможения,
принцип обратной связи,
Конвергенция - это морфологический принцип, который характеризуется схождением нескольких нейронов к меньшему их количеству.
Конвергенция, обеспечивающая схождение нервных импульсов к одному центральному нейрону, лежит в основе интегративной деятельности ЦНС. Она обусловлена схождением к телу и дендритам каждого нейрона аксонов множества других нервных клеток.
Схема организации конвергенции в ЦНС

1) вставочные нейроны, 2) эфферентный нейрон.
Конвергенция возбуждений лежит в основе облегчения рефлекторной реакции при одновременном раздражении нескольких афферентов.
Облегчение - это принцип координации рефлекторной деятельности, который характеризуется тем, что при одновременном раздражении двух афферентов возникает более сильная рефлекторная реакция, чем сумма рефлекторных ответов при раздельном возбуждении каждого из них.
Нервный центр состоит из двух зон: периферической и центральной. Центральная зона - это часть нервного центра, в которой из коллатералей афферента при передаче возбуждения выделяется достаточное количество медиатора для возникновения разрядной деятельности нейронов и рефлекторного ответа. При этом величина формирующегося ВПСП достаточна для того, чтобы ионные токи, циркулирующие между частично деполяризованной постсинаптической мембраной и аксонным холмиком, вызвали генерацию серии эфферентных ПД и рефлекторную реакцию.
Около 80% нейронов нервного центра локализуются в периферической зоне. Периферическая зона - это часть нервного центра, в которой из коллатералей афферента при передаче возбуждения выделяется недостаточное количество медиатора для возникновения разрядной деятельности нейронов и рефлекторного ответа. При этом величина ВПСП и циркулирующих местных ионных токов недостаточна для генерации в области аксонного холмика серии эфферентных ПД, а значит и рефлекторной реакции.
Облегчение рефлекторной реакции обусловлено перекрытием периферических зон расположенных рядом нервных центров. При одновременном раздражении двух афферентов, за счет конвергенции возбуждений в синапсах периферической зоны выделяется достаточное количество медиатора. Это обеспечивает возникновение разрядной деятельности нейронов, которые локализуются в периферической зоне, а значит усиление рефлекторного ответа.
Схема нейронной организации облегчения
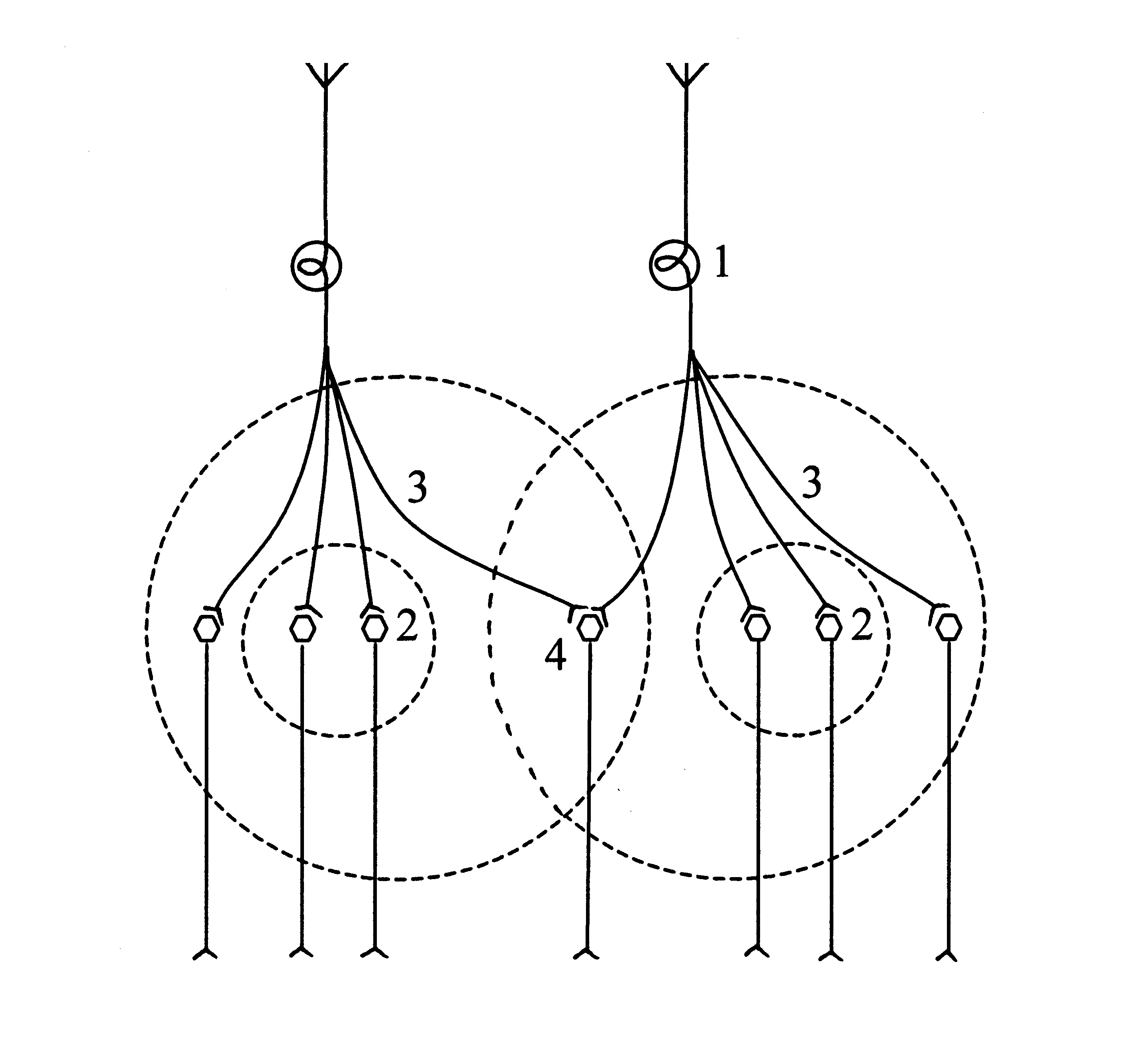
1) афференты, 2) центральные зоны нервных центров, 3) периферические зоны нервных центров, 4) область перекрытия периферических зон.

1) афференты, 2) вставочные нейроны, 3) мотонейрон.
Окклюзия - это принцип координации рефлексов, который характеризуется тем, что при одновременном раздражении двух афферентов сила рефлекторной реакции меньше, чем сумма рефлекторных ответов при раздельном возбуждении каждого афферента.
Окклюзия возникает вследствие перекрытия центральных зон нервных центров. При одновременном раздражении двух афферентов активируется меньше центральных нейронов, чем при раздельной их стимуляции, так как часть нервных клеток локализуется в области перекрытия центральных зон и являются общими для обоих нервных центров.
Схема нейронной организации окклюзии
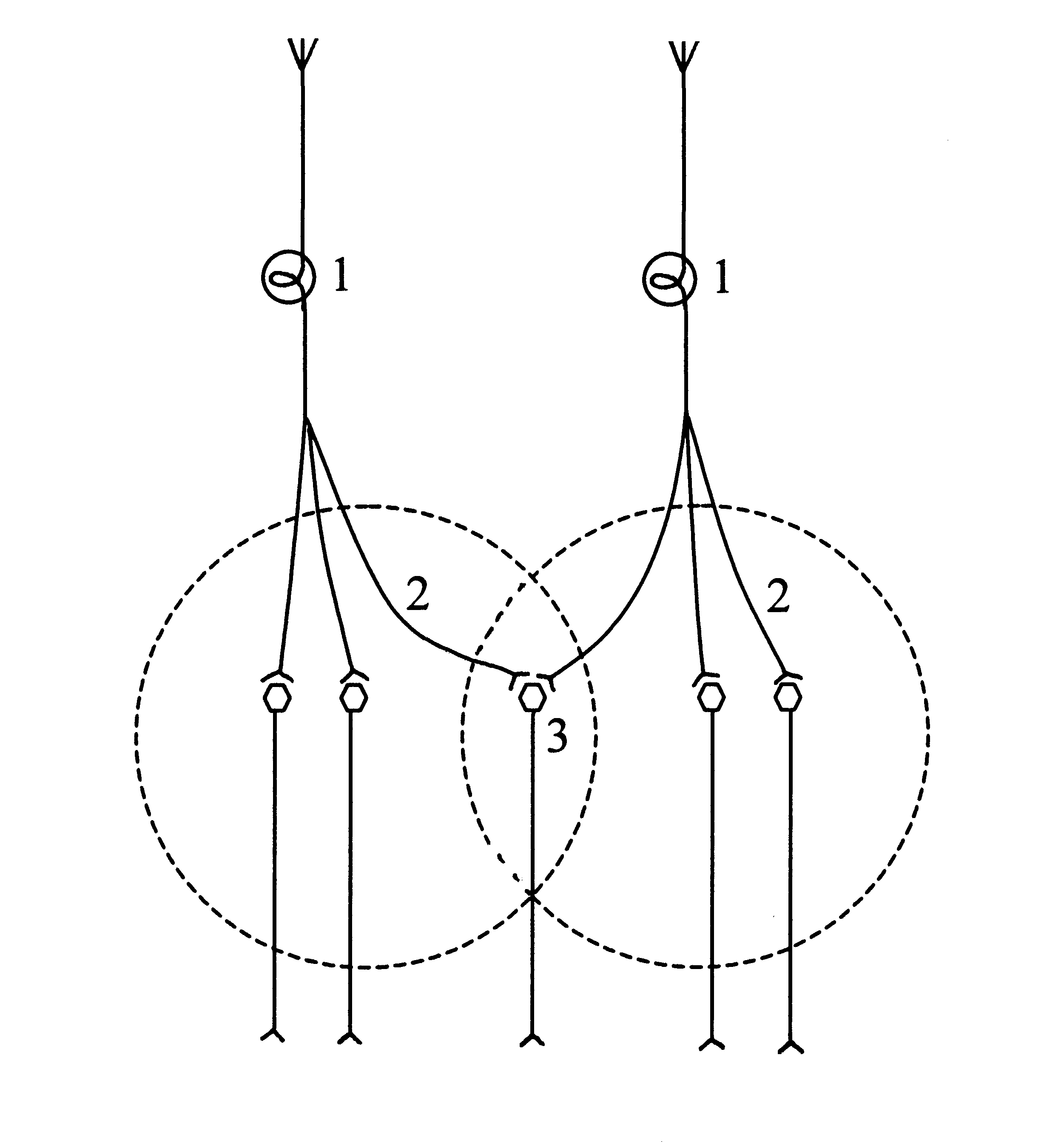
1) афференты, 2) центральные зоны нервных центров, 3) область перекрытия центральных зон.
Принцип реципрокного (сопряженного) торможения характеризуется одновременным угнетением одного нервного центра при возбуждении другого.
Принцип реципрокности лежит в основе противоположных по функциональному назначению ответных реакций. Примером реципрокного торможения является взаимодействие нервных центров, обеспечивающих поочередное сокращение и расслабление мышц-сгибателей и разгибателей во время ходьбы, бега, плавания и т.п. Возбуждение, которое идет к мотонейронам, иннервирующим мышцы-сгибатели, по коллатерали афферента поступает к тормозным клеткам Реншоу. Эти клетки тормозят мотонейроны мышц-разгибателей. Если бы одновременно возбуждались нервные центры мышц сгибателей и разгибателей, движение конечности стало бы невозможным.
Схема нейронной организации реципрокного торможения жевательных мышц
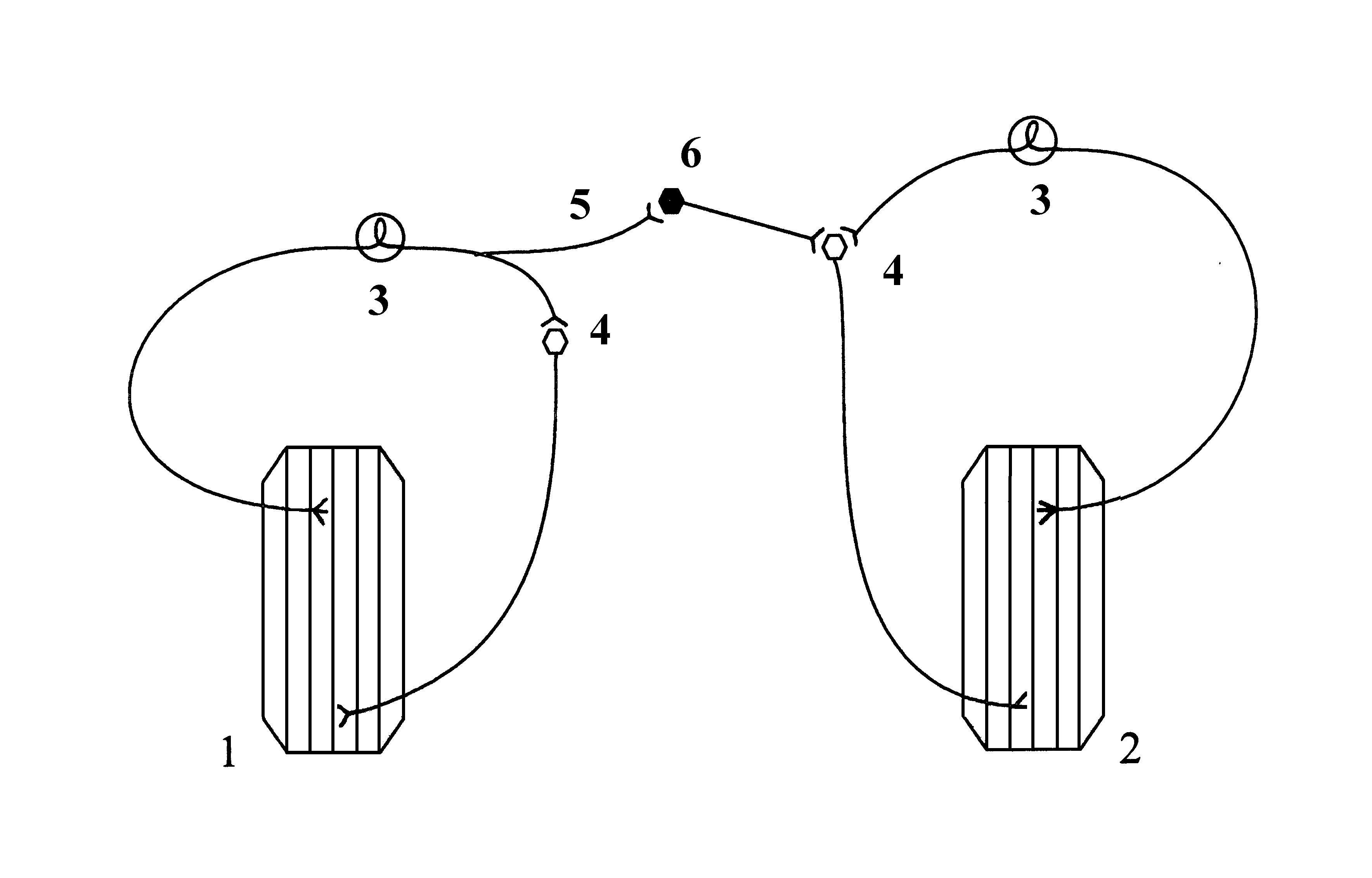
1) собственно жевательная мышца, 2) мышца, опускающая нижнюю челюсть, 3) афферентные нейроны, 4) мотонейроны, 5) коллатераль, 6) тормозной нейрон.
Другим примером использования реципрокного торможения может быть стискивание зубов при сильной боли, чтобы подавить защитные движения, или прикусывание нижней губы, чтобы не смеяться при щекотании.
Нервные клетки обладают способностью регулировать частоту проходящих через них сигналов, а значит и силу рефлекторного ответа за счет обратной связи. Обратная связь базируется на способности коллатералей аксонов нервных клеток устанавливать синаптические контакты со вставочными нейронами, роль которых сводится к обратному воздействию на нейроны, образующие эти аксонные коллатерали.
Схема положительной обратной связи

1) эфферентный нейрон, 2) коллатераль, 3) возбуждающий вставочный нейрон.
Схема отрицательной обратной связи
1) эфферентный нейрон, 2) коллатераль, 3) тормозной вставочный нейрон.
Для деятельности нервной системы, как единого целого, в естественных условиях характерно наличие преобладающих (доминантных) очагов возбуждения, которые, по А.А. Ухтомскому (1923), являются проявлением одной из основных закономерностей работы нервных центров.
Доминанта - это господствующий очаг возбуждения в ЦНС, подчиняющий себе нервные центры в интересах выполнения наиболее важной в данный момент времени поведенческой деятельности организма.
Доминанту характеризуют пять основных свойств:
способность к суммации,
способность реципрокно тормозить деятельность других нервных центров (тех, которые не связаны с организацией наиболее важной в данный момент поведенческой деятельности).
Доминанта усиливается за счет конвергенции афферентных возбуждений от различных рефлексогенных зон. Так, очаг доминантного возбуждения, который возникает при острой зубной боли, поддерживается посторонними раздражителями (прикосновение, яркий свет, резкий звук), которые усиливают страдания больного.
На способности доминантного очага угнетать другие нервные центры основан метод звуковой аналгезии (аудиоаналгезия). Звуковой рардражитель – звук или даже простой шум, создает в ЦНС доминанту, которая реципрокно тормозит очаги возбуждения, возникающие при различных стоматологических манипуляциях. При определенных условиях обезболивающий эффект аудиоаналгезии во время препарировании твердых тканей зуба может достигать 90%.
Принцип доминанты лежит в основе организации целенаправленной поведенческой деятельности, но не определяет ее адекватности биологическим и социальным потребностям человека. Примером этого могут служить алкогольная и наркотическая доминанты, которые, подчиняя себе большинство нервных центров с целью удовлетворения потребности в алкоголе или наркотическом веществе, разрушительно отражаются на физическом, психическом и социальном благополучии человека.
Рефлекторная реакция осуществляется целой группой рефлекторных дуг и нервных центров. Координация рефлекторной деятельности обеспечивает согласованную деятельность органов и систем организма. Она осуществляется с помощью следующих процессов:
Временное и пространственное облегчение. Это усиление рефлекторной реакции при действии ряда последовательных раздражителей или одновременном их воздействии на несколько рецептивных полей. Объясняется явлением суммации в нервных центрах.
Окклюзияявление противоположное облегчению. Когда рефлекторная реакция на два или более сверхпороговых раздражителя меньше, чем ответы на их раздельное воздействие. Оно связано с конвергенцией нескольких возбуждающих импульсов на одном нейроне.
Принцип общего конечного пути. Разработан Ч. Шеррингтоном. В основе его лежит явление конвергенции. Согласно этому принципу на одном эфферентном мотонейроне могут образовывать синапсы нескольких афферентных, входящих в несколько рефлекторных дуг. Этот нейрон называется общим конечным путем и участвует в нескольких рефлекторных реакциях. Если взаимодействие этих рефлексов приводит к усилению общей рефлекторной реакции, такие рефлексы называются союзными. Если же между афферентными сигналами происходит борьба за мотонейрон - конечный путь, то антагонистическими.
Реципрокное торможение. Обнаружено Ч. Шеррингтоном. Это явление торможения одного центра в результате возбуждения другого. Т.е. в этом случае тормозится антагонистический центр. В реципрокных взаимоотношениях находятся центры вдоха и выдоха продолговатого мозга, центры сна и бодрствования и т.д.
Принцип доминанты. Открыт А.А. Ухтомским. Доминанта - это преобладающий очаг возбуждения в ЦНС, подчиняющий себе другие НЦ. Доминантный центр обеспечивает комплекс рефлексов, которые необходимы в данный момент для достижения определенной цели. При некоторых условиях возникают питьевая, пищевая, оборонительная, половая и др. доминанты. Свойствами доминантного очага являются повышенная возбудимость, стойкость возбуждения, высокая способность к суммации, инертность.
Принцип обратной афферентации. Результаты рефлекторного акта воспринимаются нейронами обратной афферентации и информация от них поступает обратно в нервный центр. Там они сравниваются с параметрами возбуждения и рефлекторная реакция корректируется.
24. Роль спинного мозга в процессах регуляции соматических и вегетативных функций организма. Клинически важные спинальные рефлексы. Проводниковая функция спинного мозга.Спинной мозг находится в позвоночном канале и выполняет рефлекторную и проводниковую функции. Первая обеспечивается его нервными центрами, вторая проводящими путями. Он имеет сегментарное строение. Причем деление на сегменты является функциональным. Каждый сегмент образует передние и задние корешки. Задние являются чувствительными, т.е. афферентными, передние двигательными, эфферентными. Морфологически тела нейронов спинного мозга образуют его серое вещество. Функционально все его нейроны делятся на мотонейроны, вставочные, нейроны симпатического и парасимпатического отделов вегетативной нервной системы. Мотонейроны, в зависимости от функционального значения, разделяются на альфа- и гамма-мотонейроны. К α-мотонейронам идут волокна афферентных путей, которые начинаются от интрафузальных, т.е. рецепторных мышечных клеток. Тела α-мотонейронов расположены в передних рогах спинного мозга, а их аксоны иннервируют скелетные мышцы. Гамма-мотонейроны регулируют напряжение мышечных веретен т.е. интрафузальных волокон. Таким образом они участвуют в регуляции сокращений скелетных мышц. Поэтому при перерезке передних корешков мышечный тонус исчезает. Интернейроны обеспечивают связь между центрами спинного мозга и вышележащих отделов ЦНС.
Все рефлексы спинного мозга делятся на соматические, т.е. двигательные и вегетативные. Соматические рефлексы делятся на сухожильные и кожные. Сухожильные рефлексы возникают при механическом раздражении мышц и сухожилий. Их небольшое растяжение приводит к возбуждению сухожильных рецепторов и α-мотонейронов спинного мозга. В результате возникает сокращение мышц, в первую очередь разгибателей. Кожные рефлексы обусловлены раздражением рецепторов кожи, но проявляются двигательными реакциями. Ими являются подошвенный и брюшной (объяснение). Спинальные нервные центры находятся под контролем вышележащих. Поэтому после перерезки между продолговатым и спинным мозгом возникает спинальный шок и тонус всех мышц значительно уменьшится. Вегетативные рефлексы спинного мозга делятся на симпатические и парасимпатические. Те и другие проявляются реакцией внутренних органов на раздражение рецепторов кожи, внутренних органов, мышц. Вегетативные нейроны спинного мозга образуют низшие центры регуляции тонуса сосудов, сердечной деятельности, просвета бронхов, потоотделения, мочевыведения, дефекации, эрекции, эйякуляции и т.д.Проводниковая функция состоит в обеспечении связи периферических рецепторов, центров спинного мозга с вышележащими отделами ЦНС, а также его нервных центров между собой. Она осуществляется проводящими путями. Все пути спинного мозга делятся на собственные или проприоспинальные, восходящие и нисходящие. Проприоспинальные пути связывают между собой нервные центры разных сегментов спинного мозга. Их функция заключается в координации тонуса мышц, движений различных метамеров туловища. К восходящим путям относятся:Пучки Голля и Бурдаха проводят нервные импульсы от проприорецепторов мышц и сухожилий к соответствующим ядрам продолговатого мозга, а затем таламусу и соматосенсорным зонам коры. Благодаря этим путям производится оценка и коррекция позы туловища. Пучки Говерса и Флексига передают возбуждение от проприорецепторов, механорецепторов кожи к мозжечку. За счет этого обеспечивается восприятие и бессознательная координация позы. Спиноталамические тракты проводят сигналы от болевых, температурных, тактильных рецепторов кожи к таламусу, а затем соматосенсорные зоны коры. Они обеспечивают восприятие соответствующих сигналов и формирование чувствительности. Нисходящие пути также образованы несколькими трактами. ортикоспинальные пути идут от пирамидных и экстрапирамидных нейронов коры к α-мотонейронам спинного мозга. За счет них осуществляется регуляция произвольных движений. Руброспинальный путь проводит сигналы от красного ядра среднего мозга к α-мотонейронам мышц сгибателей. Вестибулоспинальный путь передает сигналы от вестибулярных ядер продолговатого мозга, в первую очередь ядра Дейтерса, к α-мотонейронам мышц разгибателей. За счет этих двух путей регулируется тонус соответствующих мышц при изменениях положения тела.
25. Продолговатый мозг и мост, участие их центров в процессах саморегуляции функций.Основными функциями продолговатого мозга являются проводниковая, рефлекторная и ассоциативная. Первая осуществляется проводящими путями, проходящими через него. Вторая, нервными центрами. В ромбовидной ямке продолговатого мозга находятся ядра 10, 11, 12 пар черепно-мозговых нервов, а также ретикулярная формация. Рефлекторные функции делятся на соматические и вегетативные. Соматическими являются статические рефлексы продолговатого мозга, относящиеся к позно-тоническим или рефлексам позы. Эти рефлексы осуществляются ядром Дейтерса из группы вестибулярных ядер. От него к мотонейронам разгибателей спинного мозга идут нисходящие вестибулоспинальныне тракты. Рефлексы возникают тогда, когда возбуждаются вестибулярные рецепторы или проприорецепторы мышц шеи. Коррекция положения тела происходит за счет изменения тонуса мышц. Например, при запрокидывании головы животного назад повышается тонус разгибателей передних конечностей и снижается тонус разгибателей задних. При наклоне головы вперед возникает обратная реакция. При повороте головы в сторону, повышается тонус разгибателей конечности на этой стороне и сгибателей противоположной конечности. В продолговатом мозге находятся жизненно важные центры. К ним относятся дыхательный, сосудодвигательный центры и центр регуляции сердечной деятельности. Первый обеспечивает смену фаз дыхания, второй - тонус периферических сосудов, третий регуляцию частоты и силы сердечных сокращений. В области ядер блуждающего нерва находятся центры слюноотделения, секреции желудочных, кишечных желез, поджелудочной железы и печени. Здесь же расположены центры регуляции моторики пищеварительного канала. Важной функцией продолговатого мозга является формирование защитных рефлексов. В нем находятся рвотный центр, центры кашля, чихания, смыкания век и слезотечения при раздражении роговицы. Здесь расположены бульбарные отделы центров, участвующих в организации пищевых рефлексов - сосания, жевания, глотания. В продолговатом мозге происходит первичный анализ ряда сенсорных сигналов. В частности, в нем расположены ядра слухового нерва, верхнее вестибулярное ядро, а к ядрам языкоглоточного нерва поступают сигналы от вкусовых рецепторов. От рецепторов кожи лица они идут к ядрам тройничного нерва.
Мост имеет тесные функциональные связи со средним мозгом. Эти отделы ствола мозга также осуществляют проводниковую и рефлекторную функции. Проводниковая обеспечивается восходящими и нисходящими путями, идущими через них. Рефлекторная их нервными центрами. Нейроны моста формируют его ретикулярную формацию, ядра лицевого, отводящего нервов, двигательную часть ядер тройничного нерва и его чувствительное ядро.
Читайте также: