
Рефлекторные дуги вегетативного отдела нервной системы рисунок
СОМАТИЧЕСКАЯ НЕРВНАЯ СИСТЕМА
иннервирует скелетную мускулатуру
центры находятся в передних рогах спинного мозга
рефлекторная дуга состоит как минимум из 2 нейронов:
I нейрон - чувствительный, его перикарион лежит в спинномозговом ганглии, длинный дендрит отходит на периферию, где заканчивается рецептором, аксон входит в задние рога спинного мозга, проходит в передний рог (или переключается на ассоциативный нейрон) и образует синапс со II нейроном; I нейрон - пурин-пептидергический, нейромедиаторы - АТФ, субстанция Р, кальцитонин ген-родственный пептид
II нейрон - двигательный или эфферентный, его перикарион лежит в передних рогах спинного мозга, аксон через передние рога выходит из спинного мозга и идет к скелетной мышце, где образуется аксо- мышечный синапс; II нейрон - холинергический, нейромедиатор - ацетилхолин, на постсинаптической мембране (т.е. на мембране мышечного волокна) имеются Н-холинорецепторы скелетных мышц
ВЕГЕТАТИВНАЯ НЕРВНАЯ СИСТЕМА
иннервирует все внутренние органы, сердце и сосуды, экзокринные и эндокринные железы, органы чувств
подразделяется на 2 отдела - симпатический и парасимпатический
каждый орган, как правило, получает и симпатическую, и парасимпатичеcкую иннервацию
СИМПАТИЧЕСКАЯ НЕРВНАЯ СИСТЕМА
центры находятся в боковых рогах грудного и поясничного отделов спинного мозга
рефлекторная дуга состоит как минимум из 3 нейронов:
I нейрон - чувствительный, его перикарион лежит в спинномозговом ганглии, длинный дендрит отходит на периферию, где заканчивается рецептором, аксон входит в задние рога спинного мозга, проходит в боковой рог (или переключается на ассоциативный нейрон) и образует синапс со II нейроном; I нейрон - пурин-пептидергический, нейромедиаторы - АТФ, субстанция Р, кальцитонин ген-родственный пептид
II нейрон - называется преганглионарным; эфферентный, его перикарион и дендриты лежат в боковых рогах спинного мозга, аксон через передние рога выходит из спинного мозга и идет к симпатическому ганглию,где образует синапсы с III нейроном;II нейрон- холинергический,нейромедиатор - ацетилхолин
III нейрон - называется постганглионарным; эфферентный, его перикарион и дендриты лежат в симпатических ганглиях (пре- и паравертебральные ганглии, ); на перикарионе и дендритах III нейрона имеются Н-холинорецепторы, через которые происходит синаптическая передача между II (преганглионарным) и III(постганглионарным) нейронами (ацетилхолин высвобождается из пресинаптической части, принадлежащей I нейрону и взаимодействует с Н-холинорецепторами, находящимися на постсинаптической мембране, то есть на мембране III нейрона); аксон выходит из ганглия и идет к иннервируемому органу, где образуются синаптические соединения; III нейрон - адренергический, нейромедиатор - норадреналин; синаптическая передача между III (постганглионарным) нейроном и рабочим органом осуществляется с помощью норадреналина, который высвобождается из пресинаптической части (принадлежащей III-нейрону) и взаимодействует с адренорецепторами, находящимися на постсинаптической мембране синапса, а постсинаптическая мембрана - это уже мембрана не нейрона, а органа; в различных органах имеются разные комбинации разных подтипов a-и b-адренорецепторов
ПАРАСИМПАТИЧЕСКАЯ НЕРВНАЯ СИСТЕМА
центры находятся в боковых рогах крестцового отдела спинного мозга, продолговатом мозге и мосте (ядра III, VII, IX, X, черепномозговых нервов)
рефлекторная дуга состоит как минимум из 3 нейронов:
I нейрон - чувствительный, его перикарион лежит в спинномозговом ганглии или в толще нервного ла или в специальных чувствительных ганглиях парасимпатичесокй нервной системы (ganglion geniculi, ganglia superius et inferius, g.nodosum), или непосредственно в стволе мозга (nucleus tractus solitarii - n. vagus; nucleus sensorius principallis nervi trigemini), длинный дендрит отходит на периферию, где заканчивается рецептором, аксон входит в мозг (мост, продолговатый мозг), или в боковые рога спинного мозга (или переключается на ассоциативный нейрон) и образует синапс со II нейроном; I нейрон - пyрино-пептидергический, нейромедиаторы - АТФ, субстанция Р, кальцитонин ген-родственный пептид
II нейрон - называется преганглионарным; эфферентный, его перикарион и дендриты лежат в боковых рогах крестцового отдела спинного мозга или продолговатом мозге, мосте: ядро Edinger- Westphal - III пара черепномозговых нервов; nuclei salivatory - VII и IX черепномозговые нервы; дорсальное ядро n. vagus; nucleus ambiguus - n.vagus) аксон выходит из спинного мозга или в составе черепно-мозговых нервов идет к парасимпатическому ганглию, где образует синапсы с III нейроном; II нейрон - холинергический, нейромедиатор - ацетилхолин
III нейрон - называется постганглионарным; эфферентный, его перикарион и дендриты лежат в парасимпатических ганглиях (вегетативные ганглии черепно-мозговых нервов /g. ciliary, g.oticum, g pterygopalatinum, g. submandibulare/ или внутриорганные ганглии (интрамуральные ганглии)); на перикарионе и дендритах III нейрона имеются Н-холинорецепторы, через которые происходит синаптическая передача между II (преганглионарным) и III (постганглионарным) нейронами (ацетилхолин высвобождается из пресинаптической части, принадлежащей II нейрону и взаимодействует с Н-холинорецепторами, находящимися на постсинаптической мембране, то есть на мембране III нейрона; аксон выходит из ганглия и идет к иннервируемому органу или уже находится в органе, где образуются синаптические соединения; III нейрон - холинергический, нейромедиатор - ацетилхолин; синаптическая передача между III (постганглионарным) нейроном и рабочим органом осуществляется с помощью ацетилхолина, который высвобождается из пресинаптической части (III-нейрон) и взаимодействует с М-холинорецепторами, находящимися на постсинаптической мембране синапса, а постсинаптическая мембрана - это уже мембрана не нейрона, а органа

Рефлекторные дуги соматической (А), симпатического (Б) и парасимпатического (В) отделов вегетативной нервной системы
1 — чувствительный нейрон
А: 2 — двигательный нейрон
Б, В: 2 — преганглионарный нейрон
3 — постганглионарный нейрон
D1 — постганглионарный нейрон (или клетка Догеля I типа)
D2 — клетка Догеля II типа
D3 — клетка Догеля III типа (клетки Догеля — см. раздел Ганглии)
Вопрос 20. Структурные единицы нервной системы
Функии и структура нервной системы.
Рефлекторная дуга.
Нервы.
Нервные узлы.
1. Функции нервной системы — она:
• осуществляет объединение частей организма в единое целое (интеграцию); V обеспечивает регуляцию разнообразных процессов, координацию
функции различных органов и тканей и взаимодействие организма с внешней средой;
• воспринимает многообразную информацию, поступающую из внешней среды и из внутренних органов, перерабатывает ее и генерирует сигналы, обеспечивающие ответные реакции, адекватные действующим раздражителям.
Анатомически нервную систему подразделяют:
• на ЦНС — головной и спинной мозг;
V ПНС — периферические нервные узлы (ганглии), нервы и нервные окончания.
Физиологически, в зависимости от характера иннервации органов и тканей:
• на соматическую (анимальную) нервную систему, которая регулирует преимущественно функции произвольного движения;
• автономную (вегетативную) нервную систему, которая регулирует деятельность внутренних органов и желез. Влияя на активность обмена веществ в различных органах и тканях в соответствии с меняющимися условиями их функционирования и внешней среды, она осуществляет адаптационно-трофическую функцию.
Вегетативная нервная система подразделяется на взаимодействующие друг с другом отделы, которые различаются локализацией центров в мозге и периферических узлах, а также характером влияния на внутренние органы:
В соматическую и автономную нервную систему входят звенья, расположенные в ЦНС и ПНС.
Функционально ведущей тканью органов нервной системы является нервная ткань, включающая нейроны и глию .
Скопления нейронов в ЦНС обычно называются ядрами, а в ПНС — узлами (ганглиями).
Пучки нервных волокон в ЦНС носят название трактов, в ПНС они образуют нервы.
Нервные центры — скопления нервных клеток в ЦНС и ПНС, в которых между ними осуществляется синаптическая передача. Они обладают сложной структурой, богатством и разнообразием внутренних и внешних связей и специализированы на выполнении определенных функций.
По характеру морфофункциональной организации различают нервные центры:
• ядерного типа, в которых нейроны располагаются без видимой упорядоченности (вегетативные ганглии, ядра спинного и головного мозга);
• экранного типа, в которых нейроны, выполняющие однотипные функции, собраны в виде отдельных слоев, сходных с экранами, на которых проецируются нервные импульсы (кора мозжечка, кора полушарий большого мозга, сетчатка глаза). Внутри слоев и между ними имеются многочисленные ассоциативные связи.
В нервных центрах происходят процессы конвергенции и дивергенции нервного возбуждения, функционируют механизмы обратной связи.
Конвергенция — схождение различных путей проведения нервных импульсов к меньшему числу нервных клеток. На нейронах могут иметься окончания клеток разных типов, что обеспечивает конвергенцию влияний из различных источников.
Дивергенция — образование связей одного нейрона с большим числом других, на деятельность которых он оказывает влияние, обеспечивая перераспределение импульсов с иррадиацией возбуждения.
Механизмы обратной связи дают возможность нейронам самим регулировать величину поступающих к ним сигналов благодаря связям их аксонных коллатералей со вставочными клетками. Последние оказывают влияние (обычно тормозное) как на нейроны, так и на терминали конвергирующих к ним волокон.
2. Рефлекторная дуга является функциональной единицей нервной системы и представляет собой цепочки нейронов, которые обеспечивают реакции рабочих органов (органов-мишеней) в ответ на раздражение рецепторов. В рефлекторных дугах нейроны, связанные друг с другом синапсами, образуют 3 звена
- рецепторное (афферентное);
- эффекторное;
- ассоциативное (вставочное) расположенное между ними; в простейшем варианте дуги может отсутствовать.
На различные звенья дуги оказывают регуляторные воздействия связанные с ними нейроны вышележащих центров, вследствие чего рефлекторные дуги имеют сложное строение. Рефлекторные дуги в соматическом (анимальном) и автономном (вегетативном) отделах нервной системы обладают рядом особенностей.
Соматическая (анималъная) рефлекторная дуга:
рецепторное звено образовано афферентными псевдоуниполярными нейронами, телакоторых располагаются в спиналъных ганглиях. Дендриты этих клеток образуют чувствительные нервные окончания в коже или скелетной мускулатуре, а аксоны вступают в спинной мозг в составе задних корешков и направляются в задние рога его серого вещества, образуя синапсы на телах и дендритах вставочных нейронов. Некоторые веточки (коллатерали) аксонов псевдоуниполярных нейронов проходят (не образуя связей в задних рогах) непосредственно в передние рога, где оканчиваются на мотонейронах (формируя с ними двухнейронные рефлекторные дуги);
ассоциативное звено (которого может не быть)представлено мулътиполярными вставочными нейронами, дендриты и тела которых расположены в задних рогах спинного мозга, а аксоны направляются в передние рога, передавая импульсы на телаи дендриты эффекторных нейронов;
эффекторное звено образовано мулътиполярными мотонейронами, тела и дендриты которых лежат в передних рогах, а аксоны выходят из спинного мозга в составе передних корешков, направляются к спиналъному ганглию и далее в составе смешанного нерва - к скелетной мышце, на волокнах которой их веточки образуют нервно-мышечные синапсы (моторные, или двигательные, бляшки).
Автономная (вегетативная) рефлекторная дуга
рецепторное звено, как и в соматической рефлекторной дуге, образовано афферентными псевдоуниполярными нейронами, телакоторых располагаются в спинальных ганглиях, однако дендриты этих клеток образуют чувствительные нервные окончания в тканях внутренних органов, сосудов и желез. Их аксонывступают в спинной мозг в составе задних корешков и, минуя задние рога, направляются в боковые рогасерого вещества, образуя синапсы на телах и дендритах вставочных нейронов;
ассоциативное звено представлено мулътиполярными вставочными нейронами, дендриты и тела которых расположены в боковых рогах спинного мозга, а аксоны (преганглионарные волокна) покидают спинной мозг в составе передних корешков, направляясь в один из вегетативных ганглиев, где и оканчиваются на дендритах и телах эффекторных нейронов;
эффекторное звено образовано мультиполярными нейронами, тела которых лежат в составе вегетативных ганглиев, а аксоны (постганглионарные волокна) в составе нервных стволов и их ветвей направляются к клеткам рабочих органов — гладких мышц, желез, сердца.
3. Нервы (нервные стволы) связывают нервные центры головного и спинного мозга с рецепторами и рабочими органами. Они образованы пучками нервных волокон, которые объединены соединительнотканными компонентами (оболочками):
Большинство нервов являются смешанными, т. е. включают афферентные и эфферентные нервные волокна. Пучки нервных волокон содержат до нескольких тысяч миелиновых и безмиелиновых волокон, соотношение между которыми в различных нервах неодинаково; по функциональному признаку они относятся к анимальной и вегетативной нервной системе.
Эндоневрий — тонкие прослойки рыхлой волокнистой соединительной ткани, окружающие отдельные нервные волокна и связывающие их в единый пучок. Содержит немногочисленные клетки и волокна (преимущественно ретикулярные), мелкие кровеносные сосуды.
Периневрий — оболочка, покрывающая каждый пучок нервных волокон снаружи и отдающая перегородки вглубь пучка. Периневрий образован 2—10 концентрическими пластами уплощенных клеток, связанных плотными и щелевыми соединениями. Цитоплазма клеток, в особенности наружных слоев, содержит многочисленные пиноцитозные пузырьки. Между слоями клеток в щелевидных пространствах, заполненных жидкостью, располагаются компоненты базальной мембраны и продольно ориентированные коллагеновые волокна. В терминальной части нерва периневрий образован лишь 1 слоем плоских клеток, который дистально резко обрывается и имеет вид открытой манжетки.
Эпиневрий — наружная оболочка нерва, связывающая воедино пучки нервных волокон (число которых зависит от диаметра нерва и варьирует от одного до нескольких десятков). Состоит из плотной волокнистой соединительной ткани, содержащей жировые клетки, кровеносные и лимфатические сосуды.
- Нервные узлы (ганглии)- скопление нейронов вне ЦНС.
Типы нервных узлов:
Чувствительные (сенсорные) нервные узлы содержат псевдоуниполярные или биполярные (в спиральном и вестибулярном ганглиях) афферентные нейроны и располагаются по ходу задних корешков спинного мозга (спинномозговые, или спинальные, узлы) и черепномозговых нервов (5, 7, 8, 9, 10-й пар).
Спинномозговой (спинальный) узел (ганглий) имеет веретеновидную форму и покрыт капсулой из плотной волокнистой соединительной ткани. По его периферии находятся плотные скопления тел псевдоуниполярных нейронов, а центральная часть занята их отростками и расположенными между ними тонкими прослойками эндоневрия, несущими сосуды.
Псевдоуниполярные нейроны характеризуются сферическим телом и светлым ядром с хорошо заметным ядрышком. Выделяют крупные и мелкие клетки, которые, вероятно, различаются видами проводимых импульсов. Цитоплазма нейронов содержит многочисленные митохондрии, цистерны гранулярной эндоплазматической сети, элементы комплекса Голъджи, лизосомы. Каждый нейрон окружен слоем прилежащих к нему уплощенных клеток олигодендроглии (мантийными глиоцитами, или клетками-сателлитами) с мелкими округлыми ядрами; снаружи от глиальной оболочки имеется тонкая соединительнотканная оболочка. От тела псевдоуниполярного нейрона отходит отросток, разделяющийся Т-образно на афферентную (дендритную) и эфферентную (аксональную) ветви, которые покрываются миелиновыми оболочками. Афферентная ветвь заканчивается на периферии рецепторами, эфферентная в составе заднего корешка вступает в спинной мозг. Так как переключения нервного импульса с одного нейрона на другой в пределах спинномозговых узлов не происходит, они не являются нервными центрами. Нейроны спинномозговых узлов содержат такие нейромедиаторы, как ацетилхолин, глутаминовая кислота, вещество Р, соматостатин, холецистокинин, гастрин, вазоинтестиналъный пептид.
Автономные (вегетативные) нервные узлы (ганглии) могут располагаться вдоль позвоночника (паравертебральные ганглии), или впереди него (превертебралъные ганглии), а также в стенке органов — сердца, бронхов, пищеварительного тракта, мочевого пузыря и др. (интрамуралъные ганглии) или вблизи их поверхности. Иногда они имеют вид мелкий Хот нескольких клеток до нескольких десятков клеток) скоплений нейронов, расположенных по ходу некоторых нервов или лежащих интрамурально (микроганглии). К вегетативным узлам подходят преганглионарные волокна (миелиновые), содержащие отростки клеток, тела которых лежат в ЦНС. Эти волокна сильно ветвятся и образуют многочисленные синаптические окончания на клетках вегетативных узлов. Благодаря этому осуществляется конвергенция большого числа терминалей преганглионарных волокон на каждый нейрон ганглия. В связи с наличием синаптической передачи вегетативные узлы относят к нервным центрам ядерного типа.
Вегетативные нервные узлы по функциональному признаку и локализации разделяются:
Симпатические нервные узлы (пара- и превертебральные) получают преганглионарные волокна от клеток, расположенных в вегетативных ядрах грудных и поясничных сегментов спинного мозга. Нейромедиатором преганглионарных волокон является ацетилхолин, а постганглионарных — норадреналин (за исключением потовых желез и некоторых кровеносных сосудов, имеющих холинергическую симпатическую иннервацию). Помимо этих нейромедиаторов, в узлах выявляются энкефалины, вещество Р, соматостатин, холецистокинин.
Парасимпатические нервные узлы (интрамуральные, лежащие вблизи органов, или узлы головы) получают преганглионарные волокна от клеток, расположенных в вегетативных ядрах продолговатого и среднего мозга, а также крестцового отдела спинного мозга. Эти волокна покидают ЦНС в составе 3, 7, 9, 10-й пар черепномозговых нервов и передних корешков крестцовых сегментов спинного мозга. Нейромедиатором пре- и постганглионарных волокон является ацетилхолин. Кроме него роль медиаторов в этих ганглиях играют серотонин, АТФ, возможно, некоторые пептиды.
Большинство внутренних органов имеют двойную вегетативную иннервацию, т. е. получают постганглионарные волокна от клеток, расположенных как в симпатических, так и в парасимпатических узлах. Реакции, опосредуемые клетками симпатических и парасимпатических узлов, часто имеют противоположную направленность, например: симпатическая стимуляция усиливает, а парасимпатическая - тормозит сердечную деятельность.
Общий план строения симпатических и парасимпатических нервных узлов сходен. Вегетативный узел покрыт соединительнотканной капсулой и содержит диффузно или группами расположенные тела мультиполярных нейронов, их отростки в виде безмиелиновых или, реже, миелиновых волокон и эндоневрий. Тела нейронов имеют неправильную форму, содержат эксцентрично расположенное ядро, окружены (обычно не полностью) оболочками из глиальных клеток-сателлитов (мантийных глиоцитов). Часто встречаются многоядерные и полиплоидные нейроны.
Интрамуральные узлы и связанные с ними проводящие пути ввиду их высокой автономии, сложности организации и особенностей медиаторного обмена некоторыми авторами выделяются в самостоятельный метасимпатический отдел ВНС. В частности, общее число нейронов в интрамуральных узлах кишки выше, чем в спинном мозге, а по сложности их взаимодействия в регуляции перистальтики и секреции их сравнивают с миникомпьютером.
В интрамуральных узлах описаны нейроны 3 типов:
• длинноаксонные эфферентные нейроны (клетки Догеля I типа) численно преобладают. Это крупные или средних размеров эфферентные нейроны с короткими дендритами и длинным аксоном, направляющимся за пределы к рабочему органу, на клетках которого он образует двигательные или секреторные окончания;
• равноотростчатые афферентные нейроны (клетки Догеля II типа) содержат длинные дендриты и аксон, уходящий за пределы данного ганглия в соседние и образующий синапсы на клетках I и III типов. Эти клетки, по-видимому, входят в качестве рецепторного звена в состав местных рефлекторных дуг, которые замыкаются без захода нервного импульса в ЦНС. Наличие таких дуг подтверждается сохранением функционально активных афферентных, ассоциативных и эфферентных нейронов в трансплантированных органах (например, сердце);
• ассоциативные клетки (клетки Догеля III типа) — местные вставочные нейроны, соединяющие своими отростками несколько клеток I и II типа, морфологически сходные с клетками Догеля II типа. Дендриты этих клеток не выходят за пределы узла, а аксоны направляются в другие узлы, образуя синапсы на клетках I типа.
Организация спинного мозга
\. Спинной мозг располагается в позвоночном канале и имеет вид округлого тяжа, расширенного в шейном и поясничном отделах и пронизанного центральным каналом.
Состоит из 2 симметричных половин, разделенных спереди срединной щелью, сзади — срединной бороздой, и характеризуется сегментарным строением; с каждым сегментом связана пара передних (вентральных) и пара задних (дорсальных) корешков.
В спинном мозге различают:
· серое вещество, расположенное в его центральной части;
· белое вещество, лежащее по периферии.
Серое вещество на поперечном разрезе имеет вид бабочки и включает:
• парные передние (вентральные);
• задние (дорсальные);
• боковые (латеральные) рога (в действительности представляют собой непрерывные столбы, идущие вдоль спинного мозга).
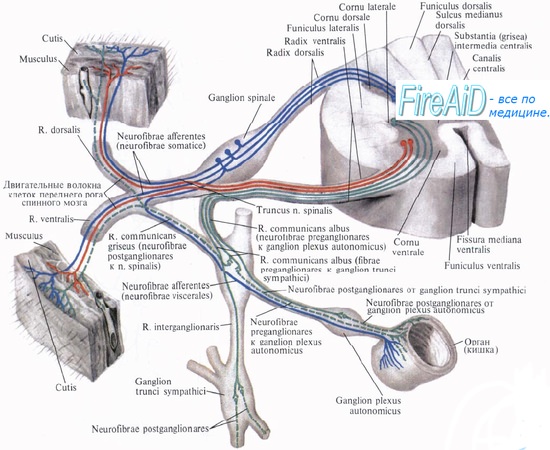
Рефлекторная дуга вегетативной нервной системы.
Клеточное тело воспринимающего нейрона как для анимальной, так и для вегетативной нервной системы помещается в спинномозговом узле, ganglion spinale, куда стекаются афферентные пути как от органов животной жизни, так и от органов растительной жизни и который, таким образом, является смешанным анимально-вегетативным узлом.
Клеточное тело вставочного нейрона вегетативной нервной системы в отличие от анимальной нервной системы помещается в боковых рогах спинного мозга. При этом аксон вставочного анимального нейрона, исходящий из клеток заднего рога, заканчивается в пределах спинного мозга среди клеток его передних рогов. Что же касается вставочного нейрона вегетативной нервной системы, то он в спинном мозге не заканчивается, а выходит за его пределы, к нервным узлам, расположенным на периферии.
Выйдя из спинного мозга, аксон вставочного нейрона подходит или к узлам симпатического ствола, ganglia trunci sympathici, относящимся к симпатическому отделу вегетативной нервной системы (они образуют симпатический ствол), или волокна не заканчиваются в этих узлах, а направляются к предпозвоночным узлам, расположенным более периферично, между симпатическим стволом и органом (ganglia coeliaca, ganglia mesenterica).
Кроме макроскопически видимых обособленных узлов, по ходу вегетативных нервов встречаются мигрировавшие сюда в ходе эмбрионального развития небольшие группы эффекторных нейронов — микроганглии. Все волокна, идущие до узлов первого, второго или третьего порядка и являющиеся аксонами промежуточного нейрона, называются предузловыми волокнами, rami preganglionares. Они покрыты миелином.

Третий, эффекторный, нейрон анимальной рефлекторной дуги помещается в передних рогах спинного мозга, а эффекторный нейрон вегетативной рефлекторной дуги вынесен в процессе развития из центральной нервной системы в периферическую, ближе к рабочему органу, и располагается в вегетативных нервных узлах. Из такого расположения эффекторных нейронов на периферии вытекает главный признак вегетативной нервной системы — двухнейронность эфферентного периферического пути: первый нейрон — вставочный; тело его лежит в вегетативных ядрах черепных нервов или боковых рогах спинного мозга, а нейрит идет к узлу; второй — эфферентный, тело которого лежит в узле, а нейрит достигает рабочего органа.
Эффекторные нейроны симпатических нервов начинаются в ganglia trunci sympathici (узлы первого порядка) или ganglia intermedia (узлы второго порядка), а для парасимпатических нервов — в около- или внутриорганных узлах, ganglia terminalia (третьего порядка); так как в названных узлах осуществляется связь вставочных и эфферентных нейронов, то отмеченная разница между симпатическим и парасимпатическим отделами вегетативной нервной системы связана именно с этими нейронами.
Аксоны эфферентных вегетативных нейронов почти лишены миелина — безмиелиновые (серые). Они составляют послеузловые волокна, rami postganglionics. Послеузловые волокна симпатической нервной системы, отходящие от узлов симпатического ствола, расходятся в двух направлениях. Одни волокна идут к внутренностям и составляют висцеральную часть симпатической системы. Другие волокна образуют rami communicantes grisei, соединяющие симпатический ствол с анимальными нервами.
В составе последних волокна достигают соматических органов (аппарата движения и кожи), в которых иннервируют непроизвольную мускулатуру сосудов и кожи, а также железы.
Совокупность описанных эфферентных вегетативных волокон, идущих от узлов симпатического ствола до органов сомы, составляет соматическую часть симпатического отдела. Такая структура обеспечивает функцию вегетативной нервной системы, которая регулирует обмен веществ всех частей организма применительно к непрерывно изменяющимся условиям среды и условиям функционирования (работы) тех или иных органов и тканей.
Соответственно этой наиболее универсальной своей функции, связанной не с какими-либо отдельными органами и системами, а со всеми частями, со всеми органами и тканями организма, вегетативная нервная система и морфологически характеризуется универсальным, повсеместным распространением в организме.
Следовательно, симпатический отдел иннервирует не только внутренности, но и сому, обеспечивая в ней обменные и трофические процессы.
В результате каждый орган, по И. П. Павлову, находится под тройным нервным контролем, в связи с чем он различает три вида нервов:
1) функциональные, осуществляющие функцию данного органа;
2) сосудодвигательные, обеспечивающие доставку крови к органу, и
3) трофические, регулирующие усвоение из доставленной крови питательных веществ.
Висцеральная часть симпатического отдела содержит все эти три вида нервов для внутренностей, а соматическая часть — только сосудодвигательные и трофические. Что же касается функциональных нервов для органов сомы (скелетная мускулатура и др.), то они идут в составе соматической, ани-мальной, нервной системы.
Таким образом, основное отличие эфферентной части вегетативной нервной системы от эфферентной части анимальной заключается в том, что анимальные, соматические, нервные волокна, выйдя из центральной нервной системы, идут до рабочего органа, нигде не прерываясь, тогда как вегетативные волокна на своем пути от мозга до рабочего органа прерываются в одном из узлов первого, второго или третьего порядка. Вследствие этого эфферентный путь вегетативной нервной системы разбивается на две части, из которых он и состоит: предузловые миелиновые волокна, rami preganglionares, и послеузловые, лишенные миелина (безмиелиновые) волокна, rami postganglionares.
Наличие узлов в эфферентной части рефлекторной дуги составляет характерный признак вегетативной нервной системы, отличающий ее от анимальной.
В основе деятельности нервной системы лежат рефлексы (лат. reflexus — отражённый). Рефлекс - ответная реакция организма на действие раздражителя.
Любой рефлекс существует на базе рефлекторной дуги - совокупности соединенных друг с другом нервных элементов, через которые последовательно проводится нервный импульс при осуществлении рефлекса. Самый прострой пример - коленный рефлекс, который часто проверяет невролог, что позволяет быстро сделать вывод о сохранности элементов рефлекторной дуги.
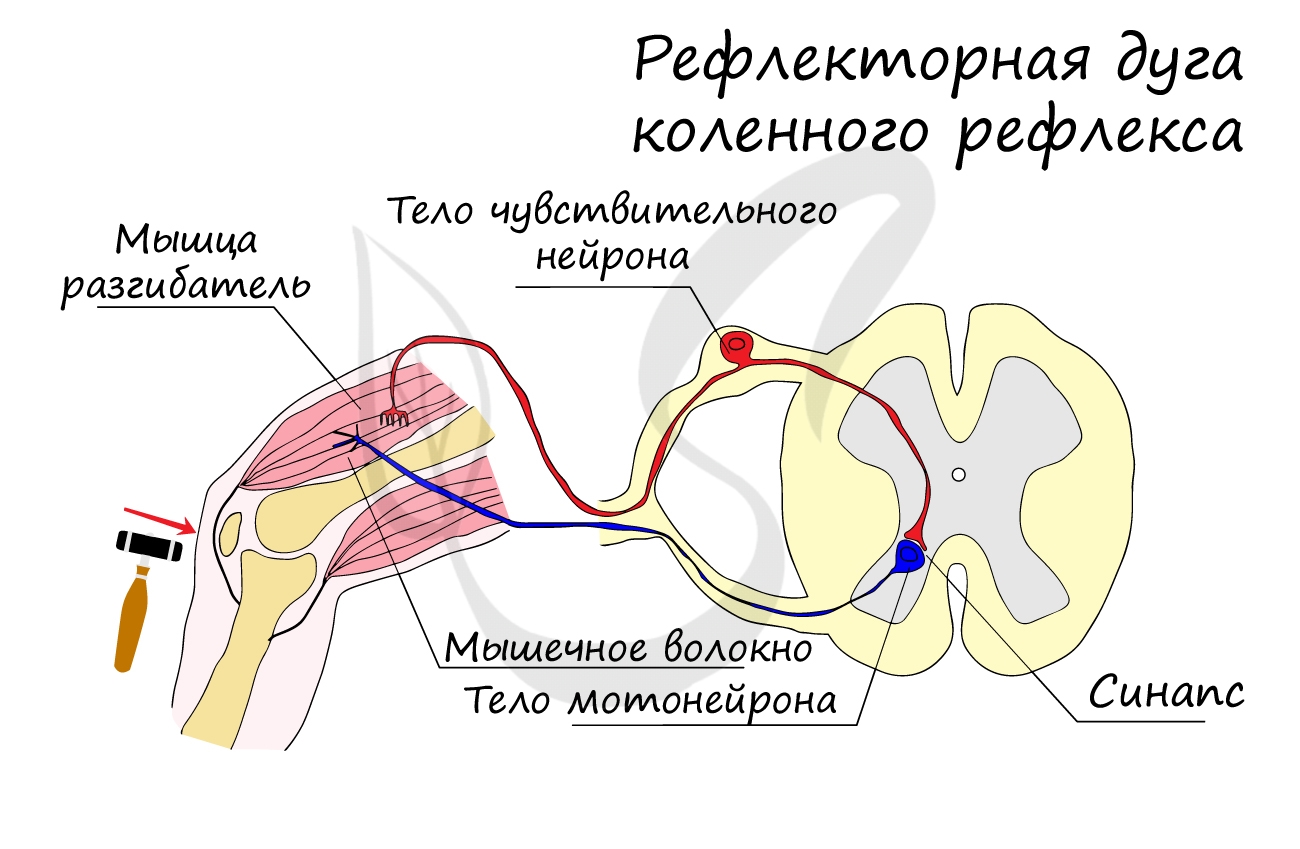
Нейроны соединяются друг с другом с помощью отростков: аксонов и дендритов, на конце которых находятся специальные контакты - синапсы, которые мы подробно изучили в статье про нервные ткани.
Рефлекторные дуги могут быть устроены очень просто: состоять из двух нейронов, подобно рефлекторной дуге коленного рефлекса (отсутствует вставочный нейрон), а могут включать десятки различных нейронов. Рефлекторная дуга может подразделяться на 3 звена:
-
Чувствительное (афферентное, центростремительное)
Состоит из рецептора (может быть расположен в коже, внутренних органах, сосудах) чувствительного нейрона и идущего от этого нейрона чувствительного волокна, которое проникает в спинной мозг через задние рога.
Тело чувствительного нейрона находится в задних корешках (!) спинного мозга. Представили? А теперь представьте дендрит, идущий от кончика вашего указательного пальца до самого спинного мозга. Именно поэтому неверно считать, что дендрит - всегда "короткий" отросток, а аксон - "длинный". Данный вопрос мы обсуждали в статье про нервные ткани.
Вставочное (ассоциативное, промежуточное)
Состоит из вставочного нейрона и его отростков. Вставочный нейрон осуществляет связь между чувствительным и двигательным звеном рефлекторной дуги. Вставочные нейроны могут осуществлять связь с другими отделами ЦНС.
Тела вставочных нейронов находятся в задних рогах спинного мозга.
Двигательное (эфферентное, центробежное)
Представлено двигательным нейроном (эфферентным, исполнительным, мотонейроном), от которого нервные волокна идут к рабочему органу (эффектору, органу-исполнителю).
В зависимости от того, чем представлен эффектор - мышца, железа - при поступлении к нему нервных импульсов его работа активируется: мышца начинает сокращаться, железа - выделять секрет.
Двигательные нейроны лежат в передних рогах спинного мозга, откуда и выходят их отростки.
Рассмотрим схему рефлекторной дуги, на базе которой осуществляется рефлекс отдергивания руки от горячего предмета. Попробуйте сами описать путь, который проходит нервный импульс и вспомнить 3 звена рефлекторной дуги. Назовите локализацию каждого из нейронов.
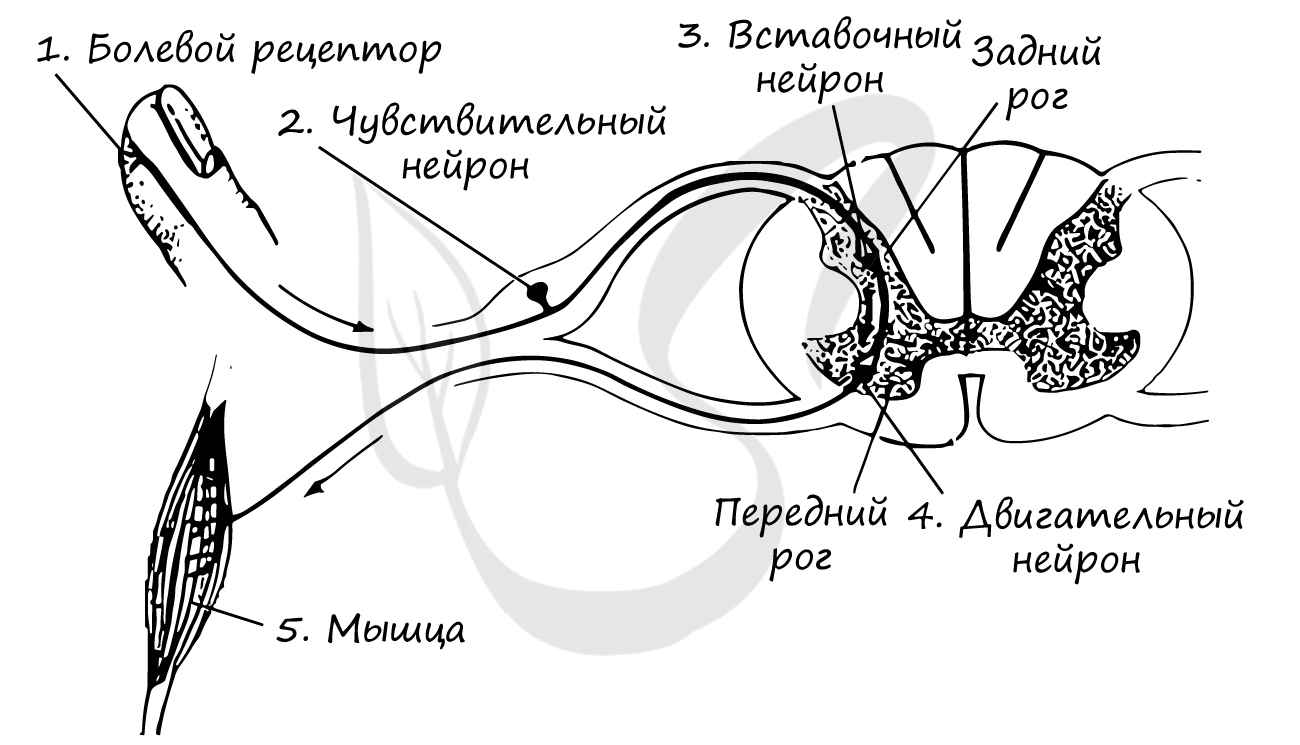
Это может показаться очевидным, но необходимо подчеркнуть, что афферентные нервные волокна входят в спинной мозг через задние корешки. Эфферентные нервные волокна выходят из спинного мозга через передние корешки.
Рефлекторные дуги подразделяются на соматические и вегетативные. С помощью соматических рефлекторных дуг осуществляются двигательные, произвольные рефлексы. С помощью вегетативных - координация деятельности внутренних органов, то есть функции, которые не поддаются нашему осознанному контролю (вспомните вегетативную нервную систему).
Ниже вы увидите схемы соматической и вегетативной рефлекторных дуг. Под картинкой будет написано существенное отличие между ними, которое вы должны запомнить, но прежде постарайтесь сами сделать вывод, изучив картинку.
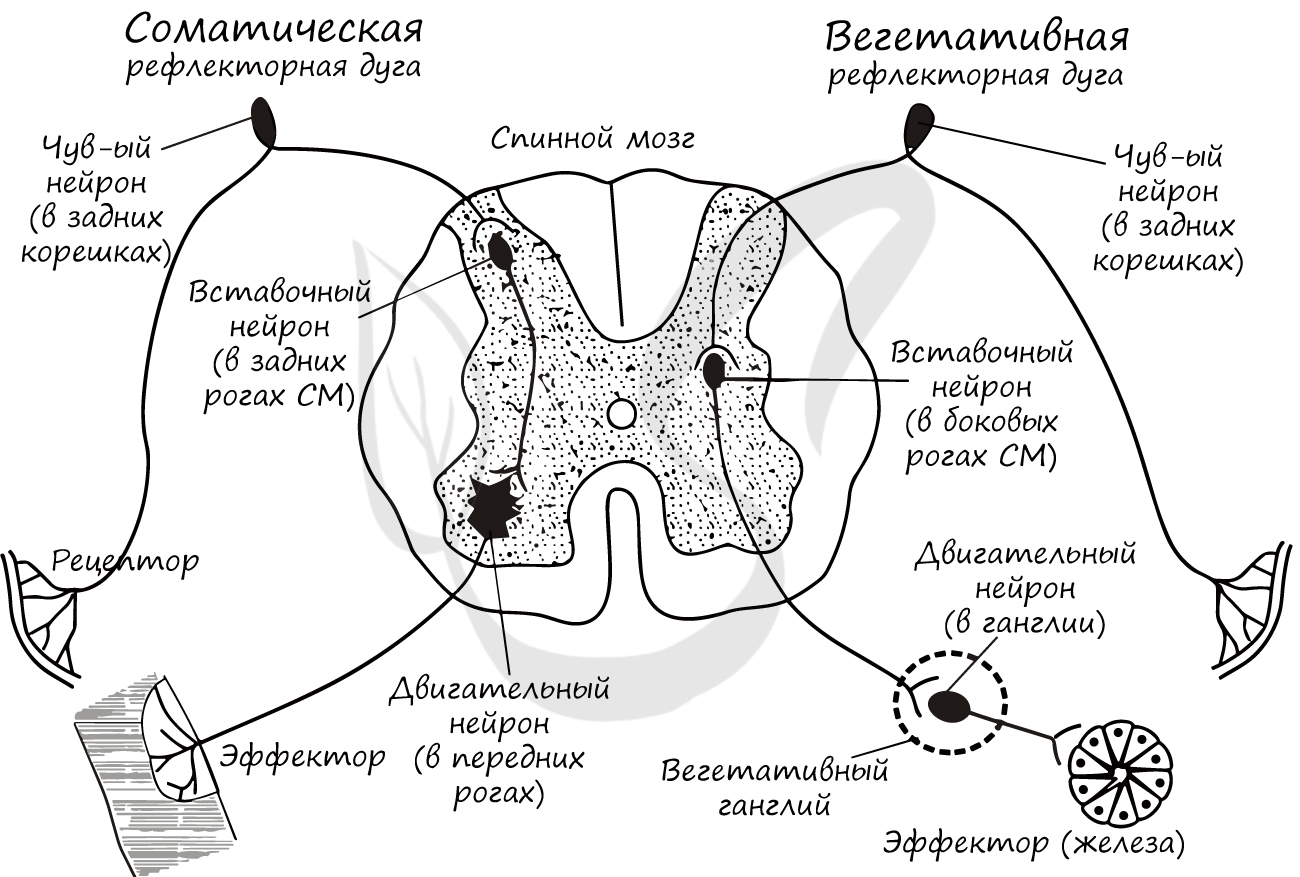
Отличием между соматической и вегетативной рефлекторными дугами в том, что в составе последней эфферентный нейрон лежит за пределами спинного мозга - в вегетативном ганглии. Данные ганглии могут располагаться по бокам от позвоночника, вблизи внутренних органов или в их стенке.
Также вы, скорее всего, обратили внимание, что вставочный нейрон вегетативной дуги локализован в другом месте - в боковых рогах спинного мозга (а не в задних, как в соматической).
Рефлекторная дуга - фундамент, на котором осуществляется рефлекс. В нервной системе возникают не только процессы возбуждения, но и торможения, о которых мы подробнее поговорим в теме, посвященной высшей нервной деятельности. Торможение заключается в ослаблении или задержке уже возникшего возбуждения.
Таким образом, координация и регулирования процессов возбуждения и торможения - основа согласованной работы органов и систем органов, составляющих единый организм.
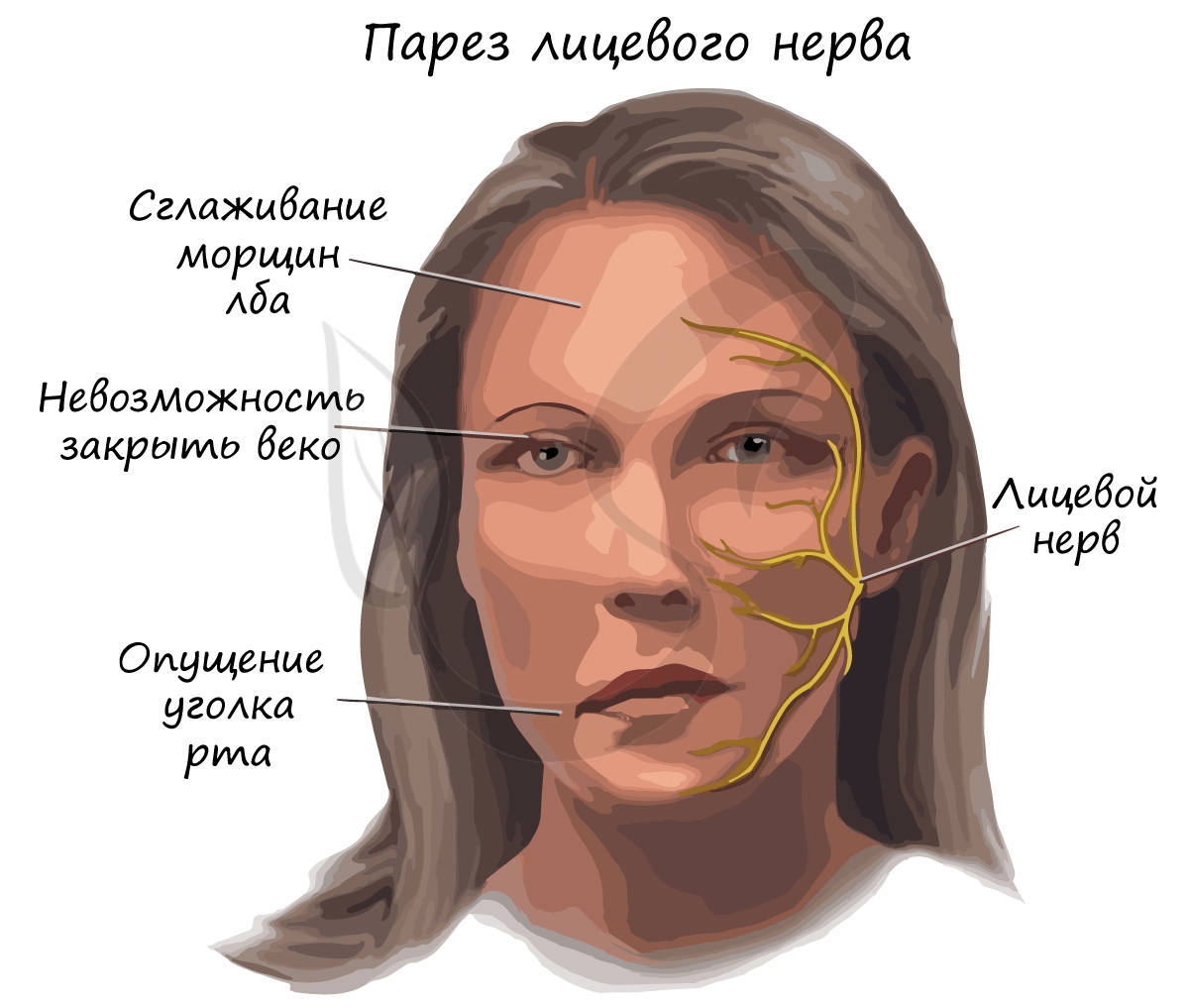
Парез (греч. πάρεσις - ослабление) - неврологический синдром, обусловленный поражением двигательного (эфферентного) пути и слабостью в конечности, или в другом органе, который данный нервный путь иннервировал. Парез проявляется снижением мышечной силы, движения в неполном объеме сохраняются.
Паралич (греч. παράλυσις - расслабление) - полное отсутствие произвольных движений, обусловленное теми же причинами, что и парез.
При переохлаждении может возникнуть парез лицевого нерва. Причиной этому служит воспаление тканей, в результате чего в узком костном канале нерв сдавливается воспаленными тканями. Нервные импульсы частично, либо полностью перестают поступать к мышцам лица, что делает невозможным для пациента движение ими.
Данная статья написана Беллевичем Юрием Сергеевичем и является его интеллектуальной собственностью. Копирование, распространение (в том числе путем копирования на другие сайты и ресурсы в Интернете) или любое иное использование информации и объектов без предварительного согласия правообладателя преследуется по закону. Для получения материалов статьи и разрешения их использования, обратитесь, пожалуйста, к Беллевичу Юрию.
Читайте также: