
Регенерация нервного волокна гистология
Нервные ткани в составе органов относятся к стабильным тканям, так как нейроны митозом не делятся. Физиологическая и репаративная регенерация происходит путем частичной полиплоидизации ядер, восстановления синапсов после их повреждения, роста поврежденных отростков, а главное — путем обновления химических и метаболических компонентов нейронов при внутриклеточном обмене веществ. На месте дефекта в нервной ткани разрастается нейроглия. Она является менее дифференцированной тканью, клетки которой способны делиться митозом. Описаны глиальные клетки, обладающие высокими потенциями к размножению и развитию. Эти клетки принимают активное участие в восстановительных процессах нервной ткани.
При повреждениях, приводящих к нарушению целостности нервных волокон (огнестрельные раны, разрывы), их периферические части распадаются на фрагменты осевых цилиндров и миелиновых оболочек, погибают и фагоцитируются макрофагами (уоллеровская дегенерация осевых цилиндров). В сохранившейся части нервного волокна начинается пролиферация нейролеммоцитов, формирующих цепочку (бюнгнеровская лента), вдоль которой происходит постепенный рост осевых цилиндров. Таким образом, нейролеммоциты являются источником факторов, стимулирующих рост осевого цилиндра. При отсутствии препятствий в виде очагов воспаления и соединительнотканных рубцов возможно восстановление иннервации тканей.
Регенерация нервных отростков идет со скоростью 2-4 мм в сутки. В условиях лучевого воздействия происходит замедление процессов репаративного гистогенеза, что обусловлено в основном повреждением нейролеммоцитов и клеток соединительной ткани в составе нерва. Способность нервных волокон к регенерации после повреждения при сохранении целостности тела нейрона используется в микрохирургической практике при сшивании дистального и проксимального отростков поврежденного нерва. Если это невозможно, то используют протезы (например, участок подкожной вены), куда вставляют концы поврежденных нервов (футлериз). Регенерацию нервных волокон ускоряет фактор роста нервной ткани — вещество белковой природы, выделенное из тканей слюнных желез и из змеиного яда.
Важнейшее свойство живого — отвечать комплексом изменений метаболизма, двигательной активности, размножения или гибели и др. на внешние и внутренние воздействия. Восприятие, трансформация и передача последних осуществляется рецепторно-трансдукторной системой клетки, элементы которой связаны с внутренней средой клетки. На клеточном уровне организации живого первично формируется каскад реакций, распространяющихся на тканевый, органный и организменный уровни.
Под реактивностью тканей с гистогенетических позиций следует понимать изменения основных закономерных сторон развития ткани — пролиферации, дифференцировки, интеграции клеток, межклеточных взаимодействий и других закономерных процессов гистогенеза, происходящих под действием внешних для ткани факторов. При самых разнообразных воздействиях (травмы, ожоги, стрессовые ситуации и т. п.) ткани, для которых в норме характерно клеточное обновление путем митоза, реагируют прежде всего понижением или повышением степени пролиферативной активности клеток. Угнетение митозов или, наоборот, "вспышки" митотической активности — непременные показатели реактивности таких тканей. В некоторых тканях результатом реактивного изменения пролиферации клеток являются эндомитоз и амитоз, образование двуядерных и многоядерных клеток.
Наряду с повышением уровня дифференцировки клеток, реактивность тканей может характеризоваться и явлениями дедифференцировки клеток. Дедифференцировка клеток — это упрощение их внутренней структуры, связанное с временным снижением уровня дифференцировки (специализации) клеток. Дедифференцированные клетки приобретают способность к пролиферации. В них активизируется синтез ДНК и общих неспецифических белков. Дедифференцировка как реактивно-приспособительное изменение клеток сопровождается увеличением относительных объемов ядер и ядрышек, увеличением числа свободных рибосом, исчезновением специальных органелл и включений в цитоплазме, редукцией мембран комплекса Гольджи, уменьшением числа митохондрий и другими признаками. Под дедифференцировкой следует понимать структурно-адаптационную реакцию клетки, а не обратное ее развитие.
При изменении функциональной нагрузки на ткани реактивно-приспособительные изменения клеток проявляются в метаболических сдвигах — трофических нарушениях типа гипертрофии, дистрофии, атрофии, гипо- или гиперсекреции и т. д.
При гипертрофии клеток наблюдается гиперплазия внутриклеточных структур (митохондрий, эндоплазматической сети, органелл) как морфологический эквивалент повышенной функциональной активности.
Реактивность тканей характеризуется также изменениями межклеточных взаимодействий. При действии повреждающих факторов в тканях может возникнуть состояние дезинтеграции клеток (например, нарушение межнейронных связей при интоксикациях, дискомплексации эпителиев при нарушении контактов между эпителиоцитами и т. д.). Как проявление реактивности тканей следует рассматривать программированную гибель клеток за счет активации внутренней программы самоуничтожения или ее запуска внешними стимулами, например факторами, вырабатываемыми клетками микроокружения.
Реактивные изменения тканей зависят, конечно, во многом от силы раздражителя — повреждающего фактора. Существенное значение в реактивности тканей имеет наследственная природа клеток самой ткани, так как каждая ткань отличается генетически детерминированным диапазоном изменчивости или нормы адаптивной реакции, обеспечивающей нормальное функционирование ткани. После воздействия повреждающих факторов реактивные изменения клеток могут выходить за пределы диапазона изменчивости, характерного для нормального функционирования ткани. Однако обычно резко выраженные вначале реактивные изменения клеток постепенно сглаживаются, и структурно-функциональные свойства их приближаются к норме, обеспечивая адаптацию ткани к функционированию в новых условиях.
Это отростки нейронов, покрытые глиальными оболочками. Отростки нейронов лежат внутри нервных волокон и называются осевыми цилиндрами. Их окружают глиальные клетки – олигодендроциты, которые здесь называются леммоцитами (оболочечными клетками), или шванновскими клетками. Нервные волокна бывают миелиновые и безмиелиновые (рис. 8-5).
Миелиновые нервные волокна. Они толще безмиелиновых, содержат только по одному осевому цилиндру (2-20 мкм). (Рис.8-5а). Поскольку миелин представляет собой обмотку из липидных мембран, на гистологических препаратах
Р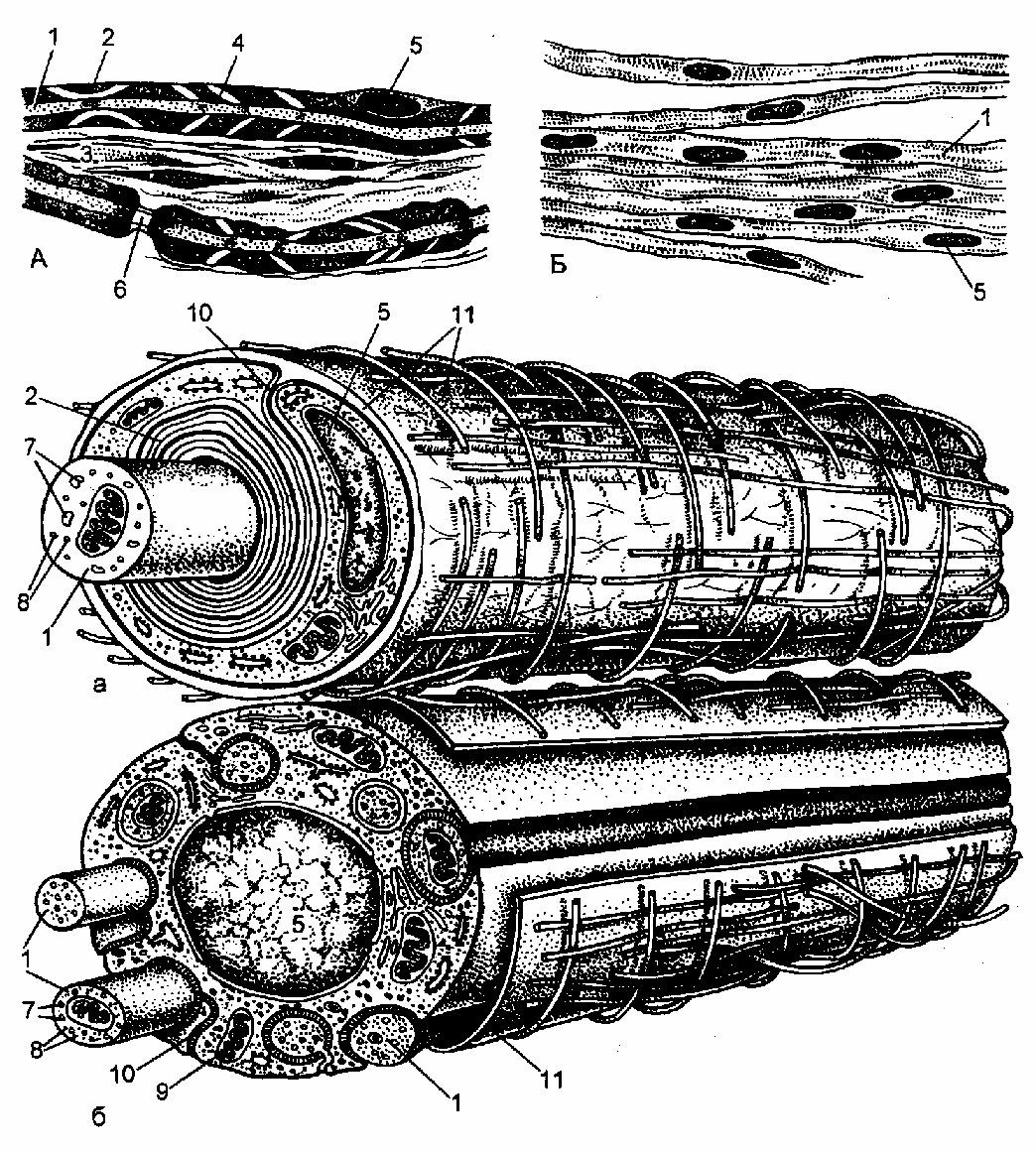
ис. 8-5. Строение нервных волокон на светооптическом (А, Б) и ультрамикроскопическом (а, б) уровнях.
А,а. Миелиновое волокно. Б,б. Безмиелиновое волокно. 1. Осевые цилиндры. 2. Миелиновый слой. 3. Соединительная ткань. 4. Насечка миелина. 5. Ядро нейролеммоцита. 6. Узловой перехват. 7. Микротрубочки. 8. Нейрофиламенты. 9. Митохондрии. 10. Мезаксон. 11. Базальная мембрана. (По Т. Н. Радостиной).
он хорошо импрегнируется осмиевой кислотой. Остатки цитоплазмы леммоцитов сохраняются между витками мезаксона, образуя насечки миелина. Они не окрашиваются осмиевой кислотой и поэтому видны на фоне миелина в виде косых светлых полос. Поскольку в процессе образования миелиновой оболочки осевой цилиндр и леммоциты продолжают расти, то каждый последующий, наружный виток мезаксона шире предыдущего. Участки волокна, не покрытые миелином, называются узловыми перехватами. Они находятся на границе между двумя соседними леммоцитами. Соответственно, участок волокна образованный одной глиальной клеткой именуется межузловым сегментом. Снаружи всё волокно, включая узловые перехваты, покрыто базальной мембраной. По миелиновым волокнам нервный импульс передается с большой скоростью, до 120 м/сек.
Безмиелиновые нервные волокна. Они состоят из тяжа леммоцитов, которые содержат несколько (10-20) осевых цилиндров, погруженных в леммоциты. Поэтому безмиелиновые нервные волокна называют волокнами кабельного типа. Каждый из них как бы подвешен на сдвоенной мембране (мезаксон), как на брыжейке. Такие волокна чаще встречаются в вегетативной нервной системе. Нервный импульс по ним проводится медленно (1-2 м/сек). (Рис. 8-5б).
Регенерация нейронов и нервных волокон
Нейроны взрослых человека и животных не способны к делению, клеточной регенерации. Однако у них хорошо развита внутриклеточная регенерация: обновление макромолекул и органелл. При гибели одних нейронов, сохранившиеся нейроны гипертрофируются и берут на себя функции погибших. Возможно также восстановление повреждённых отростков нейронов и, соответственно, регенерация периферических нервов.
После перерезки нервного волокна, наступает дегенерация осевого цилиндра дистальней места повреждения. Леммоциты и макрофаги фагоцитируют продукты распада, очищают место провреждения, а затем размножаются и образуют тяжи – ленты Бюнгера. На проксимальном отрезке осевого цилиндра образуется наплыв аксоплазмы - формируется колба роста (как в эмбриогенезе). Осевой цилиндр растёт по дорожке из леммоцитов со скоростью 2-4 мм в сутки до тех пор, пока не достигает иннервируемого органа. После этого вокруг новообразованного осевого цилиндра леммоциты образуют миелиновую оболочку, а в рабочем органе вновь формируется (восстанавливается) нервное окончание. Эти процессы завершаются в течение нескольких месяцев от момента повреждения. Однако, если возникает препятствие на пути роста осевых цилиндров, они начинают расти беспорядочно и образуют клубок, называемый ампутационной невромой. При её раздражении возникает сильная боль, которая воспринимается как происходящая из первоначально иннервируемой области, например как боль в удалённой конечности (фантомные боли). Нервные окончания
Все нервные волокна заканчиваются концевыми аппаратами, называемых нервными окончаниями. По функции они делятся на эффекторные, рецепторные и межнейрональные синапсы.
Нервный импульс в организме человека обычно передаётся с одной нервной клетки на другую или с нейрона на рабочий орган через медиатор, химический посредник. Медиатор взаимодействует со специфическими рецепторами другого нейрона или клетками рабочего органа и через целый каскад вторичных внутриклеточных посредников, меняет функцию рабочего органа или другого нейрона.
4.1. Общая характеристика нервной ткани
Нервная ткань — это система взаимосвязанных нервных клеток и нейроглии, обеспечивающих специфические функции восприятия раздражений, возбуждения, выработки импульса и его передачи. Нервные клетки (нейроны) — основные структурные компоненты нервной ткани, выполняющие специфическую функцию. Нейроглия обеспечивает существование и функционирование нервных клеток, осуществляя опорную, трофическую, разграничительную, секреторную и защитную функции. Источником развития нервной ткани служит дорсальная эктодерма. [1],[5]
Из нервной трубки в дальнейшем формируются нейроны и макроглия центральной нервной системы. Нервный гребень дает начало нейронам чувствительных и автономных ганглиев и некоторым видам глии: нейролеммоцитам (шванновским клеткам), клеткам-сателлитам ганглиев. Нервная трубка на ранних стадиях эмбриогенеза представляет собой многорядный нейроэпителий, состоящий из вентрикулярных, или нейроэпителиальных клеток. [1],[5]
В дальнейшем в нервной трубке дифференцируется 4 концентрических зоны: внутренняя - вентрикулярная (или эпендимная) зона, вокруг нее – субвентрикулярная зона, затем промежуточная (или мантийная зона) и, наконец, наружная - краевая (или маргинальная) зона нервной трубки. Вентрикулярная (эпендимная), внутренняя, зона состоит из делящихся клеток цилиндрической формы. Вентрикулярные (или матричные) клетки являются предшественниками нейронов и клеток макроглии. Субвентрикулярная зона состоит из клеток, сохраняющих высокую пролиферативную активность и являющихся потомками матричных клеток. Промежуточная (плащевая, или мантийная) зона состоит из клеток, переместившихся из вентрикулярной и субвентрикулярной зон — нейробластов и глиобластов. Нейробласты утрачивают способность к делению и в дальнейшем дифференцируются в нейроны. Глиобласты продолжают делиться и дают начало астроцитам и олигодендроцитам. [1], [5]
Способность к делению не утрачивают полностью и зрелые глиоциты. Новообразование нейронов прекращается в раннем постнатальном периоде. Из клеток плащевого слоя образуются серое вещество спинного и часть серого вещества головного мозга. Маргинальная зона (или краевая вуаль) формируется из врастающих в нее аксонов нейробластов и макроглии и дает начало белому веществу.[5]
4.2. Регенерация нервной ткани
Рассматривая процессы регенерации в нервных тканях следует сказать, что нейроциты являются наиболее высокоспециализированными клетками организма и поэтому утратили способность к митозу. Физиологическая регенерация (восполнение естественного износа) в нейроцитах хорошая и протекает по типу "внутриклеточной регенерации" - т.е. клетка не делится, но интенсивно обновляет изношенные органоиды и другие внутриклеточные структуры. Для этого в нейроцитах хорошо выражены гранулярный ЭПС, пластинчатый комплекс и митохондрии, т.е. имеется мощный синтетический аппарат для синтеза органических компонентов внутриклеточных структур.[5]
К компенсаторно-приспособительным процессам в нервной ткани относится обнаружение многоядрышковых, двухъядерных и гипертрофированных нервных клеток при различного рода болезнях, сопровождающихся дистрофическими процессами, при условии сохранения общей структуры нервной ткани. Нервные клетки вегетативной нервной системы восстанавливаются путем гиперплазии органелл, а также не исключается возможность их размножения. Периферические нервы являются в большинстве своем смешанными и состоят из двигательных волокон передних корешков (аксонов клеток передних рогов), чувствительных волокон (дендритов клеток межпозвонковых узлов) и вазомоторно-секреторно-трофических волокон (симпатических и парасимпатических) от соответствующих клеток серого вещества боковых рогов спинного мозга и ганглиев симпатического пограничного ствола. Нервное волокно, входящее в состав периферического нерва, состоит из осевого цилиндра, расположенного в центре волокна, миелиновой или мякотной оболочки, одевающей осевой цилиндр и шванновской оболочки. Миелиновая оболочка нервного волокна местами прерывается, образуя так называемые перехваты Ранвье. В области перехватов осевой цилиндр прилежит непосредственно к шванновской оболочке. Миелиновая оболочка обеспечивает роль электрического изолятора, предполагается ее участие в процессах обмена осевого цилиндра. Шванновские клетки имеют общее происхождение с нервными элементами. Они сопровождают осевой цилиндр периферического нервного волокна подобно тому, как глиозные элементы сопровождают осевые цилиндры в центральной нервной системе, поэтому шванновские клетки иногда называют периферической глией. На месте дефекта в нервной ткани разрастается нейроглия. Она является менее дифференцированной тканью, клетки которой способны делиться митозом. Существуют глиальные клетки, обладающие высокими потенциями к размножению и развитию. Эти клетки принимают активное участие в восстановительных процессах нервной ткани. Наиболее частыми формами травматического повреждения нервов, возникающими вследствие техногенного травматизма на производстве, при дорожно- транспортных происшествиях, в ходе военных действий, являются размозжение, ушиб, растяжение, а также сдавление с наличием или отсутствием разрыва нервного ствола. Однако эффективность репарации структуры и функции поврежденной ткани с применением лечебных мероприятий и лекарственных средств остается относительно низкой. Это во многом связано с малой изученностью динамики регенерации нервов после травмы. Для исследования воздействия модулирующих средств на посттравматический процесс необходимы более полные данные о динамике репаративной регенерации поврежденного нерва.[5]
При повреждениях, приводящих к нарушению целостности нервных волокон, их периферические части распадаются на фрагменты осевых цилиндров и миелиновых оболочек, погибают и фагоцитируются макрофагами. В сохранившейся части нервного волокна начинается пролиферация нейролеммоцитов, формирующих цепочку (бюнгнеровская лента), вдоль которой происходит постепенный рост осевых цилиндров. [5],[6]
Выделяемые шванновской клеткой различные стимуляторы (нейтрофические факторы) поглощаются аксоном и ретроградно транспортируются в перикардион. В перикарионе эти факторы стимулируют синтез белка и поддерживают его на высоком уровне. В регенерирующем нерве шванновские клетки пролиферируют, синтезируют компоненты базальной мембраны, внеклеточного матрикса и формируют миелин. Шванновские клетки стимулируют удлинение аксона и контролируют его направленный рост и мишени. При отсутствии Шванновских клеток аксоны не могут расти на значительные расстояния. [5]
Восстановление утраченных связей может происходить и за счет образования коллатеральных ветвей из окружающих и неповрежденных нервных волокон. Чаще коллатеральные ветви отходят от участка аксона в области перехвата Ранвье. Наличие в зоне перерезки нерва мертвых тканей, которые стимулируют разрастание здесь рубцовой ткани, большое расстояние между отрезками нервного волокна, сильное повреждение сосудов и нарушение кровоснабжения нерва ведут к резкому нарушению его регенерации. [5],[6]
Разрастание рубцовой ткани иногда вызывает развитие ампутационной невромы, состоящей из разросшихся отростков нейронов и глии, окруженных грубой рубцовой тканью. Невромы могут вызывать сильные (фантомные) боли. Регенерация нервных отростков идет со скоростью 2-4 мм в сутки. В условиях лучевого воздействия происходит замедление процессов репаративного гистогенеза, что обусловлено в основном повреждением нейролеммоцитов и клеток соединительной ткани в составе нерва. Способность нервных волокон к регенерации после повреждения при сохранении целостности тела нейрона используется в микрохирургической практике при сшивании дистального и проксимального отростков поврежденного нерва. Если это невозможно, то используют протезы (например, участок подкожной вены), куда вставляют концы поврежденных нервов (футлериз). Регенерацию нервных волокон ускоряет фактор роста нервной ткани — вещество белковой природы, выделенное из тканей слюнных желез и из змеиного яда. Нервная ткань на повреждение реагирует неоднозначно. Повреждение клеток центральной нервной системы, нейронов спинного мозга, симпатических ганглиев завершается их гибелью. Аксоны же нервных клеток сохраняют способность к репаративной регенерации. Повреждение периферического нерва сопровождается дегенерацией и атрофией конца нерва, идущего к периферии. Регенерация начинается на конце аксона, связанного с нервной клеткой. Регенерирующий конец нерва врастает в трубочки и способен восстановить иннервацию. Если же аксон не совмещен с объектом врастания, то на его конце могут образоваться своеобразные утолщения — невромы. Эффективность процесса регенерации во многом определяется условиями, в которых он протекает.[5],[6]
Значение имеет общее состояние организма. Так, истощение, авитаминозы, нарушения иннервации затормаживают репаративную регенерацию и способствуют ее переходу в патологическую. Изменение условий, в которых протекает процесс регенерации, может приводить как к количественным, так и качественным его изменениям.[5]
4.3. Возрастные особенности периферической нервной системы.
Развитие нервной системы у детей с моментом рождения не останавливается. После рождения увеличивается количество нервных пучков в составе периферических нервов: усложняется их ветвление, расширяются межнейрональные связи, усложняются рецепторные аппараты.[5]
С возрастом увеличивается толщина нервных волокон. К 9 годам во всех периферических нервах миелинизация близится к завершению. В пожилом и старческом возрасте количество нейронов в спинномозговых ганглиях снижается на 30%, часть нейронов атрофируется. Возрастные изменения в нервной ткани связаны с утратой нейроцитов в постнатальном периоде способности к делению, и как следствие этого постепенным уменьшением количества нейроцитов, особенно чувствительных нейроцитов, а также уменьшением уровня метаболических процессов в оставшихся нейроцитах. Отсутствие клеточной формы регенерации нейроцитов обуславливает разрастание нейроглии и соединительной ткани на месте повреждения.[5]
Заключение
Таким образом, на основании проанализированной литературы, можно сделать вывод о том, что нервная ткань обладает плохой способностью к регенерации. В эксперименте показано, что клетки периферической и вегетативной нервной системы, двигательные и чувствительные нейроны в спинном мозге мало регенерируют.[5]
Регенерация нервной ткани может происходить путём роста тканей на раневой поверхности, перестройки оставшейся части органа в новый, или путём роста остатка органа без изменения его формы. [2],[5]
Уровни регенерации в ходе восстановления структур следующие: молекулярный, ультраструктурный, клеточный, тканевой, органный.[5]
Новые нейроны образуются из недифференцированных предшественников, которые способны давать начало также астроцитам и олигодендроцитам и поэтому могут рассматриваться как стволовые нервные клетки. Идентификация стволовых клеток в мозге представляет большие сложности особенно в зрелом возрасте. [2],[5]
Сочетание классических хирургических способов восстановления нервных стволов и методов прямой терапии ростовыми факторами ускоряет аксональный рост, стимулирует реваскуляризацию нерва, что подтверждается многочисленными экспериментальными исследованиями и, в итоге, улучшает результаты посттравматического восстановления функции реиннервации поврежденного органа или ткани. [5], [6]
Список литературы
1. Гистология, эмбриология, цитология: учебник / Ю. И. Афанасьев, Н. А. Юрина, Б. В. Алешин и др.; под ред. Ю. И. Афанасьева, Н. А. Юриной. - 6-е изд., перераб. и доп. – М.: ГЭОТАР-Медиа, 2014. - 800 с.
2. Корочкин Л. И. Что такое стволовые клетки // Природа. – 2005. - № 6. – С. 3-11.
3. Кузнецов С.Л., Мушкамбаров Н.Н.Гистология, цитология и эмбриология: Учебник. – М.: Медицинское Информационное Агентство, 2007. – 540с.
4. Руководство по гистологии/ под ред. Р.К. Данилов. – СПб.: СпецЛит.- 2011.–Том 1.-С.160-165; 190-195; 210-216; 252-258; 280-299; 440-442; 656-669.
5. Регенерация тканей: учебное пособие – Благовещенск, 2016. – 136 с.
Дата добавления: 2018-02-15 ; просмотров: 2317 ;
Нейроны являются несменяемой клеточной популяцией. Им свойственна только внутриклеточная физиологическая регенерация, заключающаяся в непрерывной смене структурных белков цитоплазмы.
Отростки нейронов и соответственно периферические нервы обладают способностью к регенерации в случае их повреждения. При этом регенерации нервных волокон предшествуют явления дегенерации. Нейролеммоциты периферического отрезка волокна уже в первые сутки резко активизируются. В цитоплазме нейролеммоцитов увеличивается количество свободных рибосом и полисом, эндоплазматической сети. В цитоплазме нейролеммоцитов образуется значительное количество шарообразных слоистых структур различных размеров. Миелиновый слой как обособленная зона нейролеммоцита исчезает. В течение 3-4 суток нейролеммоциты значительно увеличиваются в объеме. Нейролеммоциты интенсивно размножаются, и к концу 2-й недели миелин и частицы осевых цилиндров рассасываются. В резорбции продуктов принимают участие как глиальные элементы, так и макрофаги соединительной ткани.
Осевые цилиндры волокон центрального отрезка образуют на концах булавовидные расширения — колбы роста и врастают в лентовидно расположенные нейролеммоциты периферического отрезка нерва и растут со скоростью 1-4 мм в сутки. Рост нервных волокон замедляется в области терминалей. Позднее происходит миелинизация нервных волокон и восстановление терминальных структур.
Нервные окончания
Все нервные волокна заканчиваются концевыми аппаратами, которые получили название нервные окончания.
По функциональному значению нервные окончания можно разделить на три группы:
Эффекторные нервные окончания представлены двумя типами: двигательные и секреторные.
Двигательные нервные окончания — это концевые аппараты аксонов двигательных клеток соматической или вегетативной нервной системы. При их участии нервный импульс передается на ткани рабочих органов. Двигательные окончания в поперечно-полосатых мышцах называются нервно-мышечными окончаниями (моторная бляшка). Они представляют собой окончания аксонов клеток двигательных ядер передних рогов спинного мозга или моторных ядер головного мозга. Нервно-мышечное окончание состоит из концевого ветвления осевого цилиндра нервного волокна и специализированного участка мышечного волокна. Миелиновое нервное волокно, подойдя к мышечному волокну, теряет миелиновый слой и погружается в мышечное волокно, вовлекая за собой его плазмолемму. Соединительнотканные элементы при этом переходят в наружный слой оболочки мышечного волокна. Плазмолеммы терминальных ветвей аксона и мышечного волокна разделены синаптической щелью шириной около 50 нм.
В области окончания мышечное волокно не имеет типичной поперечной исчерченности и характеризуется обилием митохондрий, скоплением круглых или слегка овальных ядер. Саркоплазма с митохондриями и ядрами в совокупности образует постсинаптическую часть синапса.
Терминальные ветви нервного волокна в мионевральном синапсе характеризуются обилием митохондрий и многочисленными пресинаптическими пузырьками, содержащими характерный для этого вида окончаний медиатор — ацетилхолин. При возбуждении ацетилхолин поступает через пресинаптическую мембрану в синаптическую щель на холинорецепторы постсинаптической (мышечной) мембраны, вызывая ее возбуждение (волну деполяризации).
Постсинаптическая мембрана моторного нервного окончания содержит фермент ацетилхолинэстеразу, разрушающий медиатор и ограничивающий этим срок его действия. Двигательные нервные окончания в гладкой мышечной ткани построены проще. Здесь тонкие пучки аксонов или их одиночные терминали, следуя между мышечными клетками, образуют четкообразные расширения (варикозы), содержащие холинергические или адренергические пресинаптические пузырьки.
Секреторные нервные окончания имеют простое строение и заканчиваются на железе. Они представляют собой концевые утолщения, или четковидные расширения волокна с синаптическими пузырьками, содержащими главным образом ацетилхолин.
12.1. Общие сведения
12.1.1. Функции клеток нервной ткани
12.1.1.1. Нейроны
I. Функции
Нервные клетки обладают 4-мя важнейшими свойствами.
б) Каждый вид нейронов настроен на восприятие строго определённых сигналов -
б) За счёт этого сигнал проходит большее или меньшее расстояние.
в) Так, определённые нейроны спинномозговых узлов с помощью своих отростков проводят сигналы
II. Способы передачи сигнала
Передача сигнала может происходить двумя способами.
12.1.1.2. Глия
12.1.2. Развитие нервной ткани
3. В процессе развития в перечисленных на схеме эмбриональных органах (нервной трубке, нервном гребешке и нейральных плакодах) образуются два типа бластных клеток . -
12.2. Нейроны
12.2.1. Подразделение по функции
12.2.1.1. Три типа нейронов
По функции нейроциты делятся на 3 вида:
Б. Эти сигналы передаются
б) Тела нейронов находятся всегда в ганглиях (т.е. вне центральной нервной системы) - в
т.е в спинном или головном мозгу (*) , где участвуют в замыкании центральных рефлекторных дуг,
б) Тела данных клеток находятся
(*) Правильно говорить: " в мозгу ", а не "в мозге".
12.2.1.2. Три типа проводящих путей
а) Отростки перечисленных нейронов могут образовывать проводящие пути, которые тоже делят на три вида.
б) Однако тип проводящих путей не всегда совпадает с типом образующих их нейронов.
б) Таким образом, в образовании этих путей принимают участие
б) В образовании этих путей участвуют
а) По форме и размерам нейроциты очень различны.
б) В нейроците выделяют тело ( перикарион ) и отростки.
12.2.2. Отростки нейронов
12.2.2.1. Дендриты и аксоны
Среди отростков нейронов различают дендриты и аксоны.
12.2.2.2. Подразделение нейронов по числу отростков
По общему количеству отростков нейроны и их предшественники делятся на несколько видов.
б) Таковыми являются
и кажется, будто клетка имеет всего один отросток,
Б. Следовательно, данные нейроны имеют
в) Большая длина дендрита обусловлена тем, что он должен обеспечивать проведение сигнала
б) Таковыми являются
12.2.3. Просмотр препаратов: общий вид нейронов
12.2.3.1. Мультиполярные нейроциты
а) На данном снимке видны нейроны

а) При данном методе окраски нейрон

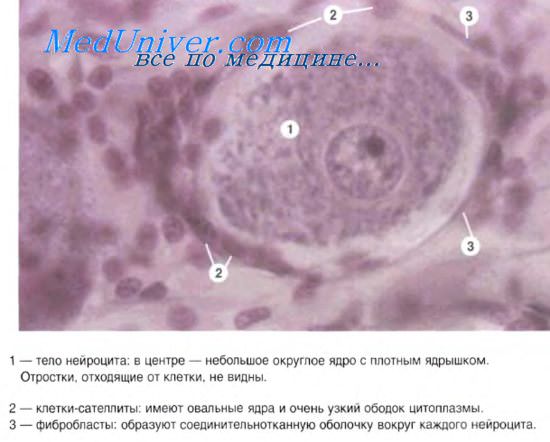
12.2.3.2. Псевдоуниполярные нейроциты
б) Они окружены многочисленными мелкими глиальными клетками-сателлитами (2) .
в) Видны также нервные волокна (3) , образованные

Б. Отростки, отходящие от клетки, не видны.
б) Клетки-сателлиты (2) имеют

12.2.4. Цитоплазма нейроцитов
12.2.4.1. Специфические структуры цитоплазмы
а) Способность нейронов к возбуждению и его проведению связана с наличием в их плазмолемме систем транспорта ионов -
12.2.4.2. Базофильное вещество
б) Оно находится

в которой интенсивно происходит белковый синтез.
12.2.4.3. Нейрофибриллы
б) Они находятся также

12.2.4.4. Нейросекреторные гранулы


б) Поэтому, кроме тела нейрона, секреторные гранулы могут обнаруживаться
12.2.5. Дополнительные вопросы
12.2.5.1. Схема строения нейрона
2. а) Изображённая клетка имеет
б) Во всех отростках содержатся параллельно расположенные
3. В теле клетки показаны органеллы:
4. Видно также, что к нейрону подходят аксоны многих других нейронов, образуя
12.2.5.2. Транспорт веществ по отросткам нейронов
12.3. Нейроглия
Нейроглию подразделяют следующим образом.
12.3.1. Олигодендроглия и периферическая нейроглия
12.3.1.1. Виды и функциональная роль
а) У олигодендроглиоцитов отростки -
12.3.1.2. Препарат
б) При этом в поле зрения - часть тела псевдоуниполярного нейрона (1) - в том числе его ядро.
2. а) Клетки-сателлиты (2)

12.3.2.1. Виды и функциональная роль
б) Толщина и длина отростков зависит от типа астроглии.
12.3.2.2. Препарат
12.3.3. Эпендимная глия
12.3.3.1. Основные сведения
б) А. Эти клетки можно рас с матриват ь как разновидность эпителия ( п. 7.1.1 ).
Б. Однако, в отличие от других видов эпителия,
Б. Он заполнен жидкостью и выстлан эпендимой (2) .
а) Малое увеличение 
б) Тем не менее, отсутствие между ними плотных контактов позволяет жидкости

12.3.3.2. Отростки клеток
б) А. Отростки имеются не у всех эпендимоцитов.
Б. Эпендимоциты с отростками называются таницитами .
В. Особенно многочисленны танициты в дне III желудочка.
в) По-видимому, отростки выполняют

12.4. Нервные волокна
12.4.1. Общие замечания
б) Сам же отросток нейрона, находящийся в составе волокна, называется
12.4.2. Безмиелиновые нервные волокна
12.4.2.1. Принцип строения

12.4.2.2. Просмотр препарата
I. Световая микроскопия
II. Электронная микроскопия
2. Под электронным микроскопом строение каждого из них соответствует вышеприведённому описанию:
12.4.3. Миелиновые нервные волокна
12.4.3.1. Принцип строения
I. Поперечное сечение

II. Продольное сечение: перехваты Ранвье
здесь остаётся только истончённая не й р о лемма.
а в тех участках цилиндра, которые покрыты миелиновой оболочкой, каналов нет.
б) Такое расположение Na + -каналов
12.4.3.2. Различия между безмиелиновыми
и миелиновыми волокнами
Различия в строении двух типов волокон сведены в таблицу.-
| Безмиелиновые нервные волокна | Миелиновые нервные волокна |
| 1. Обычно - несколько осевых цилиндров , располагающихся по периферии волокна. | 1. Один осевой цилиндр находится в центре волокна. |
| 2. Осевые цилиндры - это, как правило, аксоны эфферентных нейронов вегетативной нервной системы. | 2. Осевой цилиндр может быть как аксоном, так и дендритом нейроцита. |
| 3. Ядра олигодендроцитов находятся в центре волокон. | 3. Ядра и цитоплазма леммоцитов оттеснены к периферии волокна. |
| 4. Мезаксоны осевых цилиндров - короткие. | 4. Мезаксон многократно закручивается вокруг осевого цилиндра, образуя миелиновый слой . |
| 5. Na + -каналы располагаются по всей длине осевого цилиндра. | 5. Na + -каналы - только в перехвате Ранвье. |
12.4.3.3. Просмотр препаратов
I. Световая микроскопия: поперечный срез

II. Световая микроскопия: продольный срез

б) В этих местах концентрические листки мезаксона не так плотно прилегают друг к другу, отчего между ними сохраняются
III. Электронная микроскопия: продольный срез, перехват Ранвье
1. На снимке - миелиновое волокно в месте стыка соседних леммоцитов.
2. В центре волокна - осевой цилиндр (1) с обычными структурами:
4. а) Но в средней части снимка - в месте стыка леммоцитов - миелиновый слой сходит на нет, а нейролемма истончается:
IV. Электронная микроскопия: продольный срез, насечки миелина
1. Здесь увеличение почти в 10 раз больше, чем на предыдущем снимке.
2. Осевой цилиндр - правая светлая область снимка;
цифрой 1 обозначена его плазматическая мембрана ( аксолемма ).
3. а) Тёмные слоистые образования - миелиновая оболочка (многократно закрученный мезаксон).
б) Но в этой оболочке - светлое разрежение:
Читайте также: