
Секреция поджелудочного сока регулируется и нервным и гуморальным путями
Слабое сокоотделение натощак резко усиливается во время (через 2— 3 мин) и после приема пищи. Панкреатическое сокоотделение начинается уже на вид, запах пищи как условно-рефлекторный механизме секреции. При раздражении пищевыми веществами рецепторов слизистой оболочки ротовой полости, глотки и пищевода возникает безусловно-рефлекторное отделение панкреатического сока. Нервные импульсы от раздражаемых рецепторов по афферентным путям достигают бульбарного центра панкреатической секреции, где они переключаются на преганглионарные нейроны ядер блуждающего нерва, по эфферентным волокнам которого достигают постганглионарных нейронов. Их аксоны образуют синаптические окончания на базальных мембранах панкреацитов.
Выделяющийся при возбуждении этих окончаний ацетилхолин активирует М-холинорецепторы постсинаптической мембраны.
При раздражении чревных нервов, осуществляющих симпатическую иннервацию поджелудочной железы, ее сокоотделение тормозится (за счет активации р-адренорецепторов медиатором норадреналином). Но торможение секреции сопровождается накоплением секреторного материала в панкреацитах.
Поступление пищи в желудок во время приема пищи вызывает раздражение хемо- и механорецепторов желудка.
Поступление в двенадцатиперстную кишку желудочного содержимого вызывает вместе с дуоденопанкреатическим секреторным рефлексом (с рецепторов слизистой оболочки) выделение из эндокринных клеток гастро-интестинальных гормонов, стимулирующих или тормозящих сокоотделение поджелудочной железой.
Роль основных гуморальных регуляторов выполняют секретин и ХЦК, которые вырабатываются S- и ССК-клетками слизистой оболочки двенадцатиперстной кишки. Секретин и ХЦК усиливают влияние друг друга на панкреоциты, особенно на фоне выделения ацетилхолина в синаптических окончаниях холинергических нервных волокон, иннервирующих железу. Важным гуморальным возбудителем панкреатической секреции является гастрин.
Изменится ли амплитула потенциала действия нервной клетки, если на нее воздейсвовать стимулами сверхпороговой силы
Билет № 5
1.потенциал действия,с графиком, возбудимость,мера возбудимости,
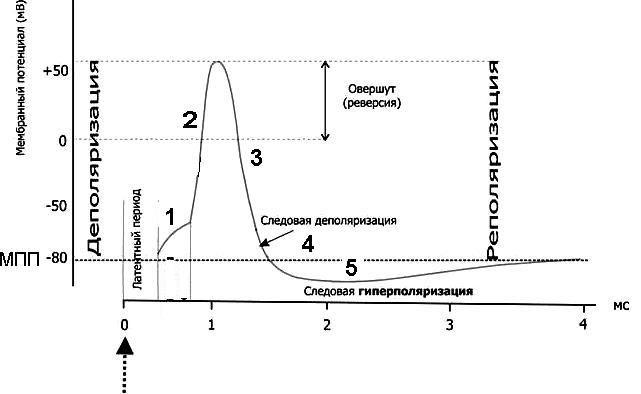
Потенциал действия (ПД) – быстрое изменение мембранного потенциала в ответ на действия раздражителя пороговой силы. ПД имеет стандартные амплитуду и временные параметры, не зависящие от силы стимула - правило "ВСЕ ИЛИ НИЧЕГО".
Начальная деполяризация мембраны под действием раздражителя:
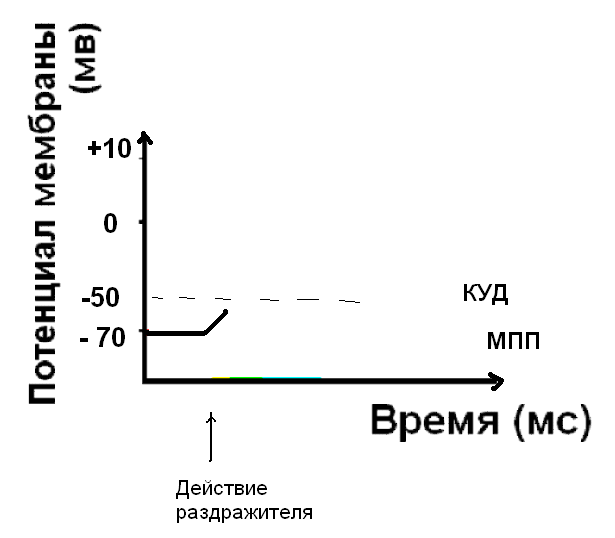
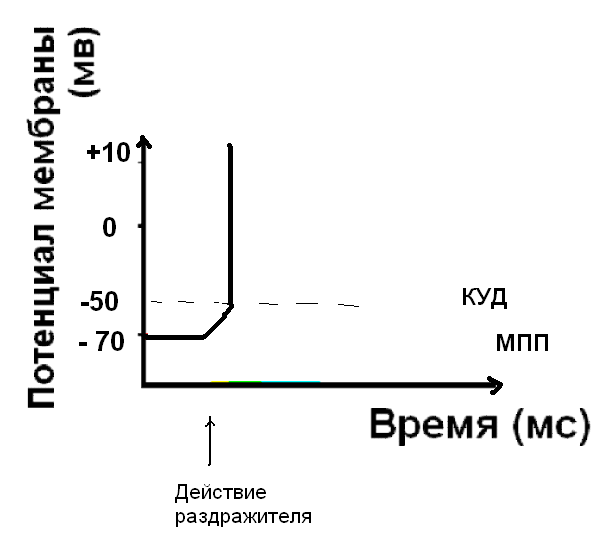
Если сила раздражителя достаточна, чтобы деполяризовать мембрану до КУД, открываются быстрые потенциал-зависимые натриевые каналы. Клетка возбуждена – возник нервный импульс.
Восстановление мембранного потенциала покоя - реполяризация мембраны.
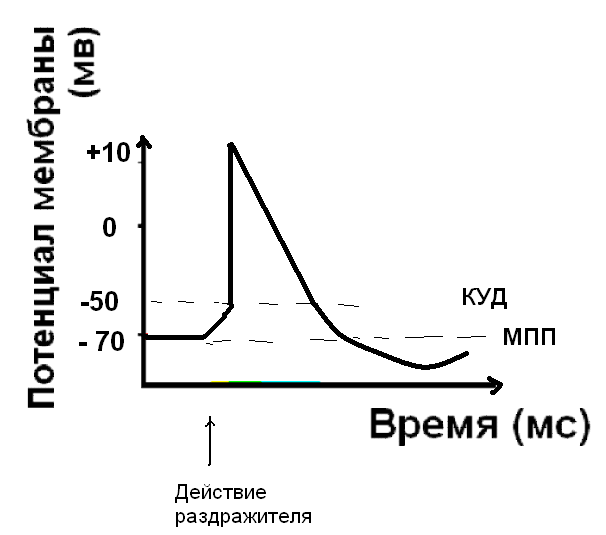
Следующий этап – восстановление мембранного потенциала покоя - реполяризация, обусловлена активным ионным транспортом. Наиболее важен процесс активного транспорта - это работа Na/K - насоса, который выкачивает ионы натрия из клетки, одновременно закачивая ионы калия внутрь клетки. Восстановление мембранного потенциала происходит благодаря току ионов калия из клетки – калиевые каналы активируются и пропускают ионы калия до достижения равновесного калиевого потенциала. Это процесс важен потому, что до тех пор, пока не восстановлен МПП, клетка не способна воспринимать новый импульс возбуждения.
ГИПЕРПОЛЯРИЗАЦИЯ - кратковременное увеличение МП после его восстановления, которое обусловлено повышением проницаемости мембраны для ионов калия и хлора. Гиперполяризация бывает только после ПД.
- Деполяризация мембраны до КУД – могут открыться любые натриевые каналы, иногда кальциевые, и быстрые, и медленные, и потенциал-зависимые, и рецептор-управляемые. Это зависит от вида раздражителя и типа клеток
- Быстрое поступление натрия в клетку - открываются быстрые, потенциал-зависимые натриевые каналы, и деполяризация достигает точки реверса потенциала – происходит перезарядка мембраны, знак заряда меняется на положительный.
- Восстановление градиента концентрации по калию – работа насоса. Калиевые каналы активированы, калий переходит из клетки во внеклеточную среду – реполяризация, начинается восстановление МПП
- Следовая деполяризация, или отрицательный следовой потенциал - мембрана еще деполяризована относительно МПП.
- Следовая гиперполяризация. Калиевые каналы остаются открытыми и дополнительный ток калия гиперполяризует мембрану. После этого клетка возвращается к исходному уровню МПП. Длительность ПД составляет для разных клеток от 1 до 3-4 мс.
Обратите внимание на три величины потенциала, важные и постоянные для каждой клетки ее электрические характеристики.
- МПП - электроотрицательность мембраны клетки в покое, обеспечивающая способность к возбуждению - возбудимость.
- КУД - критический уровень, величина мембранного потенциала, при достижении которой открываются быстрые, потенциал зависимые натриевые каналы и происходит перезарядка мембраны за счет поступления в клетку положительных ионов натрия. Чем выше электроотрицательность мембраны, тем труднее деполяризовать ее до КУД, тем менее возбудима такая клетка.
- Точка реверса потенциала (овершут) - такая величина положительного мембранного потенциала, при которой положительно заряженные ионы уже не проникают в клетку - кратковременный равновесный натриевый потенциал.
При действии раздражителя подпороговой силы возникает неполная деполяризация - ЛОКАЛЬНЫЙ ОТВЕТ (ЛО). Неполная, или частичная деполяризация – это такое изменение заряда мембраны, которое не достигает критического уровня деполяризации (КУД).
Секреция ПЖ находится под контролем нервных и гуморальных механизмов. Начальную секрецию ПЖ вызывают вид, запах пищи и другие раздражители (условнорефлекторные сигналы), а также жевание и глотание (безусловнорефлекторные сигналы). При этом нервные сигналы, формирующиеся в рецепторах полости рта и глотки, достигают продолговатого мозга, и затем эфферентные влияния по волокнам блуждающего нерва поступают к железе и вызывают её секрецию.
У человека с фистулой ГПП выделение панкреатического сока начинается через 2—3 мин после того, как он увидел пищу или услышат о ней. Это пример условнорефлекторного пути возбуждения панкреатической секреции. Симпатические волокна, иннервирующие ПЖ, тормозят её секреторную активность и в то же время изменяют реактивность железы по отношению к другим воздействиям, усиливая синтез органических веществ.
Торможение панкреатической секреции происходит при раздражении многих чувствительных нервов, при болевых реакциях, во время сна, при напряжённой физической и умственной работе.
Для стимуляции панкреатической секреции прямые нервные влияния имеют меньшее значение, нежели гуморальные. Ведущее значение в гуморальной регуляции секреции ПЖ принадлежит желудочно-кишечным гормонам.
Большинство регуляторов секреции ферментов ПЖ действуют на рецепторы мембраны ацинарных клеток, расположенные на их базолатеральной поверхности. Выделяют рецепторы ХК, бомбезина, ацетилхолина, субстанции Р, ВИП, секретина.
Стимуляторы панкреатической секреции. ВИП и секретин стимулируют панкреатическую секрецию, активируя аденилатциклазу. Как и в других типах клеток, аденилатциклаза способствует образованию циклического аденозинмонофосфата, в результате чего протеинкиназа А, усиливающая секрецию панкреатического сока, богатого бикарбонатами, переходит в активную форму. Другие стимуляторы поджелудочной секреции (ХК, ацетилхолин, гастрин-рилизинг пептид, субстанция Р) действуют на специфические рецепторы, во внутриклеточной передаче сигнала от которых задействованы альтернативные вторичные мессенджеры.
Эти вещества повышают внутриклеточное содержание циклического гуанозинмонофосфата, что приводит к увеличению внутриклеточного содержания инозитолтрифосфата, диацилглицерола, арахидоновой кислоты и кальция. Эти промежуточные вещества-посредники активируют различные протеинкиназы, в результате этого повышается секреция ферментов. Данные, полученные в опытах на животных, свидетельствуют, что действие комбинации агонистов на различные мембранные рецепторы может вызывать синергический, но не суммарный (аддитивный) эффект. Например, ХК увеличивает секрецию бикарбонатов, стимулированную секретином, но секретин не повышает секреторный ответ на действие ХК.
Ингибиторы секреции поджелудочной железы. Различные вещества, ответственные за подавление панкреатической секреции, действуют по принципу отрицательной обратной связи во время и после приёма пищи.
Панкреатический полипептид (ПП) представляет собой пептидный гормон, образующийся в островках Лангерганса и подавляющий панкреатическую секрецию воды, бикарбонатов и ферментов. Концентрация ПП в плазме крови возрастает после мнимого кормления, после приёма пищи, после экспериментального закисления среды ДПК, а также при стимуляции блуждающего нерва, при действии ХК, секретина, ВИП. ПП может выступать как антагонист мускариновых рецепторов и способен ингибировать выделение ацетилхолина из постганглионарных нейронов ПЖ; его конечный эффект проявляется на уровне ацинарных клеток.
Пептид YY высвобождается в дистальной части подвздошной кишки и и толстой кишке в ответ на поступление пищи смешанного характера, по жиры, находящиеся в просвете кишки, в большей степени способны стимулировать его секрецию. Пептид YY уменьшает чувствительность ПЖ к действию секретина и ХК, возможно, за счёт уменьшения секреции ацетилхолина и норадреналина и ингибирования выделения ХК слизистой оболочкой ДПК.
Соматостатин ингибирует секрецию секретина слизистой оболочкой ДПК, а также уменьшает чувствительность к секретину рецепторных полей. Единственный эффект соматостатина — снижение секреции ферментов и бикарбонатов ПЖ. Соматостатин синтезируют клетки слизистой оболочки желудка и кишечника, а также D-клетки островков Лангерганса. Его активность составляет только около 25% от активности гипоталамического соматостатина. Но только соматостатин, продуцируемый слизистой оболочкой тонкой кишки, оказывает угнетающее действие на секрецию ПЖ.
Выделение соматостатина происходит при участии автономной нервной системы в ответ на поступление жиров и аминокислот с пищей.
Соматостатин блокирует панкреатическую секрецию несколькими способами. Во-первых, он действует путём угнетения продукции стимулирующих пептидов (ХК). Во-вторых, посредством угнетающего воздействия на ЦНС и регулируя работу интрапанкреатических ганглиев, соматостатин ингибирует поступление ацетилхолина в пресинаптическую щель и, возможно, ингибирует выработку инсулина. Хотя рецепторы к соматостатину найдены также в ацинарных клетках, эти рецепторы скорее способствуют увеличению секреции, чем блокируют её.
Другие ингибиторы, представленные среди гормонов эндокринных клеток островков Лангерганса, включают панкреатический глюкагон и панкреастатин, а также нейропептиды: кальцнтонин-генерирующий пептид и энкефалины (табл. 1-3). Панкреатический глюкагон ингибирует секрецию ПЖ, стимулированную ХК, секретином или пищей; угнетает секрецию бикарбонатов, воды и ферментов. Панкреастатин ингибирует панкреатическую секрецию, тормозя высвобождение ацетилхолина эфферентными окончаниями блуждающего нерва. Кальцитонин-генерирующий пептид может проявлять свою активность через стимуляцию выделения соматостатина. Энкефалины и подобные им опиоиды снижают выделение секретина слизистой оболочкой ДПК и могут также ингибировать высвобождение ацетилхолина.
Пища, попавшая в двенадцатиперстную кишку подвергается воздействию поджелудочного, кишечного соков и желчи. Поджелудочный сок вырабатывается экзокринными клетками поджелудочной железы. Это бесцветная жидкость щелочной реакции. рН=7,4-8,4. В течение суток выделяется 1,5-2,0 л сока. В состав сока входит 98,7% воды и 1,3% сухого остатка.
Сухой остаток содержит:1. Минеральные вещества. Катионы натрия, калия, кальция, магния. Гидрокаробонат, фосфат, сульфат анионы, анионы хлора. Из минеральных веществ преобладает гидрокарбонат натрия. Его 1% из 1,3% сухого остатка. Он определяет щелочную реакцию сока. Благодаря ей кислый химус желудка приобретает нейтральную или даже слабощелочную реакцию. Это создает оптимальную среду для действия панкреатических и кишечных ферментов с рН=7-8.2. Простые органические вещества: мочевина, мочевая кислота, креатинин, глюкоза.3. Ферменты. Они играют важнейшую роль в переваривании белков, жиров и углеводов и делятся на следующие группы:а. Пептидазы. К ним относятся такие эндопептидазы, как трипсин, химотрипсин и эластаза. Они расщепляют внутренние связи белков с образованием поли- и олигопептидов. Экзопептидазами являются карбоксипептидазы А и В. Они отщепляют конечные аминокислотные цепи с образованием ди- и трипептидов и аминокислот. Все эти протеолитические ферменты выделяются железой в неактивной форме в виде трипсиногена, химотрипсиногена, и прокарбоксипетидаз. При поступлении сока в 12-перстную кишку, трипсиноген подвергается воздействию фермента энтерокиназы. От него отщепляется белок ингибитор и трипсиноген переходит в активный трипсин. Этот первоначально образовавшийся трипсин в дальнейшем осуществляет активацию остального трипсиногена и других проферментов поджелудочного сока. Ингибитор трипсина образуется в тех же железистых клетках, что и трипсин. Это предупреждает воздействие пептидаз на клетки железы.б. Липазы. Ими являются панкреатическая липаза и фосфолипаза А. Липаза расщепляет нейтральные жиры до жирных кислот и глицерина, а фосфолипаза - фосфолипиды.в. Карбогидразы. Это а-амилаза сока, которая расщепляет крахмал до мальтозы.г. Нуклеазы. ДНК-аза и РНК-аза. Они гидролизуют нуклеиновые кислоты до нуклеотидов.
Рефлекторная и гуморальная регуляция секреции панкреатического сока (И.П.Павлов, У.Бейлис, Э.Старлинг).
Секреторные клетки поджелудочной железы вне периода пищеварения находятся в состоянии покоя и отделяют сок лишь в связи с периодической деятельностью желудочно-кишечного тракта. Секреция поджелудочной железы происходит иод воздействием нервных влияний и гуморальных раздражителей, возникающих при поступлении пищи в пищеварительный тракт, а также при виде, запахе пищи и в случае действия привычной обстановки ее приема. Как и в случае желудочной секреции, процесс отделения поджелудочного сока разделяется на три фазы: сложнорефлекторную (мозговую или цефалическую), желудочную и кишечную.
Сложнорефлекторная фаза секреции начинается не только при непосредственном воздействии элементов пиши на рецепторы ротовой полости, но и при виде, запахе пищи и при действии той обстановки, в которой происходит акт еды. Поступление пищи в полость рта и глотки вызывает рефлекторное возбуждение, наслаивающееся на уже начавшуюся секрецию поджелудочной железы.. Дуга рефлекса включает рецепторы полости рта, чувствительные нервные волокна, идущие в продолговатый мозг, центральные парасимпатические нейроны, эфферентные волокна вагуса, секреторные клетки поджелудочной железы. Эта фаза значительно менее выражена, нежели в предыдущих отделах пищеварительного тракта.
Желудочная фаза секреции является следствием поступления пищи в желудок. Возбуждение поджелудочной железы возникает при механическом, химическом и гуморальном раздражении рецепторов желудка. Афферентные импульсы, возникающие в результате раздражения хеморецепторов слизистой оболочки желудка, по чувствительным нервным волокнам поступают в центральную нервную систему, откуда по блуждающему нерву эфферентные влияния направляются к поджелудочной железе. Химическими раздражителями являются как вещества, вырабатывающиеся в самом желудочно-кишечном тракте, так и содержащиеся в пище. Натуральными раздражителями, вызывающими возбуждение поджелудочной железы, являются НС1, овощные соки, жиры и продукты их гидролиза. Гуморальным регулятором поджелудочной железы в этой фазе является гормон антраль-ного отдела желудка гастрин. Последний, всасываясь в кровь, возбуждает секрецию поджелудочной железы.
Кишечная фаза секреции начинается после поступления химуса в двенадцатиперстную кишку. В это время вырабатывается большая часть панкреатического сока. Количество и состав секрета поджелудочной железы зависит от качества и количества пищи, контролируется рецептивными клетками кишечника и, в первую очередь, двенадцатиперстной кишки. Эта фаза секреции развивается под влиянием рефлекторных влияний и кишечных гормонов. Доказано существование мощных дуоденопанкреатических рефлексов. Общность иннервации поджелудочной железы, двенадцатиперстной кишки и печени с желчными ходами обусловливает их функциональную взаимосвязь.
Поступление хлористоводородной кислоты и продуктов переваривания пищи в верхний отдел тонкой кишки стимулирует секрецию поджелудочной железы.Панкреатическую секрецию стимулируют также вазоактивный интестинальный полипептид (ВИП), серотонин, образующийся в энтерохромафинных клетках слизистой желудочно-кишечного тракта и ткани поджелудочной железы, инсулин, бомбезин, субстанция Р, соли желчных кислот.
Тормозное влияние на секрецию оказывают глюкагон, кальцитонин, ПП, соматостатин, ГИП. Эффекты интестинальных пептидов опосредуются путем их влияния на секреторную активность желудочных желез, что сопровождается увеличением кислотности желудочного химуса, который при поступлении в двенадцатиперстную кишку стимулирует выделение ее гормонов.
Поперечные профили набережных и береговой полосы: На городских территориях берегоукрепление проектируют с учетом технических и экономических требований, но особое значение придают эстетическим.
Механическое удерживание земляных масс: Механическое удерживание земляных масс на склоне обеспечивают контрфорсными сооружениями различных конструкций.


Опора деревянной одностоечной и способы укрепление угловых опор: Опоры ВЛ - конструкции, предназначенные для поддерживания проводов на необходимой высоте над землей, водой.











- Валеология
- Выбор программы упражнений и ее реализация

Поджелудочная железа
Поджелудочная железа — вторая по величине железа пищеварительной системы, ее масса 60-100 г, длина 15-22 см.
Железа имеет серовато-красный цвет, дольчатая, простирается в поперечном направлении от 12-перстной кишки до селезенки. Ее широкая головка располагается внутри подковы, образованной 12-перстной кишкой. Железа покрыта тонкой соединительной капсулой.
Поджелудочная железа, по существу, состоит из двух желез: экзокринной и эндокринной. Экзокринная часть железы вырабатывает у человека в течение суток 500-700 мл панкреатического сока, который содержит ферменты, участвующие в переваривании белков, жиров и углеводов. Эндокринная часть поджелудочной железы продуцирует гормоны, регулирующие углеводный и жировой обмен (инсулины, глюкагон, соматостатин и др.).
Экзокринная часть поджелудочной железы представляет собой сложную альвеолярно-трубчатую железу, разделенную на дольки очень тонкими соединительными междольковыми перегородками, отходящими от капсулы. В дольках тесно лежат ацинусы, образованные ациноцитами (клетки поджелудочной железы). Клетки тесно соприкасаются друг с другом.
Ацинус со вставочным протоком является структурно-функциональной единицей экзокринной части поджелудочной железы. Секрет поступает в просвет ацинуса. Из вставочных протоков секрет поступает во внутридольковые протоки. Окруженные рыхлой соединительной тканью внутридольковые протоки впадают в междольковые, которые впадают в главный проток поджелудочной железы и, соединяясь с общим желчным протоком, впадают в просвет 12-перстной кишки.
Поджелудочная железа человека натощак выделяет небольшое количество секрета. При поступлении пищевого содержимого из желудка в 12-перстную кишку поджелудочная железа человека выделяет сок со средней скоростью 4,7 мл/мин. За сутки выделяется 1,5-2,5 л сока сложного состава.
Сок представляет собой бесцветную прозрачную жидкость со средним содержанием воды 987 г/л. Сок поджелудочной железы щелочной реакции (рН = 7,5-8,8). Сок поджелудочной железы участвует в нейтрализации и ощелачивании кислого пищевого содержимого желудка в 12-перстной кишке, богат ферментами, которые переваривают все виды питательных веществ.
Таблица. Основные компоненты секрета поджелудочной железы
Показатели
Характеристика
Удельный вес, г/мл
НСО - 3 — до 150 ммоль/л, а также Са 2+ , Mg 2+ , Zn 2+ , НРО4 2- , SO4 2-
Трипсин, химотрипсин, карбоксипептидаза А и В, эластаза
Липаза, фосфолипаза, холестеролипаза, лецитиназа
Секреция сока поджелудочной железы резко усиливается через 2-3 мин после приема пищи и продолжается 6-14 ч. От количества и качества пищи зависит объем, состав выделяемого сока, динамика выделения. Чем выше кислотность пищевого содержимого желудка, поступающего в 12-перстную кишку, тем больше выделяется сока поджелудочной железы.
Секреция поджелудочной железы при стимуляции ее приемом пищи имеет характерную динамику и проходит несколько фаз.

Первая, или мозговая, фаза секреции обусловлена видом, запахом пищи и другими раздражителями, связанными с приемом пищи (условно-рефлекторные раздражения), а также воздействиями на рецепторы слизистой оболочки рта, жеванием и глотанием (безусловно-рефлекторные раздражения). Нервные импульсы, возникающие в рецепторах, достигают продолговатого мозга и затем по волокнам блуждающего нерва поступают к железе и вызывают ее секрецию.
Вторая, или поджелудочковая, фаза характеризуется тем, что секреция железы стимулируется и поддерживается рефлексами с механо- и хеморецепторов желудка.
С переходом желудочного содержимого в 12-перстную кишку начинается третья, или кишечная, фаза секреции, связанная с действием на слизистую оболочку 12-перстной кишки кислого ее содержимого. Механизм секреции направлен на срочную адаптацию секреции ферментов поджелудочной железы к виду принятой пищи. Прием пищи вызывает увеличение выделения всех ферментов в составе сока, но для разных видов пищи это увеличение выражено в разной мере. Пища с большим содержанием углеводов вызывает увеличение в соке амилазов (ферментов, расщепляющих углеводы), белков — трипсина и тринсиногена, жирной пищи — липазы, т.е. поджелудочная железа синтезирует и секретизирует больше того фермента, который гидролизует преобладающие в рационе питательные вещества.
Пищеварение в тонкой кишке
Пищеварение в тонком кишечнике (двенадцатиперстной, тощей и подвздошной кишках) обеспечивает гидролиз большинства компонентов пищи до образования мономеров, в форме которых питательные вещества могут всасываться из кишечника в кровь и лимфу. Пищеварение в нем осуществляется под действием ферментов сока поджелудочной железы в полости кишечника (полостное пищеварение) и под действием ферментов, фиксированных на микроворсинках и нитях гликокаликса (пристеночное пищеварение). Часть этих ферментов продуцируется поджелудочной железой, а часть железами кишечной стенки. Заключительной стадией пищеварения в тонком кишечнике является пищеварение на мембранах эпителиальных клеток кишечника (мембранное пищеварение), осуществляемое под действием ферментов желез кишечной стенки и сопряженное с процессами всасывания питательных веществ.
Основная роль в переваривании пищи в тонком кишечнике принадлежит процессам, происходящим в двенадцатиперстной кишке. Кислый химус, поступивший в нее из желудка, представлен остатками механически обработанной и частично переваренной пищи. В его составе находятся непереваренные жиры в виде триглицеридов, эфиров холестерола, фосфолипидов; частично переваренные до полипептидов и олигопептидов белки; частично переваренные и непереваренные углеводы в виде крахмала, гликогена, клетчатки, а также нуклеиновые кислоты и другие органические и неорганические вещества. Таким образом, для их переваривания пищеварительные железы должны продуцировать большой набор разнообразных ферментов и в кишечнике должны быть созданы оптимальные условия для проявления их активности.
Создание таких условий начинается с того, что химус постепенно нейтрализуется бикарбонатами соков поджелудочной железы, кишки и желчью. Действие пепсина в двенадцатиперстной кишке прекращается, так как рН ее содержимого смещается в сторону щелочной среды, достигая 8,5 (колеблется от 4 до 8,5). Бикарбонаты, другие вещества неорганической природы и вода выделяются в состав сока поджелудочной железы эпителиальными клетками канальцев и протоков железы. Выделение бикарбонатов зависит от рН содержимого кишки и чем выше его кислотность, тем больше выделяется щелочных продуктов, замедляется эвакуация химуса в тощую кишку.
Ферменты сока поджелудочной железы образуются эпителием ацинусов железы. Их образование зависит от характера принимаемой пищи и действия разнообразных регуляторных механизмов.
Сок поджелудочной железы является бесцветной прозрачной жидкостью, содержит много бикарбонатов, а также хлоридов, солей калия, натрия, кальция, магния, небольшое количество сульфатов и фосфатов. В соке много белков, 90% которых — ферменты, расщепляющие белки, жиры и углеводы. За сутки выделяется 1,5-2,5 л сока. Содержание электролитов в соке поджелудочной железы, так же как в слюне и желудочном соке, зависит от скорости его секреции. При увеличении скорости повышается содержание NaHCO3, при снижении возрастает концентрация NaCI.
Основные протеолитические ферменты панкреатического сока секретируются в виде зимогенов, т.е. в неактивном состоянии. Это трипсиноген, химотрипсиноген, проэластаза, прокарбоксипептидазы А и В. Физиологическим активатором трипсиногена и превращения его в трипсин является энтерокиназа (эндопептидаза), вырабатываемая слизистой двенадцатиперстной кишки. Последующее образование трипсина идет аутокаталитически. Трипсин активирует образование из неактивных форм химотрипсина, эластазы, карбоксипептидаз А и В, а также процесс высвобождения энтерокиназы. Трипсин, химотрипсин и эластаза являются эндопептидазами. Они расщепляют белки и высокомолекулярные полипептиды до низкомолекулярных пептидов и аминокислот. Карбоксипептидазы А и В (экзопептидазы) расщепляют пептиды до аминокислот.
Таблица. Гидролитическое действие ферментов поджелудочной железы
Фермент
Участок гидролиза
Протеолитические
Внутренние пептидные связи между соседними аминокислотными остатками

Поджелудочный сок выделяется только во время пищеварения. Когда пищеварение кончается, прекращается и выделение сока. Поджелудочный сок начинает выделяться через 2— 3 минуты после начала приема пищи.
Количество и свойство поджелудочного сока меняются в зависимости от пищи. На разные пищевые вещества поджелудочный сок, как и желудочный сок, выделяется в разных
количествах и с разным содержанием ферментов.
Рис. ОТДЕЛЕНИЕ ПОДЖЕЛУДОЧНОГО СОКА У СОБАКИ ПРИ КОРМЛЕНИИ МЯСОМ, ХЛЕБОМ И МОЛОКОМ.
Кривые выделения поджелудочного сока на мясо, хлеб и молоко, приведенные на рис , почти с точностью совпадают с кривыми выделения желудочного сока на эти же вещества.
Рефлекторное выделение поджелудочного сока
Рефлекторный механизм выделения сока поджелудочной железы был доказан И. П. Павловым в опытах на собаках, в частности в опыте с эзофаготомированной собакой. У такой собаки, помимо перерезки пищевода и желудочной фистулы, была наложена фистула поджелудочной железы. У оперированной таким образом собаки при мнимом кормлении через 2—3 минуты начинается выделение поджелудочного сока. Такой короткий промежуток между раздражением рецепторов полости рта и началом выделения сока говорит о том, что здесь имеет место рефлекторный механизм.
К. М. Быков и его сотрудники проводили наблюдение над человеком с фистулой поджелудочной железы. Выделение поджелудочного сока у него начиналось через 2—3 минуты после разговора о еде. В данном случае имело место условно-рефлекторное сокоотделение.
В эксперименте с предварительной перерезкой блуждающего нерва и последующим перерождением его волокон, тормозящих секрецию поджелудочной железы, И. П. Павлов доказал, что секреторным нервом поджелудочной железы является именно блуждающий нерв, раздражение которого вызывает отделение небольшого количества сока. При еде пища раздражает окончания центростремительных нервов, находящиеся в слизистой оболочке рта и глотки. Возникшее в них возбуждение передается в центр поджелудочного сокоотделения, который находится в продолговатом мозгу, и оттуда по блуж дающему нерву достигает поджелудочной железы; последняя начинает выделять сок.
Нервно-гуморальный механизм выделения поджелудочного сока
При введении в двенадцатиперстную кишку желудочного сока, который содержит соляную кислоту, начинается выделение поджелудочного сока. Поджелудочная железа начинает секретировать и при введении в кишку одной соляной кислоты.
Было высказано предположение, что соляная кислота раздражает нервные окончания и рефлекторно вызывает сокоотделение. Это предположение не оправдалось, так как поджелудочная железа продолжает секретировать и после того, как все нервы, идущие к ней, перерезаны.
Однако соляная кислота вызывает сокоотделение только в том случае, когда ее вводят в двенадцатиперстную кишку. Если соляную кислоту ввести в кровь, минуя кишку, то сокоотделения не наступает; следовательно, не сама соляная кислота вызывает секрецию поджелудочной железы.
Механизм действия соляной кислоты при введении ее В двенадцатиперстную кишку был исследован в специальных опытах. Он состоит в том, что в слизистой оболочке кишки под влиянием соляной кислоты образуется особое вещество — с е к р е т и н. Секретин, всасываясь и поступая в ток крови, кровью доставляется к поджелудочной железе, действует на ее нервно-железистый аппарат и вызывает сокоотделение. Окончательным доказательством явился следующий опыт: у собаки вырезали двенадцатиперстную кишку, положили ее в 0 ,5% раствор соляной кислоты, а через некоторое время этот раствор ввели в кровь другой собаке; тогда поджелудочная
железа второй собаки начала секретировать. В естественных условиях пища, обильно смоченная кислым желудочным соком, поступает в двенадцатиперстную кишку. Соляная кислота желудочного сока действует на слизистую оболочку и вызывает образование в ней из недеятельного просекретина секретина, который затем разносится кровью и возбуждает поджелудочную железу.
Данные, полученные в последнее время, показывают, что секретин действует при участии нервной системы. Это было установлено в опытах, в которых симпатическая нервная система выключалась эрготоксином. Если такому животному в двенадцатиперстную кишку ввести соляную кислоту, то сок из поджелудочной железы почти не будет выделяться. Отсюда следует, что, вероятно, секретин действует на секреторные клетки поджелудочной железы через нервную систему.
Жир, продукты расцеплении которого —мыла — обладают сильным сокогонным действием, также вызывает выделение поджелудочного сока; кроме того, поступление жира в кишечник способствует усиленному образованию секретина в слизистой оболочке кишечника.
Выделение поджелудочного сока усиливает вода, особенно газированная, клюквенный морс и т. д.
Деятельность поджелудочной железы нарушается у людей, желудочный сок которых лишен соляной кислоты или беден ею. Пониженное содержание соляной кислоты в желудочном соке или ее отсутствие называется ахилией. Ввиду того что у таких людей соляная кислота в двенадцатиперствую кишку не поступает, то не происходит и образования секретина и деятельность поджелудочной железы нарушается. Это нарушение отчасти восполняется тем, что вода и некоторые другие вещества способствуют деятельности поджелудочной железы и обеспечивают ее функционирование.
Статья на тему Выделение поджелудочного сока
Читайте также: