
Скорость проведения информации в нервной
Нервная система человека выступает своеобразным координатором в нашем организме. Она передаёт команды от мозга мускулатуре, органам, тканям и обрабатывает сигналы, идущие от них. В качестве своеобразного носителя данных используется нервный импульс. Что он собой представляет? С какой скоростью работает? На эти, а также на ряд других вопросов можно будет найти ответ в этой статье.
Чем является нервный импульс?
Исследование строения и работы
Впервые прохождение нервного импульса было продемонстрировано немецкими учеными Э. Герингом и Г. Гельмгольцем на примере лягушки. Тогда же и было установлено, что биоэлектрический сигнал распространяется с указанной ранее скоростью. Вообще, такое является возможным благодаря особенному построению нервных волокон. В некотором роде они напоминают электрический кабель. Так, если проводить параллели с ним, то проводниками являются аксоны, а изоляторами – их миелиновые оболочки (они являют собой мембрану шванновской клетки, которая намотана в несколько слоев). Причем скорость нервного импульса зависит в первую очередь от диаметра волокон. Вторым по важности считается качество электрической изоляции. Кстати, в качестве материала организмом используется липопротеид миелин, который обладает свойствами диэлектрика. При прочих равных условиях, чем больше будет его слой, тем быстрее будут проходить нервные импульсы. Даже на данный момент нельзя сказать, что эта система полноценно исследована. Многое, что относится к нервам и импульсам, ещё остаётся загадкой и предметом исследования.
Особенности строения и функционирования
Где они создаются?
Типы клеток
- Рецепторные (чувствительные). Ими кодируются и превращаются в нервные импульсы все температурные, химические, звуковые, механические и световые раздражители.
- Вставочные (также называются кондукторными или замыкательными). Они служат для того, чтобы перерабатывать и переключать импульсы. Наибольшее их число находится в головном и спинном мозге человека.
- Эффекторные (двигательные). Они получают команды от центральной нервной системы на то, чтобы были совершены определённые действия (при ярком солнце закрыть рукой глаза и так далее).
Каждый нейрон имеет тело клетки и отросток. Путь нервного импульса по телу начинается именно с последнего. Отростки бывают двух типов:
- Дендриты. На них возложена функция восприятия раздражения расположенных на них рецепторов.
- Аксоны. Благодаря им нервные импульсы передаются от клеток к рабочему органу.
Интересный аспект деятельности
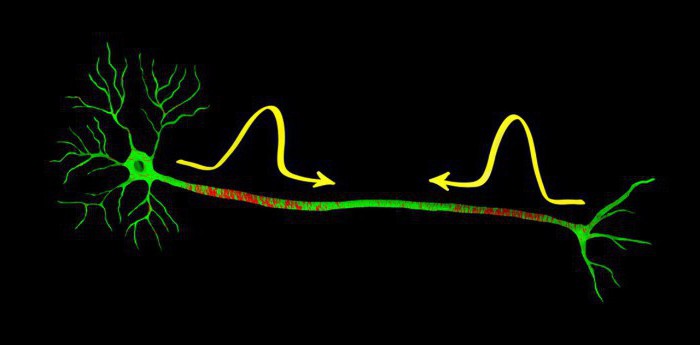
О потенциале действия
Как всё работает в мозгу?
Работа нейромедиаторов
Когда они передают нервные импульсы, то существует несколько вариантов, что произойдёт с ними:
- Они будут диффундированы.
- Подвергнутся химическому расщеплению.
- Вернутся назад в свои пузырьки (это называется обратным захватом).
В конце 20-го века сделали поразительное открытие. Ученые узнали, что лекарства, что влияют на нейромедиаторы (а также их выброс и обратный захват), могут изменять психическое состояние человека коренным образом. Так, к примеру, ряд антидепрессантов вроде "Прозака" блокируют обратный захват серотонина. Есть определённые причины считать, что в болезни Паркинсона виноват дефицит в головном мозге нейромедиатора дофамина.
Если кратко, то они могут работать с тысячами нейромедиаторов, которые посылаются их соседями. Детали относительно обработки и интеграции данного типа импульсов нам почти не известны. Хотя над этим работает много исследовательских групп. На данный момент получилось узнать, что все полученные импульсы интегрируются, а нейрон выносит решение – необходимо ли поддерживать потенциал действия и передавать их дальше. На этом фундаментальном процессе базируется функционирование головного мозга человека. Ну что ж, тогда это неудивительно, что мы не знаем ответа на эту загадку.
Некоторые теоретические особенности
Где же создаются нервные импульсы?
Откуда они начинают свой путь? Ответ на этот вопрос может дать любой студент, который прилежно изучал физиологию возбуждения. Есть четыре варианта:
- Рецепторное окончание дендрита. Если оно есть (что не факт), то возможным является наличие адекватного раздражителя, что создаст сначала генераторный потенциал, а потом уже и нервный импульс. Подобным образом работают болевые рецепторы.
- Мембрана возбуждающего синапса. Как правило, такое возможно только при наличии сильного раздражения или их суммирования.
- Триггерная зона дентрида. В этом случае локальные возбуждающие постсинаптические потенциалы формируются как ответ на раздражитель. Если первый перехват Ранвье миелинизирован, то они на нём суммируются. Благодаря наличию там участка мембраны, которая обладает повышенной чувствительностью, здесь возникает нервный импульс.
- Аксонный холмик. Так называют место, где начинается аксон. Холмик – это наиболее частый создать импульсов на нейроне. Во всех остальных местах, которые рассматривались ранее, их возникновение гораздо менее вероятное. Это происходит из-за того, что здесь мембрана имеет повышенную чувствительность, а также пониженный критический уровень деполяризации. Поэтому, когда начинается суммирование многочисленных возбуждающих постсинаптических потенциалов, то раньше всего на них реагирует холмик.
Пример распространяющегося возбуждения
Вспомните сводки из новостей прошлого лета (также это скоро можно будет услышать опять). Пожар распространяется! При этом деревья и кустарники, которые горят, остаются на своих местах. А вот фронт огня идёт всё дальше от места, где был очаг возгорания. Аналогичным образом работает нервная система.
Часто бывает необходимо успокоить начавшееся возбуждение нервной системы. Но это не так легко сделать, как и в случае с огнем. Для этого совершают искусственное вмешательство в работу нейрона (в лечебных целях) или используют различные физиологические средства. Это можно сравнить с заливанием пожара водой.
Передача информации в нервной системе
Передача информации в нервной системе
Электрические сигналы, обеспечивающие функционирование нервных клеток, опосредованы потоком ионов через водопроницаемые поры клеточной мембраны. Эти поры, образованные трансмембранными белками, называются ионными каналами. К настоящему времени разработаны высокочувствительные методы, позволяющие зарегистрировать и измерить ионные токи, протекающие через одиночные ионные каналы.
Некоторые ионные каналы избирательно проницаемы только для катионов, тогда как другие проводят только анионы. Катионные каналы могут быть высоко избирательными по отношению к одному иону, например натрию. Ионные каналы совершают переходы между открытым и закрытым состоянием и имеют, как правило, характерное время открытого состояния. Их вклад в ионный ток через клеточную мембрану определяется относительным количеством времени, которое они находятся в открытом состоянии.
Открытие канала регулируется различными механизмами. Некоторые из этих механизмов физические, такие как растяжение мембраны или изменения мембранного потенциала. Другие механизмы химические, включающие связывание активных молекул (лигандов) с активным центром, который располагается либо с внеклеточной, либо с внутриклеточной стороны канала.
Важным свойством каналов, в дополнение к кинетике открытия и закрытия, является способность открытого канала проводить ионный ток. Один из способов, которым ионы могут проникать через открытый канал, является простая диффузия. Другой способ – взаимодействие ионов с внутриканальными центрами связывания и перескакивание внутри водной поры от одного центра к другому. В любом случае движение иона через канал является пассивным и определяется градиентом концентрации и градиентом электрического потенциала на мембране.
Количество тока, проходящего через открытый канал по электрическому градиенту, зависит от проницаемости канала для данного типа ионов. Величина тока также зависит от концентрации ионов в устьях канала. Эти два фактора, проницаемость и концентрация, определяют проводимость канала.
Передача импульса в нервной системе опосредуется изменениями мембранного потенциала. В сенсорных нейронах адекватный стимул, такой как прикосновение, звук, свет, вызывает локальную деполяризацию (делая мембранный потенциал менее негативным) или гиперполяризацию (мембранный потенциал становится более негативным). Подобным же образом нейротрансмиттеры в синапсах вызывают деполяризацию или гиперполяризацию постсинаптической клетки. Потенциалы действия, представляющие собой короткие деполяризационные сигналы большой амплитуды, проводят по отросткам нейрона информацию из одного отдела нервной системы в другой.
Все эти изменения мембранного потенциала вызваны движением ионов через клеточную мембрану. Например, направленное внутрь клетки движение положительно заряженных ионов натрия снижает общий отрицательный заряд мембраны или, другими словами, вызывает деполяризацию. Наоборот, результатом движения положительно заряженных ионов калия из клетки является рост общего отрицательного заряда, то есть гиперполяризация. Гиперполяризация может быть обусловлена также движением внутрь клетки отрицательно заряженных ионов хлора.
Как движутся ионы через клеточную мембрану и чем их движение регулируется? Главным путем для быстрого перемещения ионов внутрь клетки и из нее являются ионные каналы. Ионные каналы представляют собой встроенные в мембрану молекулы белка, которые образуют поры, проницаемые для ионов. Ионные токи регулируются через открытие и закрытие этих ионных каналов. Знание механизмов работы ионных каналов позволяет понять, как генерируются электрические сигналы.
Свойства ионных каналов. Клеточная мембрана нервной клетки
Клеточные мембраны состоят из жидкой фазы липидов и встроенных в липиды белковыхмолекул. Молекулы липидов организованы в двухслойную мембрану (бислой) толщиной около 6 нм. Полярные гидрофильные головки липидов обращены к поверхностям мембраны, а гидрофобные хвосты вытянуты к середине бислоя. Липиды плохо пропускают воду и практически непроницаемы для ионов. Белковые молекулы частично погружены в слой липидов, либо с внеклеточной, либо с цитоплазматической стороны. Некоторые белки целиком пронизывают мембрану. Именно пронизывающие мембрану (трансмембранные) белки образуют ионные каналы. Основные ионы, участвующие в генерации электрических сигналов, такие как калий, натрий, кальций или хлор, движутся через ионные каналы пассивно благодаря градиенту концентраций и электрическому потенциалу мембраны.
Другие трансмембранные белки служат в качестве насосов и переносчиков, обеспечивающих транспорт веществ через клеточную мембрану против электрохимических градиентов. Транспортные механизмы поддерживают ионный состав цитоплазмы, удаляя или возвращая те ионы, которые прошли клеточную мембрану по их электрохимическим градиентам. Они также выполняют важную функцию переноса через клеточные мембраны субстратов метаболических реакций, таких как глюкоза и аминокислоты
Для ионных каналов функционально важными являются переходы между открытым и закрытым состояниями. Эти переходы совершаются практически моментально. С другой стороны, при системном изучении поведения любого ионного канала мы обнаружим, что время открытого состояния варьирует случайным образом. Иногда канал открыт только одну миллисекунду или даже меньше, хотя в следующий раз он может быть открыт на гораздо более продолжительное время. Тем не менее, каждый канал имеет характерное среднее время открытого состояния (т), и все вариации происходят вокруг этого среднего показателя.
Некоторые ионные каналы открываются достаточно часто даже в покое. Иными словами, вероятность нахождения таких каналов в открытом состоянии в неактивированной клетке относительно высока. Большинство таких ионных каналов проницаемо для калия или хлора. Они важны для генерации мембранного потенциала покоя. Остальные ионные каналы при этом закрыты, то есть вероятность нахождения их в открытом состоянии очень низка. Активация этих каналов адекватным стимулом резко увеличивает вероятность открытия. Этот же стимул может деактивировать ионные каналы, бывшие активными в покое. Важно помнить, что активация или деактивация канала означает возрастание или снижение вероятности открытия канала, но не увеличение или уменьшение времени открытого состояния (т) канала.
Помимо активации и деактивации, ионный ток через каналы регулируется двумя другими факторами. Первый фактор заключается в том, что ионный канал переходит в новое конформационное состояние, в котором обычный активирующий стимул не способен вызвать открытие канала. Для ионных каналов, активируемых деполяризацией, такое состояние называется инактивацией. Для каналов, отвечающих на химические стимулы, это состояние известно как десенситнзация. Второй механизм – блок открытого канала. Такое случается, когда, например, крупная молекула (такая как токсин) связывается с ионным каналом и физически закупоривает пору. Другим примером может служить блокирование некоторых катионных каналов ионами магния. В этом случае ионы магния сами не проникают через ионный канал, но связываются с каналом в области его устья и тем самым мешают проникновению других катионов.
Некоторые каналы специфически отвечают на физические изменения в клеточной мембране нейрона. Наиболее яркими представителями этой группы являются потенциал-активируемые каналы. Примером может служить чувствительный к потенциалу натриевый канал, который отвечает за регенеративную деполяризацию, лежащую в основе генерации потенциала действия. К этой группе относятся также механочувствительные ионные каналы, которые отвечают на механическое воздействие на клеточную мембрану. Рецепторы растяжения, содержащие ионные каналы такого рода, найдены в механорецепторах кожи.
Другие ионные каналы открываются тогда, когда химические агенты активируют связывающие центры на молекуле канала. Такие лиганд-активируемые ионные каналы подразделяются на две подгруппы, в зависимости от того, являются ли активные центры внутриклеточными или внеклеточными. Каналом, отвечающим на внеклеточную активацию, является катионный канал постсинаптической мембраны в скелетной мышце. Этот канал активируется нейротрансмиттером ацетилхолином, освобождающимся из двигательного нервного окончания. Открытие ацетилхолин-активируемого ионного канала позволяет ионам натрия войти в клетку, вызывая деполяризацию мышечного волокна.
Лиганд-активируемые каналы, отвечающие на внутриклеточные стимулы, включают каналы, чувствительные к местным изменениям концентрации специфических ионов. Например, кальций-активируемые калиевые каналы активируются локальным повышением концентрации внутриклеточного кальция. Такие каналы играют важную роль в реполяризации клеточной мембраны во время завершения потенциала действия. Помимо ионов кальция, типичными представителями лигандов, активирующих ионные каналы с цитоплазматической стороны мембраны, являются циклические нуклеотиды. Циклический ГМФ, например, отвечает за активацию натриевых каналов в палочках сетчатки. Такой тип канала играет принципиальную роль в работе зрительного анализатора.
Эта классификация не является достаточно строгой. Например, кальций-активируемые калиевые каналы чувствительны также к изменению потенциала, а некоторые потенциал-активируемые ионные каналы чувствительны к внутриклеточным лигандам.
Для измерения ионных токов через одиночные каналы первоначально был предложен непрямой метод анализа мембранного шума. Затем был разработан способ прямой регистрации одиночных ионных каналов с помощью метода, который называется пэтч-кламп (patch-clamp). В совокупности эти подходы дали прямые ответы на вопросы, касающиеся функции ионных каналов, как то: какой заряд проходит через одиночный канал? как долго канал остается открытым? как время нахождения ионного канала в открытом или закрытом состоянии зависит от мембранного потенциала?
Таким образом, пэтч-кламп метод предоставляет новые уникальные возможности для изучения поведения ионных каналов. Во-первых, изоляция маленького участка мембраны позволяет наблюдать активность всего нескольких ионных каналов, а не тысяч, которые активируются в целой клетке. Во-вторых, высокое сопротивление контакта дает возможность регистрировать даже крайне одиночных ионных каналов и можем провести анализ кинетики каналов.
До разработки пэтч-кламп метода свойства ионных каналов в клеточных мембранах исследовались в экспериментах, в которых для измерения мембранного потенциала или мембранного тока использовались стеклянные микроэлектроды. Использование Лингом и Джерардом в 1949 году стеклянных микроэлектродов для внутриклеточной регистрации ионных токов в живых клетках было не менее важным событием, чем введение пэтч-кламп метода три десятилетия спустя. Этот метод обеспечивал точное измерение мембранного потенциала покоя клетки, потенциала действия, а также ответов на синаптическую активацию мышечных волокон и нейронов.
Метод внутриклеточной регистрации. Острая стеклянная микропипетка, диаметр кончика которой не превышает 0,5 мкм, заполненная концентрированным солевым раствором (например, 3 M KC1), служит электродом и присоединяется к вольтметру для записи потенциала. Момент прокалывания пипеткой клеточной мембраны, приводящий к проникновению ее в клеточную цитоплазму, проявляется мгновенным появлением потенциала, соответствующего мембранному потенциалу покоя. При удачном проникновении в клетку мембрана обхватывает внешнюю поверхность пипетки, благодаря чему цитоплазма остается изолированной от внеклеточной жидкости.
Используя известную из физики технику анализа шума, Катц и Миледи смогли получить информацию о среднестатистическом поведении отдельного ионного канала, активируемого АХ. Позднее подобные эксперименты были проведены на том же объекте Anderson и Stevens. В отличие от предшественников, эти исследователи измеряли мембранный ток, вызванный АХ, что позволило установить величину и продолжительность ионных токов через одиночный канал.
Принципы анализа шума достаточно просты: во-первых, если токи одиночного канала являются большими, суммарный шум также будет большим. Во-вторых, ионные каналы, открывающиеся на относительно длительное время, будут продуцировать низкочастотный шум; наоборот, каналы, открывающиеся на короткое время, будут продуцировать высокочастотный шум. Исследование амплитудно-временных характеристик шумов, активированных АХ в нервно-мышечном синапсе, показало, что через одиночный открытый ионный канал проходит около 10 миллионов ионов в секунду. Кроме того, выяснилось, что значение среднего открытого времени ионного канала составляет от 1 до 2 мс.
Несмотря на широкое вытеснение пэтч-кламп методом, анализ шума до сих пор используется для изучения ионных каналов в клетках, которые не поддаются исследованию с помощью пэтч-клампа, например, в некоторых областях центральной нервной системы. Кроме того, анализ шума является сравнительно быстрым методом для получения информации о свойствах большой популяции каналов и используется в комбинации с пэтч-кламп регистрацией от целой клетки для идентификации типов каналов. Тем не менее, надо понимать, что с помощью анализа шума невозможно получить детальную информацию о поведении одиночного канала, особенно в каналах со сложной кинетикой или при наличии нескольких уровней проводимости канала.
Кинетическое поведение канала, то есть время его нахождения в закрытом и открытом состояниях, может предоставить информацию о механизмах открытия и закрытия канала, а также о константах скоростей этих процессов. С другой стороны, величина тока, проходящего через ионный канал, является прямым отражением того, как быстро проникающие ионы движутся через канал. Ток ионов зависит не только от свойств канала, но также от трансмембранного потенциала. На этом рисунке изображен фрагмент мембраны, который содержит один спонтанно активный ионный канал, проницаемый для калия. Растворы, как в пипетке, так и в ванночке для объекта, содержат одинаковую (150 ммоль) концентрацию ионов калия. Ионы калия через открытый канал могут двигаться в обоих направлениях. Однако поскольку концентрации ионов по обе стороны мембраны идентичны, а трансмембранный потенциал отсутствует, то нет никакого движения ионов ни в одном Пэтч-кламп метод имеет достоинство, которое еще не было упомянуто: мы можем менять потенциал на регистрирующей пипетке и варьировать, таким образом, трансмембранную разность потенциалов. Например, при мембранном потенциале +20 Мв каждое открытие калиевого ионного канала сопровождается током, направленным наружу. Это связано с тем, что положительно заряженные ионы калия двигаются через канал по электрическому градиенту между раствором в пипетке и в ванночке. С другой стороны, когда внутри пипетки создан отрицательный потенциал величиной в -20 мВток направлен в обратном направлении (через открытый канал в пипетку).
Зависимость тока является линейной: ток (I), проходящий через канал, пропорционален потенциалу (V): 
Это формула представляет собой преобразованный закон Ома. Константа 7 называется проводимостью канала. При одном и том же потенциале на мембране канал с высокой проводимостью переносит много тока, канал с низкой проводимостью проводит малый ток.
Проводимость измеряется в сименсах (См). В нейронах трансмембранный потенциал обычно выражается в милливольтах (1 мВ = 10 –3 В), токи одиночных ионных каналов в пикоамперах (1 пА = 10 –12 А), проводимость в пикосименсах (1 пСм = 10 –12 См). потенциал +20 мВ продуцировал ток около 2,2 пА, соответственно проводимость канала (I/V) составила 2,2 пА/20 мВ = 110 пСм
Электрические сигналы в нервной системе генерируются движением ионов через мембрану нервной клетки. Эти ионные токи протекают через водные поры трансмембранных белков, известных как ионные каналы.
Каналы различаются по своей избирательности: некоторые катионные каналы пропускают только натрий, калий или кальций, другие являются менее избирательными. Анионные каналы сравнительно не избирательны для малых анионов, но они пропускают в основном ионы хлора, так как хлор является самым распространенным анионом внеклеточной и внутриклеточной жидкостей.
Крутько В.Н., Славин М.Б., Смирнова Т.М. Математические основания геронтологии.
Реутов В.П. и др. Проблема оксида азота в биологии и медицине и принцип цикличности.
Методология биологии: новые идеи (синергетика, семиотика, коэволюция). Ред. Баксанский О.Е.
Идею использования простых физиологических показателей для оценки индивидуальных различий по интеллекту предложил выдающийся биолог Ф. Гальтон (цит. по: Ждан, 2004). Он рассматривал интеллект как биологическое образование, которое можно количественно оценить с помощью физиологических индикаторов. Экспериментальное воплощение эти идеи нашли в ряде работ, где в качестве коррелята интеллекта и частично способа его измерения предлагалось рассматривать время выполнения простых заданий. В современной психологии этот подход четко сформулирован П. Верноном: некоторая часть индивидуальных различий в успешности выполнения тестов интеллекта объясняется различиями в том, насколько быстро индивид может обрабатывать информацию; последнее сравнительно независимо от любых приобретенных знаний или навыков. Таким образом, понятие психической скорости или скорости выполнения умственных действий
Гипотезы о связи умственных способностей и переработки информации в ЦНС. В качестве объекта исследования для установления связей с психометрическим интеллектом могут выступать разные показатели этой скорости, характеризующие отдельные этапы или операции деятельности человека. Это может быть время простой реакции, время реакции выбора и время опознания (inspectin time). По некоторым представлениям скорость проведения нервного импульса должна коррелировать с показателями психометрического интеллекта. Считается, что скорость проведения нервного импульса отражает эффективность функционирования нервной системы: чем выше скорость, тем быстрее нервная система обрабатывает информацию. Ускоренная обработка информации в нервной системе рассматривается как физиологическое условие увеличения умственных возможностей индивида (Рийсдик, Бумсма, 2001).
Согласно другой теории, общие способности могут быть связаны с точностью и безошибочностью передачи информации от нейрона к нейрону (Eysenck, 1982). Иначе говоря, предполагается, что индивиды, нейроны которых быстро проводят информацию вдоль аксона, во-первых, передают информацию эффективно и точно через синапсы, во-вторых, будут более успешно обрабатывать информацию по сравнению с теми, у кого скорость распространения нервных импульсов меньше и при передаче информации происходит больше искажений.
В этом контексте была сформулирована так называемая миели-новая гипотеза (Miller, 1994), согласно которой более высокий интеллект связан с большими размерами мозга, которые, в свою очередь, зависят от толщины миелиновой оболочки. Толстая миелино-вая оболочка, во-первых, обеспечивает более высокую скорость проведения импульса, во-вторых, препятствует вмешательству в этот процесс посторонних сигналов. Тем самым она повышает надежность передачи информации. Подтверждают эту гипотезу положительные корреляции между размером мозга и показателями интеллекта, которые были получены при применении метода ядерно-маг-нитного резонанса.
Время как фактор эффективности. Как уже отмечалось, скорость выполнения умственных действий приобретает роль фактора, объясняющего происхождение индивидуальных различий в познавательной деятельности и показателях интеллекта. Показано, что показатель интеллекта связан с временем реакции, взятом в разных вариантах оценки, отрицательной корреляцией, составляющей в среднем -0,3 (Deary et al., 2010). Помимо этого, хронометрию процессов переработки информации можно провести, используя латентности компонентов ВП, интерпретируемые как маркеры времени выполнения отдельных когнитивных операций. Закономерно, что существует целый ряд исследований взаимосвязи показателей ВП и интеллекта.
Для сопоставления с показателями IQ привлекаются и другие параметры ВП: различные варианты амплитудных оценок, вариативность, асимметрия. Наибольшую известность в связи с этим приобрели исследования взаимосвязи некоторых особенностей ВП, синаптической передачи и формирования энграмм памяти (Hendrickson, 1982; пит. по: Айзенк, 1995). Предполагалось, что при обработке информации на уровне синапсов в коре мозга могут возникать ошибки. Чем большее число таких ошибок продуцирует индивид, тем ниже показатели его интеллекта. Количественно оценить число этих ошибок невозможно, но они проявляются в индивидуальных особенностях конфигурации ВП. Согласно этой концепции индивиды, безошибочно обрабатывающие информацию, должны продуцировать высокоамплитудные и имеющие сложную форму ВП, т.е. с дополнительными пиками и колебаниями (рис. 17.1).
ВП упрощенной формы характерны для индивидов с низкими показателями интеллекта. Эти предположения получили статистическое подтверждение при сопоставлении ВП и показателей интеллекта по тестам Векслера и Равена. Таким образом, есть основания утверждать, что эффективность передачи информации на нейронном уровне определяется двумя параметрами: скоростью и точностью (безошибочностью). Оба параметра можно рассматривать как характеристики биологического интеллекта.
Не только вызванные потенциалы, но и индивидуальные особенности ЭЭГ дают основания для прогноза интеллекта (Разумникова, 2009). Это также подтверждают работы А.Н. Лебедева с соавторами (1997). С использованием статистических средств обработки данных ими было показано, что интеллектуальные способности индивида можно прогнозировать по таким показателям ЭЭГ, как частота альфа-ритма, степень синхронизации колебаний в лобных областях, соотношение мощности колебаний в альфа- и тета-диапазонах.


Рис. 17.1. Вызванные потенциалы у шести испытуемых с высокими (слева) и шести испытуемых с низкими (справа) показателями IQ
(по Eysenck, 1995)
что он ограничивается меньшим числом локусов активации по сравнению с менее успешными испытуемыми. Не исключено, что причина возникшего противоречия в степени сложности заданий. По мере усложнения заданий число локусов активации у индивидуумов с более высоким IQ может увеличиваться (Deary et al., 2010).
Топографические факторы. Роль топографических факторов в обеспечении мышления и интеллекта можно рассматривать по крайней мере в двух аспектах. Первый зависит от морфологических и функциональных особенностей отдельных структур мозга, которые связаны с высокими умственными достижениями. Второй касается особенностей взаимодействия между структурами мозга, при которых возможна высокоэффективная умственная деятельность.
Мышление и интеллект представляют собой свойство мозга как целого, поэтому особое значение приобретает анализ взаимодействия различных регионов мозга, при котором достигается высокоэффективная умственная деятельность. В этом контексте представляет интерес модель теменно-лобной интеграции (P-FIT model), объясняющая корковый механизм решения проблемы. Р. Юнг и Р. Хайер (Jung, Haier, 2007) провели метаанализ 37 эмпирических исследований, выполненных разными вариантами нейровизуализации. Теоретически они исходили из того, что человек собирает и обрабатывает информацию, объединяя данные зрения и слуха. Поэтому особенно важна ранняя сенсорная обработка в экстра-стриарной коре (поля 18 и 19) и веретеновидной извилине (поле 37), отвечающих за формирование визуального опыта, а также зона Вернике (поле 22), отвечающая за смысловую интерпретацию акустических стимулов. Обработанная сенсорная информация направляется в теменную кору, а точнее — в супрамаргинальную извилину (поле 40), верхнюю теменную (поле 7), ангулярную извилину (поле 39) (рис. 17.2). В этих центрах осуществляется смысловая обработка информации, включающая символизацию, абстракцию и уточнение проблемы. Далее теменная кора взаимодействует с лобными областями (поля 6, 9, 10, 45—47). Эти зоны обеспечивают выбор подходящего решения проблемы. Если решение найдено, оно поступает в переднюю поясную кору (поле 32), которая ограничивает возможность дальнейшего выбора и тормозит конкурирующие ответы. Авторы подчеркивают: конечный результат зависит от точности функционирования дугообразного пучка передающих информацию нервных волокон.

Рис. 17.2. Корковые поля, обозначенные по карте Бродмана, связанные с высокими показателями выполнения тестов интеллекта и мышления. Эти поля образуют теменно-лобную интеграционную систему; темные кружки — доминирующие центры левого полушария; серые кружки — доминирующие двусторонние центры; белая стрелка — дугообразный пучок, связывающий центры
Брока и Вернике (по Jung, Haier, 2007)
Межполушарные отношения. Проблема специализации полушарий в познавательной деятельности человека хорошо изучена (см. п. 5.4 и 8.5). Аналитическая, знаково опосредованная стратегия познания характерна для работы левого полушария, синтетическая, образно опосредованная — правого. Закономерно, что функциональные свойства полушарий, а точнее, степень их индивидуальной выраженности, могут служить физиологическим условием высоких достижений в решении задач разного типа (вербально-логических или пространственных).
Исходно предполагалось, что условием высоких достижений в умственной деятельности является развитие функций доминантного левого полушария, однако в настоящее время все большее значение в этом плане придается функциям субдоминантного правого полушария. Возникла гипотеза эффективного билатерального взаимодействия как физиологической основы общей одаренности. Чем лучше праворукий человек использует возможности своего субдоминантного правого полушария, тем больше он способен: одновременно обдумывать разные вопросы; привлекать больше ресурсов для решения интересующей его проблемы; одновременно сравнивать свойства объектов, вычленяемые познавательными стратегиями каждого из полушарий. Гипотеза билатерального взаимодействия и эффективного использования всех возможностей левого и правого полушарий в интеллектуальной деятельности представляется оптимальной, поскольку она, во-первых, адресуется к работе мозга как целого и, во-вторых, использует представления о ресурсах мозга.
Читайте также: