
Строение ганглиев у животных
Центральная нервная система человека осуществляет контроль над деятельностью его организма и разделяется на несколько отделов. Головной мозг посылает и получает сигналы из организма и после их обработки имеет информацию о процессах. Нервная система разделяется на вегетативную и соматическую нервную системы.
Отличия вегетативной и соматической нервной системы
Соматическая нервная система регулируется сознанием человека и может управлять деятельностью скелетной мускулатуры. Все компоненты реакции человека на внешние факторы находятся под контролем полушарий мозга. Она обеспечивает сенсорные и моторные реакции человека, контролируя их возбуждение и торможение.
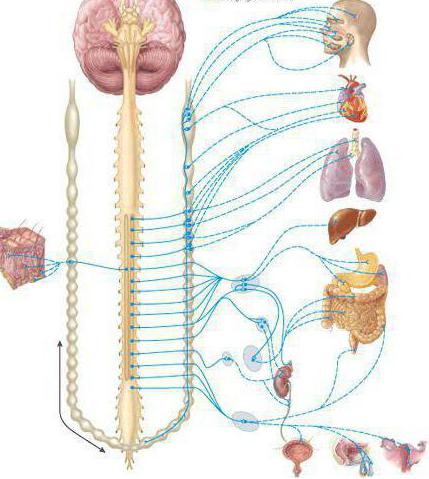
Вегетативная нервная система контролирует периферическую деятельность организма и не контролируется сознанием. Для нее характерны автономность и генерализованность воздействия на организм при полном отсутствии сознания. Эфферентная иннервация внутренних органов позволяет ей контролировать обменные процессы в организме и осуществлять обеспечение трофических процессов скелетной мускулатуры, рецепторов, кожи и внутренних органов.
Строение вегетативной системы
Работа вегетативной нервной системы контролируется гипоталамусом, который находится в центральной нервной системе. Вегетативная нервная система имеет метасегментарное строение. Ее центры находятся в головном, спинном мозге и коре головного мозга. Периферические отделы образованы стволами, ганглиями, сплетениями.
В вегетативной нервной системе различают:
- Симпатическую. Ее центр расположен в грудопоясничном отделе спинного мозга. Для нее характерны паравертебральные и предвертебральные ганглии ВНС.
- Парасимпатическую. Ее центры сосредоточены в среднем и продолговатом мозге, крестцовом отделе спинного мозга. Нервные узлы в основном интрамуральные.
- Метасимпатическую. Иннервирует желудочно-кишечный тракт, сосуды, внутренние органы организма.
В состав ее входит:
- Ядра нервных центров, расположенных в головном и спинном мозге.
- Вегетативные ганглии, которые расположены по периферии.
- Нервные волокна.
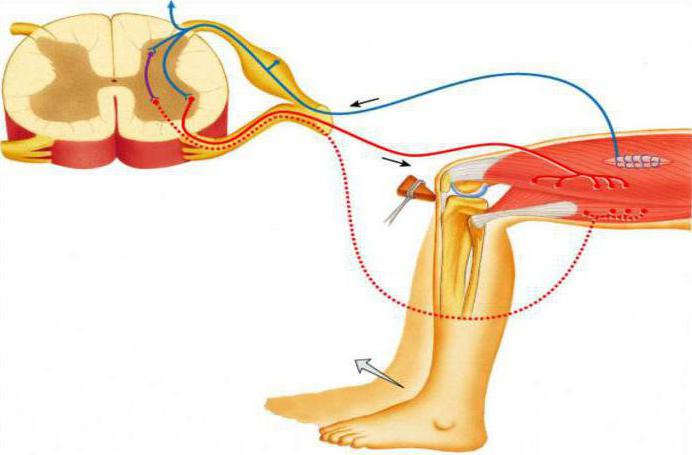
Рефлекторная дуга автономной нервной системы
Рефлекторная дуга вегетативной нервной системы состоит из трех звеньев:
- чувствительное или афферентное;
- вставочное или ассоциативное;
- эффекторное.

Чувствительное звено
Чувствительное звено расположено в спинномозговом ганглии. Этот ганглий имеет нервные клетки, сформированные группами, и их контроль осуществляется ядрами центрального головного мозга, большими полушариями и их структурами.
Чувствительное звено представлено частично униполярными клетками, которые имеют один приносящий или уносящий аксон, и они принадлежат спинальным или черепно-мозговым узлам. А также узлами блуждающих нервов, имеющих строение, похожее на спинальные клетки. В это звено входят клетки Догеля II типа, которые являются компонентами вегетативных ганглиев.
Вставочное звено
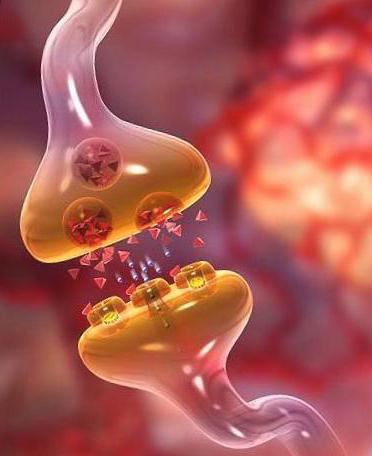
Вставочное звено в вегетативной нервной системе служит для передачи через низшие нервные центры, которыми являются вегетативные ганглии, и осуществляется это через синапсы. Расположено оно в боковых рогах спинного мозга. Нет прямой связи от афферентного звена на преганглионарные нейроны для их связи, существует кратчайший путь от афферентного нейрона до ассоциативного и от него до преганглионарного нейрона. Передача сигналов и нервных импульсов от афферентных нейронов в различных центрах осуществляется с различным количеством вставочных нейронов.
Например, в дуге спинального автономного рефлекса между чувствительным и эффекторным звеном существует три синапса, два из которых расположены в спинном мозге, а один в вегетативном узле, в котором расположен эфферентный нейрон.
Эфферентное звено
Эфферентное звено представлено эффекторными нейронами, которые расположены в вегетативных узлах. Их аксоны образуют безмиелиновые волокна, которые в составе со смешанными нервными волокнами иннервируют внутренние органы.
Вегетативные рефлекторные дуги расположены в боковых рогах.
Строение нервного узла
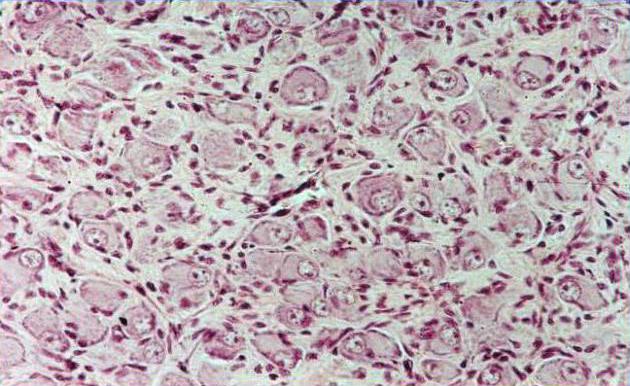
Ганглий – это скопление нервных клеток, которые имеют вид узелковых расширений толщиной около 10 мм. По своему строению вегетативный ганглий сверху покрыт соединительнотканной капсулой, которая образует строму из рыхлой соединительной ткани внутри органов. Мультиполярные нейроны, которые строятся из округлого ядра и крупных ядрышек, состоят из одного эфферентного нейрона и нескольких расходящихся афферентных нейронов. Эти клетки относятся по своему типу к клеткам головного мозга и являются двигательными. Их окружает неплотная оболочка – мантийная глия, которая создает постоянную среду для нервной ткани и обеспечивает полноценное функционирование нервных клеток.
Вегетативный ганглий имеет диффузное расположение нервных клеток и множество отростков, дендритов и аксонов.
Спинномозговой ганглий имеет нервные клетки, которые расположены группами, и их расположение имеет порядок обусловленный.
Вегетативные нервные ганглии разделяются на:
- Сенсорные нейроны, которые расположены близко к спинному или центральному отделу мозга. Униполярные нейроны, из которых состоит этот ганглий, представляют собой приносящий или уносящий отросток. Они служат для афферентной передачи импульсов, и их нейроны образуют бифуркацию при ветвлении отростков. Эти отростки передают информацию от периферии к центральному афферентному нейрону – это периферический отросток, центральный – от тела нейрона в мозговой центр.
- Моторные, двигательные состоят из эфферентных нейронов, и в зависимости от их положения их называют паравертебральные, предвертебральные.
Симпатические ганглии
Паравертебральные цепочки ганглиев расположены вдоль позвоночного столба в симпатических стволах, которые идут длинной вереницей от основания черепа до копчика.
Предвертебральные нервные сплетения находятся ближе к внутренним органам, и их локализация сосредоточена перед аортой. Они образуют брюшное сплетение, которое состоит из солнечного, нижне- и верхнебрыжеечного сплетений. Они представлены двигательными адренергическими и тормозящими действие холинергическими нейронами. Также связь между нейронами осуществляется преганглионарными и постганглионарными нейронами, которые используют медиаторы ацетилхолин и норадреналин.
Интрамуральные нервные узлы имеют нейроны трех типов. Их описание было сделано русским ученым Догелем А.С., который, исследуя гистологию нейронов вегетативной нервной системы, выделил такие нейроны, как длинноаксонные эфферентные клетки первого типа, равноотросчатые афферентные клетки второго типа и ассоциативные клетки третьего типа.
Рецепторы ганглиев
Афферентные нейроны выполняют узкоспециализированую функцию, и их роль заключается в восприятии раздражителей. Такими рецепторами являются механорецепторы (реакция на растяжение или давление), фоторецепторы, терморецепторы, хеморецепторы (отвечают за реакции в организме, химические связи), ноцицепторы (реакция организма на болевые раздражители – это повреждение кожи и другие).
В симпатических стволах эти рецепторы по рефлекторной дуге передают в центральную нервную систему информацию, которая служит сигналом о повреждениях или нарушениях в организме, а также нормальной его работе.
Функции ганглиев
Каждый ганглий имеет свое местоположение, кровоснабжение, и его функции определяются этими параметрами. Спинномозговой ганглий, имеющий иннервацию из ядер головного мозга, обеспечивает непосредственную связь процессов в организме через рефлекторную дугу. От этих структурных компонентов спинного мозга иннервируются железы, гладкая мускулатура мышц внутренних органов. Сигналы, поступающие по рефлекторной дуге, идут медленнее, чем в ЦНС, и они полностью регулируются вегетативной системой, также она обладает трофической, сосудодвигательной функцией.
Шейно-грудной или звездчатый ганглий (ЗГ) относится к паравер-тебральным узлам и располагается в первом межреберье на длинной мышце шеи (И.Ф. Крупачев, 1934; В.Н. Самохвалова, 1958), позади подключичной артерии. У животных он образуется в результате слияния каудального шейного с первыми узлами грудной части пограничного симпатического ствола. По мнению одних авторов, ЗГ сформирован путем слияния каудального шейного с первым грудным (В.Н Самохвалова, 1958, ЕПеп-berger-Baum, 1943), а других - слиянием каудального шейного с тремя краниальными грудными (Ф.С. Гусев, 1958; А.Д. Ноздрачев, Е.Л. Поляков, 2001). Ветви ЗГ участвуют в образовании сердечного, подключичного, щитовидного, внутреннего грудного, позвоночного сплетений, по которым отростки нейронов достигают органов шеи и грудной полости (Abraham, 1951).
По данным Ленгли и др. авторов, через ЗГ осуществляются пиломо-торные, вазомоторные, секреторные и трофические симпатические эффекты в передней конечности, на шее и груди, а также ускорение ритма сердечных сокращений. Вместе с ускоряющими, от ЗГ идут к сердцу волокна, усиливающие сердечные сокращения (И.П.Павлов, 1883; Langley, 1891, 1899). Возбуждение ЗГ уменьшает асинхронность возбуждения и сокращения волокон миокарда, повышает скорость проведения возбуждения в миокарде (Priola, Randall, 1964) и ведет к расширению коронарных сосудов (Brown, 1968). ЗГ посылает к легким бронходилятаторные волокна (Daly, Mount, 1951), волокна к пищеводу (М.В.Сергиевский, 1964), к желудку (Jourdan, 1957), а также небольшое количество волокон в блуждающий нерв через подключичную петлю (Lanlgey, 1899). ЗГ является самым крупным узлом грудного отдела пограничного симпатического ствола. Его размеры у различных видов животных составляют: у морской свинки длина узла 6-10, ширина — 1.0 — 1.5 мм; у кошки длина - 4 — 6, ширина — 2.5 - 3.0 мм; у собаки длина — 20 — 25, ширина — 5 - 7 мм (Б. М.Эрез, 1957; А.Д. Ноздрачев, М.М. Фатеев, 2002); у человека длина узла составляет от 8 до 42, а ширина — от 10 до 20 мм (Т.В. Золотарева с соавт., 1979).
Форма узла различна у разных видов млекопитающих и даже у представителей одного вида она довольно сильно варьирует от типично звездчатой до сильно вытянутой в рострокаудальном направлении. Так, у человека выделено 6 форм звездчатого узла (Т.В Золотарева с соавт., 1979), отличающихся не только по внешнему виду, но и по числу ветвей, выходящих из ганглия. Форма ганглия зависит от полного или неполного слияния нижнего шейного и первого грудного симпатических узлов.
В настоящее время проведено довольно большое число исследований, касающихся клеточного состава паравертебральных ганглиев. Было обнаружено, что количество, размеры, форма и расположение нейронов в вегетативных узлах сильно варьирует.
Наибольшее количество нервных клеток ганглиев у млекопитающих имеют размер тела от 15 до 60 мкм (Ю.М.Жаботинский, 1953; В.П.Бабминдра, 1972). В краниальном шейном (КШГ) и ЗГ кошки наибольшее число нейронов имеет максимальный диаметр 20 - 40 мкм (В.П.Бабминдра с соавт., 1982; А.Д. Ноздрачев, М.М. Фатеев, 2002; П.М. Маслюков, 2003; Masliukov, 2001). В КШГ взрослой мыши средний диаметр тел нейронов составляет 34, в чревном — 32, в грудном симпатическом - 31 мкм (Jobling and Gibbins, 1999). В ЗГ макак размер тел нейронов варьирует от 10 до 50 мкм (Leong et al., 1989). У человека в соответствии с клеточным объемом различают большие клетки (с максимальным диаметром 36 — 60 мкм), средние (25 — 35 мкм) и малые (15 - 25 мкм) (Ю.М.Жаботинский, 1953).
Средняя площадь сечения нейронов КШГ, чревного и грудного симпатического узлов у мыши составляет 712,0+92,5, 602,5±120,7, 637,8+51,1 мкм" , соответственно (Jobling and Gibbins, 1999). Площадь сечения нейро-нов ЗГ у кошки равняется 634,2±3,64 мкм" (А.Д. Ноздрачев, М.М. Фатеев, 2002). Площадь поперечного сечения нейронов ЗГ крысы составляет 489 — 754 мкм2 (Richardson et al., 2006). Результаты исследований морфометри-ческих параметров нейроцитов свидетельствуют, что размеры нейронов в постнатальном онтогенезе увеличиваются. Так, площадь сечения нейронов чревного узла морской свинки на ранней фетальной стадии составляет 170 мкм" и достигает 2000 мкм" у взрослых животных (Anderson et al., 2001). Площадь сечения нейроцитов ЗГ у новорожденного котенка - 132,2+0,87 мкм" , у шестимесячного равняется 626,5+3,67 мкм (Masliukov, 2001). Колебания размеров тела нейрона встречаются среди нервных клеток одного и того же ганглия при сравнении различных ганглиев, а также наблюдается группирование более крупных нейроцитов среди равномерно распределенных мелких (Н.Г.Колосов, 1968; В.И. Скок с соавт., 1989; Gabella, 1976).
Нейроны симпатических узлов млекопитающих имеют округлую, овальную, грушевидную или полигональную форму тел и в основном являются мультиполярными (А.С. Догель, 1896; Н.Г. Колосов с соавт., 1978; Б.И. Швалев с соавт., 1992; Jobling and Gibbins et al., 1999; Anderson et al., 2001). Количество отростков может колебаться от 3 до 20 и, например, для КШГ крысы, их число равно в среднем 13 (McLachlan, 1974), у мыши - от 1 до 5 (Jobling and Gibbins et al., 1999). Уни - и биполярные клетки встречаются сравнительно редко. Мультиполярные клетки обычно одинакового вида, однако в звездчатом узле человека встречаются нейроны с чрезвычайно длинным дендритом (A braham, 1951). Согласно данным, приведенным в обзоре Габеллы (Gabella, 1976), у крысы КШГ содержит в среднем 42 000 нейронов, у кошки - около 100 000 нейронов, у человека — 911 000 нейронов. Таким образом, с увеличением размеров тела количество нейронов в ганглии возрастает. Увеличивается и средний диаметр тела нейронов (с 15,5 мкм у крысы до 35 мкм у быка), а также количество и длина их ден-дритов. Нейроны симпатических ганглиев новорожденных значительно меньше по размеру, чем у взрослых животных, и имеют несколько коротких и маловетвящихся отростков (B.C. Шевелева, 1977; Hirst, McLachlan, 1986). Имеются данные об изменении числа нейронов в паравертебраль-ных узлах в раннем постнатальном онтогенезе. У крыс в КШГ и шейно-грудном узле число нейронов значительно уменьшается в течение первой постнатальной недели с 39500 до 26000 и не изменяется до 2 месяца жизни (Wright et al., 1983). В ЗГ кошки количество нервных клеток также уменьшается в первые 10 дней жизни с 2,11х105 до 1,86х105 (Masliukov, 2001).
В структуре ЗГ основными элементами являются его волокна и нервные клетки. Снаружи ганглий покрыт соединительнотканной капсулой, от которой внутрь отходят перегородки, разделяющие клетки на отдельные группы. Между скоплениями нейронов проходят нервные волокна (Н.Г. Колосов, А.Я. Хабарова, 1978).
Все нейроны ЗГ окружены клетками-сателлитами или швановскими клетками. По ультраструктурной организации и набору цитоплазматиче-ских органелл нейроциты симпатических ганглиев существенно не отличаются от нейронов центральной нервной системы (В.П.Бабминдра, 1972; В.Н. Швалев с соавт., 1992).
Спинномозговые ганглии – округлые или овальные тельца, расположены по бокам спинного мозга на дорсальных корешках спинномозговых нервов и около головного мозга на чувствительных черепномозговых нервах. Ганглии одеты капсулой из соединительной ткани, которая проникает внутрь узла в виде тонких прослоек, образующих их остовы. В прослойках проходят сосуды. Размеры ганглиев от микроскопических до 2 см.Ганглии – скопления ложноуниполярных чувствительных нейронов. Тела округлой формы, содержат светлые крупные округлые ядра с крупным ядрышком и имеют хорошо развитый пластинчатый комплекс Гольджи в виде многочисленных стопок цистерн. Нейроны окружены клетками нейроглии.Их дендриты в виде миелиновых нервных волокон идут на периферию в составе спинномозгового нерва, а аксоны формируют дорсальный корешок спинномозгового нерва, входящего в спинной мозг. Разновидностью биполярных нейронов является псевдо-униполярный нейрон, от тела которого отходит один общий вырост — отросток, разделяющийся затем на дендрит и аксон. Псевдоуниполярные нейроны присутствуют в спинальных ганглиях, биполярные — в органах чувств. Большинство нейронов - мультиполярные. Их формы чрезвычайно разнообразны. Аксон и его коллатерали оканчиваются, разветвляясь на несколько веточек, называемых телодендронами, последние заканчиваются терминальными утолщениями.Нейроглия, или просто глия — Сложный комплекс вспомогательных клеток нервной ткани, общий функциями и, частично, происхождением (исключение — микроглия). Глиальные клетки составляют специфическое микроокружение для нейронов, обеспечивая условия для генерации и передачи нервных импульсов, а также осуществляя часть метаболических процессов самого нейрона. Нейроглия выполняет опорную, трофическую, секреторную, разграничительную и защитную функции.
3. Развитие, строение и функции вегетативных ганглиев.
Вегетативная нервная система (ВНС) координирует и регулирует деятельность внутренних органов, обмен веществ, гомеостаз. Ее деятельность подчинена центральной нервной системе и в первую очередь коре головного мозга.ВНС состоит из симпатического и парасимпатического отделов. Оба отдела иннервируют большинство внутренних органов и часто оказывают противоположное действие. Центры ВНС расположены в четырех отделах головного и спинного мозга. Импульсы из нервных центров к рабочему органу проходят по двум нейронам. В процессе эмбриогенеза происходит увеличение количества клеток в ганглиях, приводящее на первых этапах к густому расположению их в узлах. Позже, по мере развития в узлах соединительной ткани, клетки располагаются менее плотно. Увеличиваются и размеры клеток, некоторые из них на поздних стадиях эмбриогенеза становятся крупными, способными вступить в синаптическуюсвязь.Зачатокинтрамуральных сплетений представлен отдельными нейробластами, количество которых увеличивается, они собираются в группы (образования нервных узлов), в которых отдельные клетки нейробластического ряда находятся в состоянии митоза (пищевод 15- и 20-дневных зародышей, 12-перстная кишка 20-дневного зародыша кролика). Возле этих клеток располагаются мелкие элементы глии. Появляются мультиполярные нейроны с короткими отростками, их сопровождают глиальные клетки. Ганглий окружен соединительнотканной капсулой, содержащей преколлагеновые волокна (20-дневный зародыш). Внутри ганглия соединительная ткань имеет редкие еще преколлагеновые волокна и капилляры. Большинство клеток интрамуральных узлов старших зародышей и новорожденных - еще нейробласты. Только отдельные нейроны достигают больших размеров и могут вступать в синаптические связи. Физиологические наблюдения показывают, что в это время (у кролика с 22-23-го дня эмбриогенеза) раздражение блуждающего и чревного нервов вызывает усиление спонтанных сокращений 12-перстной кишки. Подобного эффекта не получается у 21-дневного зародыша. В 12-перстной кишке раньше, чем в других отделах кишечника, появляются ритмические, а затем и перистальтические сокращения в соответствии с развитием мышечных слоев (циркулярного и продольного).
4. Развитие спинного мозга.
Белое вещество спинного мозга представлено отростками нервных клеток, которые составляет тракты, или проводящие пути спинного мозга:
1) короткие пучки ассоциативных волокон , связывающие сегменты спинного мозга, расположенные на различных уровнях;
2) восходящие (афферентные, чувствительные) пучки, направляющиеся к центрам большого мозга и мозжечка ;
3) нисходящие (эфферентные, двигательные) пучки, идущие от головного мозга к клеткам передних рогов спинного мозга .
Белое вещество спинного мозга располагается по периферии серого вещества спинного мозга и представляет собой совокупность миелинизированных и отчасти маломиелинизированныхнервных волокон , собранных в пучки. В белом веществе спинного мозга расположены нисходящие волокна (идущие из головного мозга) и восходящие волокна , которые начинаются от нейронов спинного мозга и проходят в головной мозг . По нисходящим волокнам передается преимущественно информация от моторных центров головного мозга к мотонейронам (двигательным клеткам) спинного мозга. По восходящим волокнам поступает информация как от соматических, так и от висцеральных чувствительных нейронов. Расположение восходящих и нисходящих волокон носит закономерный характер. На спинной (дорсальной) стороне расположены преимущественно восходящие волокна , а на брюшной (вентральной) - нисходящие волокна .
Борозды спинного мозга разграничивают белое вещество каждой половины на передний канатик белого вещества спинного мозга ,боковой канатик белого вещества спинного мозга и задний канатик белого вещества спинного мозга ( рис. 7 ).
Передний канатик ограничен передней срединной щелью и переднебоковой бороздой . Боковой канатик расположен между переднебоковой бороздой и заднебоковой бороздой . Задний канатик находится между задней срединной бороздой и заднебоковой бороздой спинного мозга .
Белое вещество обеих половин спинного мозга связано двумя комиссурами (спайками): дорсальной, лежащей под восходящими путями, и вентральной, находящейся рядом с моторными столбами серого вещества .
6. Строение серого вещества спинного мозга. Характеристика нейроцитов серого вещест-ва спинного мозга.
Нейроциты в сером веществе окружены спутанными как войлок нервными волокнами - нейропилью. Аксоны в нейропилеслабомиелинизированы, а дендриты и вовсе не миелинизированы. Сходные по размерам, тонкому строению и функциям нейроциты СМ располагаются группами и образуют ядра.
Среди нейроцитов СМ различают следующие типы:
1. Корешковые нейроциты - располагаются в ядрах передних рогов, по функции являются двигательными; аксоны корешковых нейроцитов в составе передних корешков покидают СМ , проводят к скелетной мускулатуре двигательные импульсы.
2. Внутренние клетки - отростки этих клеток не покидают пределы серого вещества СМ, оканчиваются в пределах данного сегмента или соседнего сегмента, т.е. по функции являются ассоциативными.
3. Пучковые клетки - отростки этих клеток образуют нервные пучки белого вещества и направляются в соседние сегменты или вышележащие отделы НС, т.е. по функции тоже являются ассоциативными.
Задние рога СМ более короткие, узкие и содержат следующие виды нейроцитов:
а) пучковые нейроциты - располагаются диффузно, получают чувствительные импульсы от нейроцитов спинальных ганглиев и передают по восходящим путям белого вещества в вышележащие отделы НС (в мозжечок, в кору больших полушарий);
б) внутренние нейроциты - передают чувствительные импульсы со спинальных ганглиев в двигательные нейроциты передних рогов и в соседние сегменты.
7. Строение белого вещества спинного мозга.
Белое вещество спинного мозга представлено отростками нервных клеток, которые составляет тракты, или проводящие пути спинного мозга:
1) короткие пучки ассоциативных волокон , связывающие сегменты спинного мозга, расположенные на различных уровнях;
2) восходящие (афферентные, чувствительные) пучки, направляющиеся к центрам большого мозга и мозжечка ;
3) нисходящие (эфферентные, двигательные) пучки, идущие от головного мозга к клеткам передних рогов спинного мозга .
Белое вещество спинного мозга располагается по периферии серого вещества спинного мозга и представляет собой совокупность миелинизированных и отчасти маломиелинизированныхнервных волокон , собранных в пучки. В белом веществе спинного мозга расположены нисходящие волокна (идущие из головного мозга) и восходящие волокна , которые начинаются от нейронов спинного мозга и проходят в головной мозг . По нисходящим волокнам передается преимущественно информация от моторных центров головного мозга к мотонейронам (двигательным клеткам) спинного мозга. По восходящим волокнам поступает информация как от соматических, так и от висцеральных чувствительных нейронов. Расположение восходящих и нисходящих волокон носит закономерный характер. На спинной (дорсальной) стороне расположены преимущественно восходящие волокна , а на брюшной (вентральной) - нисходящие волокна .
Борозды спинного мозга разграничивают белое вещество каждой половины на передний канатик белого вещества спинного мозга ,боковой канатик белого вещества спинного мозга и задний канатик белого вещества спинного мозга ( рис. 7 ).
Передний канатик ограничен передней срединной щелью и переднебоковой бороздой . Боковой канатик расположен между переднебоковой бороздой и заднебоковой бороздой . Задний канатик находится между задней срединной бороздой и заднебоковой бороздой спинного мозга .
Белое вещество обеих половин спинного мозга связано двумя комиссурами (спайками): дорсальной, лежащей под восходящими путями, и вентральной, находящейся рядом с моторными столбами серого вещества .
В составе белого вещества спинного мозга различают 3 группы волокон (3 системы проводящих путей):
- короткие пучки ассоциативных (межсегментных) волокон, связывающие участки спинного мозга на различных уровнях;
- длинные восходящие (афферентные, чувствительные) проводящие пути, которые идут от спинного мозга к головному;
- длинные нисходящие (эфферентные, двигательные) проводящие пути, идущие от головного мозга к спинному.
Межсегментные волокна образуют собственные пучки, расположенные тонким слоем по периферии серого вещества и осуществляющие связи между сегментами спинного мозга. Они присутствуют в переднем, заднем и боковом канатиках.
Большую часть переднего канатика белого вещества составляют нисходящие проводящие пути.
В боковом канатике белого вещества есть и восходящие, и нисходящие пути. Они начинаются как из коры больших полушарий , так и от ядер ствола головного мозга.
В заднем канатике белого вещества расположены восходящие проводящие пути. В верхней половине грудной части и в шейной части спинного мозга задняя промежуточная борозда спинного мозга делит задний канатик белого вещества на два пучка: тонкий пучок (пучок Голля) , лежащий медиально , и клиновидный пучок (пучок Бурдаха) , расположенный латерально . Тонкий пучок содержит афферентные пути , идущие от нижних конечностей и от нижней части тела. Клиновидный пучок состоит из афферентных путей , проводящих импульсы от верхних конечностей и от верхней части тела. Разделение заднего канатика на два пучка хорошо прослеживается в 12 верхних сегментах спинного мозга начиная с 4-го грудного сегмента.
8. Характеристика нейроглии спинного мозга.
Нейроглия состоит из макро - и микроглиальных клеток. К нейроглиальным элементам также относят эпендимные клетки , которые у некоторых животных сохраняют способность к делению.
Макроглия подразделяется на астроциты, или лучистые глиоциты , и олигодендроциты . Астроциты - самые разнообразные глиальные клетки, имеющие звездчатую или паукообразную форму. Астроцитарнаяглия состоит из протоплазматических и фиброзных астроцитов .
В сером веществе мозга встречаются преимущественно протоплазматическиеастроциты. Их тело имеет относительно крупные размеры (15-25 мкм) и многочисленные ветвистые отростки.
В белом веществе мозга располагаются волокнистые, или фиброзные, астроциты. У них небольшое тело (7-11 мкм) и длинные малоразветвленные отростки.
Астроциты - единственные клетки, располагающиеся между капиллярами и телами нейронов и участвующие в транспорте веществ из крови к нейронам и транспорте продуктов метаболизма нейронов обратно в кровь. Астроциты формируют гематоэнцефалический барьер . Он обеспечивает избирательное прохождение из крови в ткань мозга различных веществ. Благодаря гематоэнцефалическому барьеру в экспериментах многие продукты обмена, токсины, вирусы, яды при введении в кровь почти не обнаруживаются в спинномозговой жидкости.
Олигодендроциты - мелкие (размеры тела около 5-6 мкм) клетки со слаборазветвленными, относительно короткими и немногочисленными отростками. Одна из основных функций олигодендроцитов - формирование оболочек аксонов в ЦНС .Олигодендроцит наматывает свою мембрану вокруг нескольких аксонов нервных клеток, образуя многослойную миелиновую оболочку . Олигодендроциты выполняют еще одну очень важную функцию - они участвуют в нейронофагии (от греч. фагос - пожирающий), т.е. удаляют омертвевшие нейроны путем активного поглощения продуктов распада.

Механическое удерживание земляных масс: Механическое удерживание земляных масс на склоне обеспечивают контрфорсными сооружениями различных конструкций.

Организация стока поверхностных вод: Наибольшее количество влаги на земном шаре испаряется с поверхности морей и океанов (88‰).

Опора деревянной одностоечной и способы укрепление угловых опор: Опоры ВЛ - конструкции, предназначенные для поддерживания проводов на необходимой высоте над землей, водой.
Впервые в эволюции НС появляется у типа кишечнополостные. Это диффузная НС. Кишечнополостные обладают радиальной (лучевой) симметрией и имеют две жизненные формы — прикрепленные полипы и свободноплавающие медузы. Тело у них состоит из двух слоев клеток — эктодермы и энтодермы. Основу каждого слоя составляют эпителиальномускульные клетки. Таким образом, моторика кишечнополостных бесске- летная, носящая перистальтический характер. Нейроны расположены как в экто-, так и в эктодермальном слоях. Сенсорные функции выполняют специализированные эктодермальные нервные клетки, чувствительные волоски которых выходят на поверхность тела и воспринимают раздражения (см. рис. 16.2, б). Отдельные части тела животных с диффузной НС способны к автономным движениям за счет того, что в каждом участке есть чувствительная клетка, двигательная клетка и нервное волокно. Но даже у самых примитивных кишечнополостных можно найти элементы концентрации — нейроны около рта и на подошве располагаются гуще, чем на остальном теле.
Нервные сети кишечнополостных могут иметь синцитиальное строение, т.е. отростки нервных клеток могут сливаться между собой, проводя возбуждение в разные стороны. Но в большинстве случаев возбуждение передается с помощью типичных синапсов, большинство которых поляризовано, т.е. проводят возбуждение только в одну сторону.
У медуз, которые способны к более активному передвижению, НС несколько усложняется: но краю зонтика у них наблюдаются скопления нервных клеток, которые вместе со своими отростками образуют сплошное нервное кольцо. Таким образом, концентрация нервных элементов у медуз больше, чем у полипов. Более того, у медуз появляются примитивные органы чувств двух типов — статоцисты и глазки — поверхностные скопления светочувствительных клеток. Статоцисты — это органы равновесия, которые можно встретить почти у всех беспозвоночных животных. В большинстве случаев они представляют собой замкнутые пузырьки, на внутренних стенках которых лежат чувствительные клетки. Полость пузырька заполнена жидкостью, в которой находится сгатолит — кристаллик углекислой извести. При изменении положения тела в пространстве чувствительные клетки испытывают давление и возбуждаются.
Животные, следующие в эволюционном развитии за кишечнополостными, — это разные типы червей. Все животные, расположенные на эволюционном древе выше кишечнополостных, — животные трехслойные, проходящие в процессе онтогенеза стадию трех зародышевых листков, которые в своем дальнейшем развитии дают ткани. Таким образом, даже у самых примитивных червей — плоских — уже есть гладкая мышечная ткань, но движения сохраняют перистальтический характер. Концентрация нейронов, только намечавшаяся у медуз, приобретает у червей вполне выраженный характер — появляются нервные ганглии. Но в отличие от кишечнополостных, у которых сокращение мускульных волоконец охватывает все тело, черви способны последовательно сокращать отдельные участки тела, что возможно благодаря иннервации разных участков нейронами, расположенными в разных ганглиях.
Принципиальное отличие червей, как и всех остальных беспозвоночных (за исключением гребневиков и иглокожих) от кишечнополостных, — это наличие билатеральной (двусторонней) симметрии. В связи с этим у них появляется передний конец тела, в сторону которого обычно осуществляется движение. Вследствие этого на нем концентрируются важнейшие органы чувств и, соответственно, передние ганглии укрупняются.
В типе плоские черви, самом примитивном типе червей (например, пла- нария, бычий цепень), централизация нервных клеток еще очень невелика. Их НС состоит из парного головного ганглия и нескольких продольных стволов, соединенных кольцевыми перемычками. Начиная с круглых червей (например, аскарида) у беспозвоночных появляется окологлоточное нервное кольцо, окружающее переднюю часть пищеварительной трубки. Оно обычно состоит из парных надглоточных ганглиев, которые часто сливаются, образуя так называемый головной мозг, и непарного подглоточного ганглия, соединенных нервными волокнами.
В наиболее выраженном виде узловая НС наблюдается в типах кольчатые черви (самые высокоорганизованные черви, например, дождевой червь или нереида), членистоногие и моллюски.
Тело кольчатых червей и членистоногих построено но метамерному принципу и состоит из хорошо выраженных сегментов. От подглоточного узла начинается брюшная нервная цепочка. Исходно (у кольчатых червей) она состоит из парных ганглиев, соединенных комиссурами. Части нервных стволов, соединяющие ганглии каждой стороны соседних сегментов, называются коннективами (см. рис. 16.3, а). Вся цепь имеет характерный вид лестницы (лестничная НС). У разных видов кольчецов наблюдается постепенное сближение и слияние парных ганглиев, при этом коннективы остаются парными. В дальнейшем объединяются и коннективы, и в своем классическом виде брюшная нервная цепочка выглядит как на рис. 16.3, б.
Расположение ганглиев на брюшной стороне связано, по-видимому, с расположением основных рецептивных областей. Черви передвигаются путем ползания и получают многие сенсорные импульсы от вентральной поверхности. В передней же части тела наибольшее значение имеет рецепция, получаемая от рострального конца, в связи с чем головной ганглий поднимается вверх.
НС членистоногих (ракообразные, паукообразные, насекомые) устроена значительно совершеннее, чем у кольчатых червей. Это связано как с более сложным поведением, так и с особенностями организации. Для этих животных характерна гетерономная сегментация, т.е. сегменты тела не одинаковы, как у кольчатых червей, а отличаются друг от друга как по строению, так и по функциям. Конечности состоят из нескольких члеников, подвижно соединенных друг с другом и с телом с помощью суставов. Следовательно, принципиально меняется характер движения — он становится рычаговым. Помимо этого, большая часть гладкой мускулатуры заменяется поперечнополосатой (впервые в эволюции), что способствует гораздо более быстрым движениям. Что касается НС, то концентрация и централизация нейронов (если говорить о беспозвоночных) достигает своего максимума. Нередко наблюдается слияние нервных узлов брюшной нервной цепочки с образованием более крупных ганглиев, т.е. наблюдается и продольная концентрация элементов НС.
Но особенно сильно НС членистоногих отличается от НС червей в строении ее головного конца. У большинства этих животных ГМ (надглоточный ганглий) состоит из трех отделов: переднего — протоцеребрума, среднего — дейтоцеребрума и заднего — тритоцеребрума. Каждый отдел головного мозга иннервирует разные органы чувств и состоит из нескольких ганглиев, отделенных друг от друга белым веществом. В протоцере- бруме расположены особенно важные ассоциативные центры — грибовидные тела (рис. 16.5).
Наибольшее развитие головного мозга наблюдается у общественных насекомых (муравьев, пчел, термитов), т.е. у животных с особенно сложным поведением. Причем даже в пределах одного вида величина мозга может значительно варьироваться в зависимости от сложности поведения. Так, например, у рабочих муравьев грибовидные тела развиты значительно сильнее, чем у цариц и самцов.
Что касается моллюсков, то это резко отграниченный тип беспозвоночных, ведущий начало от древних кольчатых червей, но имеющий несегмен- тированное тело. От окологлоточного кольца у моллюсков отходят четыре продольных нервных ствола. У высших форм моллюсков на стволах формируются парные ганглии, соединенные коннективами, некоторые ганглии соединяются и с помощью комиссур (рис. 16.6). Нервные клетки имеются не только в ганглиях, но и рассеяны в периферических нервах. Такой тип нервной системы называют разбросанно-узловым.

Рис. 16.5. Нервная система насекомого

Рис. 16.6. Нервная система брюхоногого моллюска
У высших моллюсков крупные ганглии расположены, как правило, рядом с иннервируемым органом или непосредственно в его стенках. Надглоточные ганглии иннервируют глаза и статоцисты, глотку и щупальца; ганглии, иннервирующие мускулатуру ноги, расположены в самой ноге; рядом с ними лежат ганглии, иннервирующие мантию; следующая пара ганглиев связана с жабрами и органами химического чувства; еще одна пара управляет внутренними органами.
Особенно высокой сложности строения НС достигает у головоногих моллюсков (рис. 16.7).

Рис. 16.7. Головной мозг головоногого моллюска
Большинство этих животных — свободноплавающие и очень подвижные хищники. Для них характерно настолько сложное поведение, что их иногда называют приматами моря. Ганглии у них очень велики и образуют сплошную окологлоточную нервную массу (ГМ). В этой общей массе выделяют отдельные ганглии, выполняющие различные функции. В мозге головоногих даже найдены участки (в зрительной доле), имеющие слоистое строение. Для защиты такой НС головоногие имеют подкожную хрящевую капсулу, окружающую мозг аналогично черепу позвоночных.
Читайте также: