
Точное описание нервной системы малощетинковых червей
Эти организмы играют колоссальную роль при формировании почвы - верхнего плодородного слоя земли. Типичным представителем класса малощетинковых является дождевой червь - почвенная форма, встречаются также и водные формы - трубочник (излюбленный корм многих рыб).

Имеет типичную червеобразную форму, в длину достигает 15-30 см.
-
Покровы тела, опорно-двигательная система
Специализированные органы движения отсутствуют, их тело снаружи покрыто немногочисленными хитинообразными выростами - щетинками (чему соответствует название класса - малощетинковые). На каждом сегменте дождевого червя расположено по 8 щетинок попарно или поодиночке.
Щетинки играют важную роль при движении, они подобно рычагам, помогают найти червю точку опоры и способствуют его продвижению в почве. Щетинки в прямом смысле "спасают" дождевого червя, выполняя защитную функцию - удерживая его в почве, если им захотят пообедать. Именно из-за них дождевого червя так трудно вытащить из его норки.

Однослойный кожный эпителий образует на поверхности кожи эластичную кутикулу. Многочисленные слизистые клетки, расположенные в эпителии, выделяют секрет - слизь, которая выполняет защитную функцию, препятствуя пересыханию тела, и способствует газообмену.
Под эпителием лежат кольцевые и продольные мышцы червя, образующие мышечную систему. Поверхность мышц и внутренних органов выстилается изнутри особым эпителием мезодермального происхождения - целотелием.
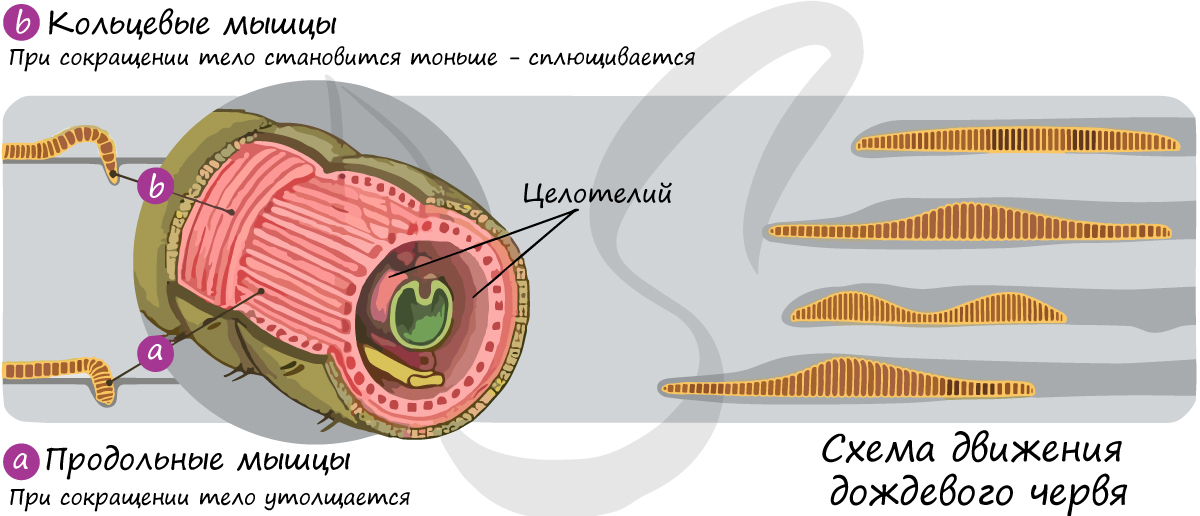
В совокупности слой кутикулы, эпителия, кольцевых и продольных мышц составляет кожно-мускульный мешок. Между кожно-мускульным мешком и внутренними органами находится пространство, вторичная полость тела, заполненная жидкостью и выполняющая ряд важнейших функций: опорную, транспортную, выделительную и защитную.

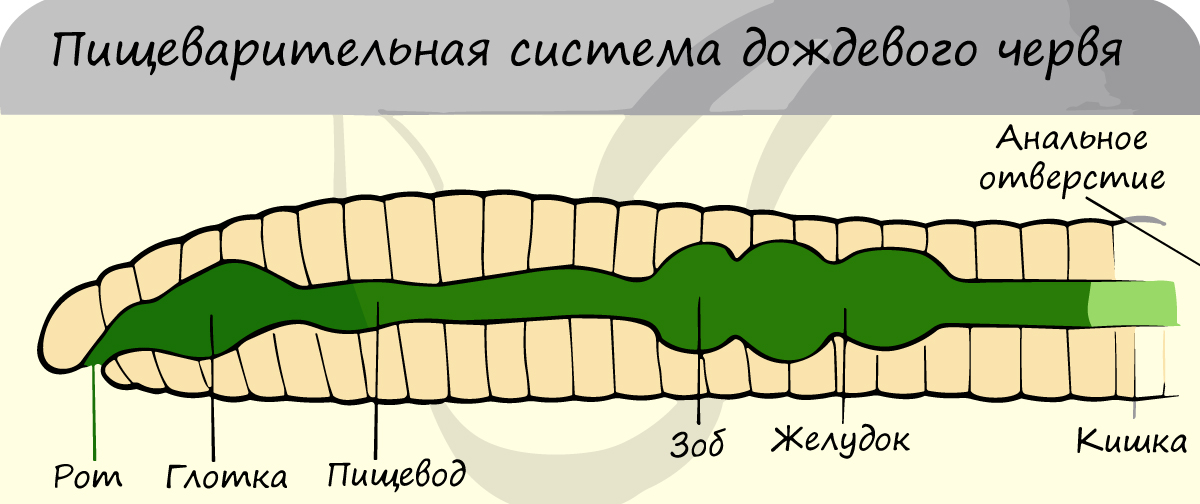
Состоит из трех отделов: переднего, среднего и заднего. Передний отдел начинается ротовым отверстием, расположенным на переднем конце тела. Далее следует глотка, переходящая в узкий пищевод, в котором имеются 3 парных выпячивания - известковые железы. Эти железы выделяют в просвет пищевода углекислый кальций - CaCO3, который нейтрализует находящиеся в почве гуминовые кислоты, таким образом, уменьшая кислотность почвы, что повышает ее плодородие.
Передний отдел также включает в себя следующий после пищевода зоб - расширенную часть пищевода, которая служит для накопления, хранения и предварительной обработки пищи и заканчивается мускулистым желудком (может быть один или несколько), где пища измельчается.
В среднем отделе - кишке, происходит переваривание и всасывание расщепленных веществ. Здесь имеется глубокое продольное выпячивание стенки кишки - тифлозоль, которое значительно увеличивает поверхность всасывания. Такого выпячивания в заднем отделе нет, непереваренные остатки пищи выводятся из задней кишки через анальное отверстие и попадают во внешнюю среду, значительно улучшая состав и свойства почвы.
За счет наличия анального отверстия пищеварительная система у дождевого червя сквозного типа, это означает, что для него возможно непрерывное питание.
Специализированные органы отсутствуют, дыхание осуществляется диффузно всей поверхностью тела. Интересно заметить, что известковые железы тоже участвуют в процессах, связанных с дыханием, так как способствуют удалению углекислоты из крови в пищеварительный тракт в виде CaCO3.
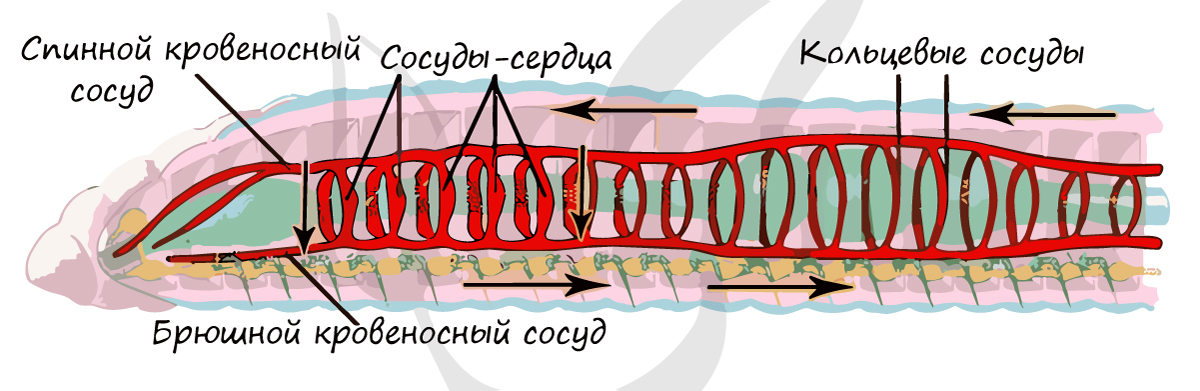
Кровь движется только по сосудам, не изливаясь в полость тела, такой тип кровеносной системы называется - замкнутый. Выделяют спинной и брюшной сосуды, соединенные кольцевыми перемычками. Настоящего сердца нет, его заменяют 5 крупных кольцевых сосуда - "сосуды-сердца", которые, сокращаясь, обеспечивают ток крови.
В сосуды кишечного сплетения, оплетающего кишечник, попадают расщепленные питательные вещества, которые с током крови доставляются к органам и тканям организма.
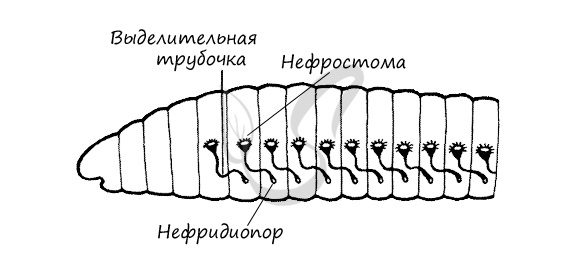
Звездчатых клетки (соленоциты) у кольчатых червей исчезают. Их органы выделения представлены метанефридиями.
Метанефридии (от греч. meta - между, через и греч. nephros — почка) - парные органы выделения, имеющие вид извитых железистых трубок (эктодермального происхождения), которые одним концом - мерцательной вороной (нефростомой) открываются во вторичную полость тела - целомические мешки, а другим концом - нефридиопором - во внешнюю среду. Эволюционно метанефридии образовались из протонефридиев.
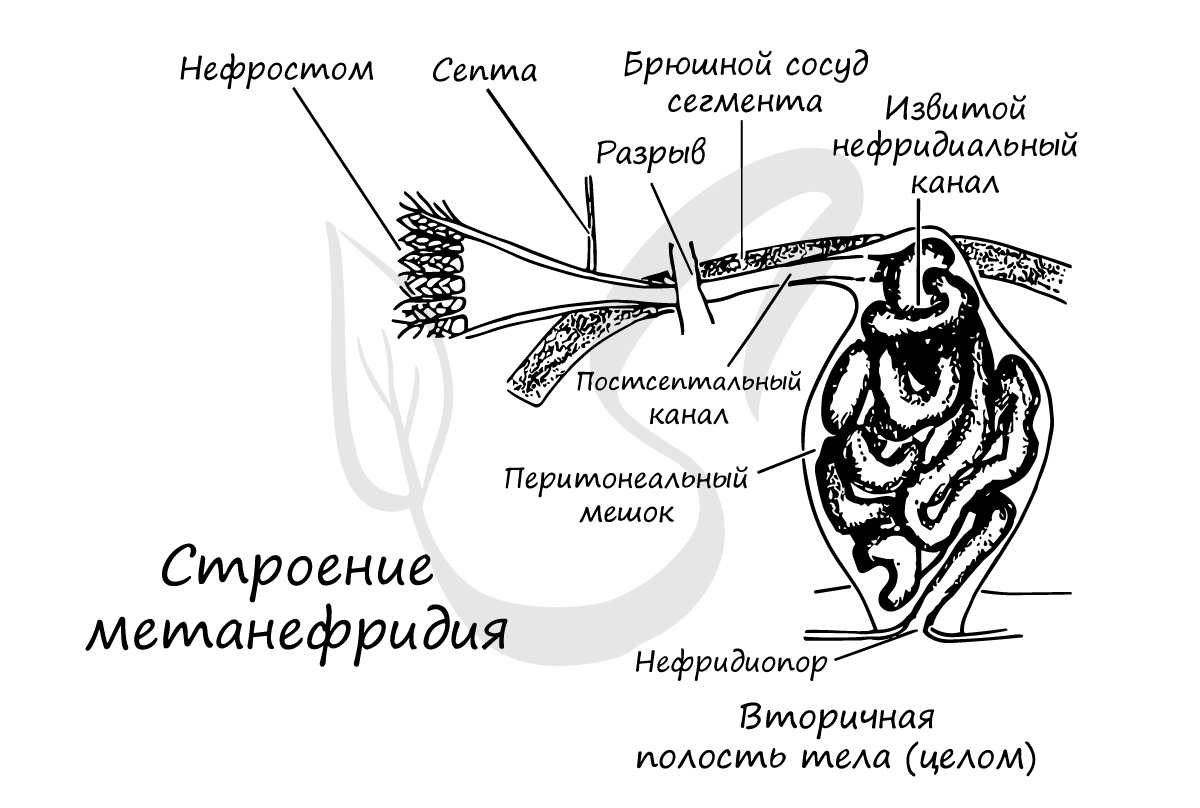
Обращаю ваше пристальное вниманием на следующую особенность: канал, идущий от нефростомы, пронизывает перегородку между двумя сегментами и заканчивается в следующем сегменте выделительным отверстием (нефридиопорой), обращенным во внешнюю среду.
Парные надглоточные нервные узлы соединяясь коннективами (коннективы в отличие от комиссур соединяют разноименные ганглии) с подглоточными нервными узлами вместе образуют единую структуру - окологлоточное нервное кольцо.
Надглоточные ганглии развиты сильнее подглоточных, поэтому их часто называют головным мозгом. Он слаборазвит, но такое название, прежде всего, подчеркивает цефализацию нервных структур - крайне важное эволюционное явление.
От окологлоточного нервного кольца отходит брюшная нервная цепочка, состоящая из нервных тяжей, на которых находятся ганглии по паре в каждом сегменте. От нервных ганглиев отходят ветви, иннервирующие внутренние органы и ткани данного сегмента червя.

В целом у кольчатых червей половая система может быть как раздельнополой, так и гермафродитного плана. В частности для дождевых червей характерен гермафродитизм. Половые органы расположены в строго определенных сегментах тела.
В 10 и 11 сегментах расположено по паре семенников, с идущими от них семявыносящими каналами и семяпроводами, открывающимися на брюшной стороне тела в 15 сегменте. В 9 и 10 сегментах имеются кожные выпячивания - семяприемники, в которых накапливается семенная жидкость партнера во время копуляции.
В 13-м сегменте локализуются яичники с яйцеводами, которые пронизывают перегородку между сегментами, и в следующем членике открывают половыми отверстиями на брюшной стороне.

Оплодотворение обычно перекрестное - участвуют две особи. Брюшные поверхности тела двух копулирующих особей сближаются, в это время железистые клетки пояска начинают выделять быстрозатвердевающущю жидкость - образуется муфта. В ходе копуляции сперма попадает в семяприемники партнера, ограниченные от женских гонад.
Таким образом, несмотря на копуляцию, внутреннее оплодотворение у дождевых червей отсутствует. Само оплодотворение произойдет позже.
Муфта постепенно смещается к концу тела, в момент, когда она оказывается напротив отверстий яйцевода, в нее выделяются яйца. Муфта спускается ниже, и оказывается напротив семяприемников, сперматозоиды выходят наружу и оплодотворяют яйца. Муфта смещается к самому концу тела, после чего червь выползает из нее. Она превращается в кокон, в котором в течение 2-4 недель развиваются яйца. После этого времени из кокона выходят достаточно хорошо сформированные молодые организмы.
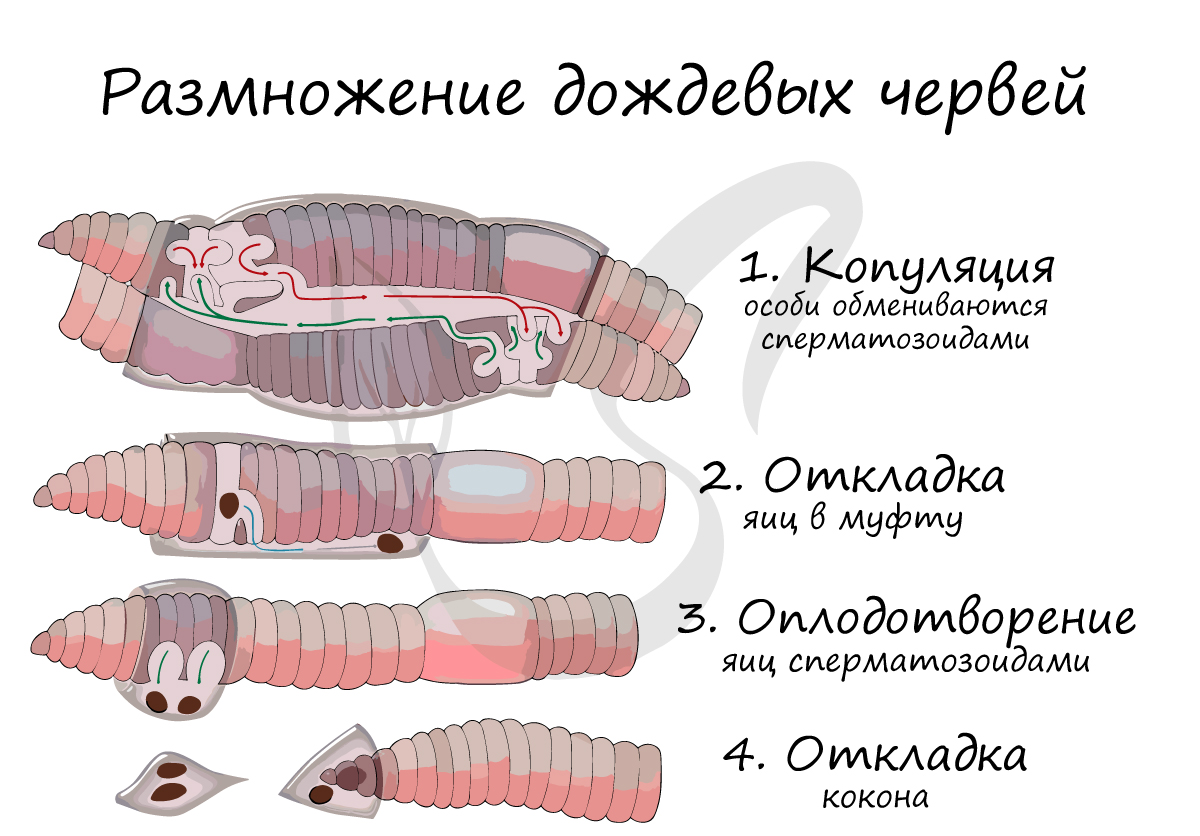
По итогам можно сказать, что оплодотворение у дождевых червей протекает вне организма - по сути, является внешним (наружным). Но нельзя не отметить, что совершается оно под надежной защитой оболочки кокона, в который превращается муфта после сползания ее с тела червя.
Дождевые черви оказывают на почву весьма благоприятный эффект - они способствуют ее перемешиванию, разрыхлению - улучшают ее аэрацию, что очень важно для хорошего развития корней растений.
Уничтожая растительные остатки, содержащие множество гуминовых кислот, они снижают кислотность за счет секрета известковых желез, способствуют ускорению минерализации органических веществ, стимулируют процессы нитрификации - повышают плодородие почвы.

Данная статья написана Беллевичем Юрием Сергеевичем и является его интеллектуальной собственностью. Копирование, распространение (в том числе путем копирования на другие сайты и ресурсы в Интернете) или любое иное использование информации и объектов без предварительного согласия правообладателя преследуется по закону. Для получения материалов статьи и разрешения их использования, обратитесь, пожалуйста, к Беллевичу Юрию.
Общая характеристика
Малощетинковые кольчецы, или олигохеты, представляют собой большую группу кольчецов, включающую около 3100 видов. Они, несомненно, происходят от полихет, но отличаются от них многими существенными особенностями.
Малощетинковые кольчатые черви в подавляющем большинстве живут в почве и на дне пресных водоемов, где они часто зарываются в илистый грунт. Почти в каждом пресном водоеме можно найти червя Tubifex, иногда в огромном количестве. Червь живет в иле, причем сидит зарывшись головным концом в грунт, а задним концом все время совершает колебательные движения.
К почвенным олигохетам относится многочисленная группа дождевых червей, примером которых может служить обыкновенный дождевой червь (Lumbricus terrestris).
Питаются олигохеты преимущественно растительной пищей, главным образом загнивающими частями растений, которые они находят в почве и в иле.
Рассматривая особенности олигохет, мы будем иметь в виду главным образом обыкновенного дождевого червя.
Внешнее строение
Тело олигохет более или менее гомономно сегментировано и часто состоит из очень большого количества сегментов. На переднем конце имеется головная лопасть — простомиум, несущий лишь у некоторых олигохет непарное щупальце. У дождевых червей никаких придатков на простомиуме нет. За простомиумом идет первый ротовой сегмент, или перистомиум, на котором с нижней стороны расположен рот. Далее следуют многочисленные туловищные сегменты. Их число у обычных видов варьирует от 90 до 300, но есть формы с очень большим числом сегментов (до 500-600).
Параподий у олигохет нет, но имеются в небольшом количестве щетинки. У дождевого червя они расположены на каждом сегменте, начиная со второго, четырьмя группами (у некоторых форм поодиночке), по две маленькие щетинки в каждой группе, а всего по восемь щетинок на сегменте. Такое расположение щетинок соответствует их положению на двух параподиях полихет. Это указывает на то, что у олигохет редуцировались параподии, от которых сохранились только щетинки. Параподии мешали бы при передвижении червей в почве или в грунте водоема. Щетинки направлены остриями назад и препятствуют обратному движению тела червя при его передвижении в земляных норках.
На некотором расстоянии от переднего конца тела имеется утолщение покровов, охватывающее несколько сегментов (у дождевого червя с 32-го по 37-й членик). Это — поясок, или clitellum, заключающий много железистых клеток, секрет которых идет на образование кокона.
Тело олигохет заканчивается небольшой анальной лопастью — пигидиумом, на котором открывается заднепроходное отверстие — порошица.
Покровы дождевых червей образованы однослойным кожным эпителием, выделяющим очень тонкую эластичную кутикулу. Кожа дождевых червей богата одноклеточными слизистыми железами, благодаря которым обеспечивается постоянное увлажнение поверхности тела, что особенно важно при роющем (не водном) образе жизни.
Кожно-мускульный мешок олигохет устроен сходно с таковым полихет. Особенностью, свойственной только олигохетам, является наличие: 1) боковых
продольных мышц между дорзальными и вентральными пучками щетинок, что связано с редукцией параподий; 2) спинных пор между сегментами, через которые полостная жидкость выделяется наружу, увлажняя поверхность тела червя.
У олигохет имеется типичный сегментированный целом. Отсутствие диссепиментов наблюдается редко. Спинная брыжейка исчезает, а брюшная не доходит до брюшной стенки тела. Она связывает кишечник с брюшным кровеносным сосудом и брюшной нервной цепочкой. Таким образом, целомическая жидкость может свободно поступать из одного сегмента в другой.
Дождевые черви питаются находящимися в почве перегнивающими остатками растений, а также листьями и травянистыми стеблями, которые они втаскивают в свои норки. Роясь в земле, червь пропускает почву через кишечник, вынося ее по ночам на поверхность земли. У дождевого червя передняя кишка состоит из глотки, пищевода, зоба и мускульного желудка. В пищевод открываются протоки известковых, или морреновых желез, расположенных по его бокам. Выделяемый этими железами углекислый кальций нейтрализует гуминовые кислоты, которыми богата пища дождевых червей. Из пищевода пища поступает в зоб, где накапливается, а затем небольшими порциями переходит в мускульный желудок. Здесь происходит измельчение растительных остатков путем перетирания их между минеральными частицами грунта. Далее идет самый длинный отдел кишечника — средняя кишка. Последняя у большинства олигохет образует на спинной стороне складчатый выступ — тифлозоль, вдающийся внутрь просвета кишки. Таким образом увеличивается переваривающая и всасывающая поверхность кишки. Короткая задняя кишка тифлозоля не имеет, она открывается наружу анальным отверстием.
Основным и часто единственным органом дыхания служит кожа, в которой разветвляется густая сеть капилляров. После дождя вода проходит через верхние слои почвы, где поглощается имеющийся в ней кислород. Такая вода, лишенная кислорода, губительна для дождевых червей. Когда вода заливает норки, черви, спасаясь от нее, вынуждены выползать на поверхность (отсюда — дождевые черви). У некоторых водных олигохет в газообмене участвует задний отдел кишечника.
У олигохет имеется хорошо Развитая замкнутая кровеносная система, сходная с кровеносной системой полихет. Спинной кровеносный сосуд служит основным пропульсаторным органом. Он соединяется с брюшным сосудом кольцевыми, метамерно расположенными сосудами. Кольцевые сосуды переднего конца тела, охватывающие пищевод, самостоятельно пульсируют, за что их называют сердцами. Для олигохет характерно развитие густой сети капилляров в коже, метанефридиях, половых железах и т. д.
Органы выделения олигохет представлены типичными метанефридиями, часто имеющими сложное строение. В экскреторных процессах существенную роль играют также особые, так называемые хлорагогенные клетки. Это видоизмененный перитонеальный эпителий, покрывающий среднюю кишку снаружи. В хлорагогениых клетках накапливаются экскреты, затем клетки выпадают в полостную жидкость и выносятся через воронки метанефридиев.
У олигохет имеются типичные для кольчецов парные надглоточные ганглии, или головной мозг. Он соединяется окологлоточными коннективами с подглоточным ганглием, которым начинается брюшная нервная цепочка. В связи с отсутствием пальп, щупалец и слабым развитием органов чувств головной мозг у олигохет устроен более просто, чем у полихет. Он несколько сдвинут назад и у дождевых червей лежит в III сегменте.
Органы чувств у олигохет развиты значительно слабее, чем у полихет, что объясняется их жизнью в почве или в грунте водоемов.
Наиболее развито осязание. Осязательные чувствительные клетки расположены поодиночке и группами но всей поверхности тела олигохет. Щетинки, по- видимому, также имеют осязательную функцию.
Известно, что дождевые черви, несмотря на отсутствие глаз, очень чувствительны к свету. Восприятие света выполняется светочувствительными ретинальными клетками, расположенными преимущественно на переднем конце тела. У некоторых водных олигохет имеются глаза. Опыты показывают, что дождевые черви чувствуют запахи. Обоняние помогает червю находить необходимую ему пищу. У водных олигохет имеются органы химического чувства в виде ресничных ямок.
Половая система
В отличие от многощетинковых кольчецов олигохеты — гермафродиты. Их половые органы устроены довольно сложно. Рассмотрим их строение на примере полового аппарата дождевого червя.
Половые органы дождевого червя расположены в области 15-го сегментов. Мужские половые органы состоят из двух пар крошечных семенников, лежащих попарно в 10 — 11-м сегментах. Здесь же, вблизи каждого семенника, расположено по одной воронке семявынocящих каналов, идущих назад. Семявыносящие каналы каждой стороны соединяются в семяпроводы, которые открываются двумя половыми отверстиями на брюшной стороне 15-го сегмента.
К семенникам и воронкам примыкают, охватывая их, очень большие семенные пузырьки. Их 3 пары, они расположены в 9, 10 и 12-м сегментах. Не вполне зрелые семенные клетки из семенников поступают в семенные пузырьки, где заканчивается созревание сперматозоидов. Сперматозоиды сохраняются в пузырьках до спаривания червей. Во время спаривания сперма выводится через воронки и семяпроводы наружу.
Женские половые органы состоят из пары небольших яичников, расположенных в 13-м сегменте. В этом же сегменте начинаются воронки яйцеводов, открывающихся половыми отверстиями па 14-м сегменте. Кроме того, к женскому половому аппарату надо отнести две пары мешочков, представляющих собой впячивание эктодермальных покровов на брюшной стороне 9-го и 10-го сегментов. Это — семяприемники, в которые поступает сперма во время спаривания.
Между половозрелыми червями происходит спаривание. Два червя прикладываются друг к другу передними концами так, что поясок одного червя приходится против семяприемников (9-й и 10-й сегменты) другого. При этом пояски выделяют слизистый секрет, который, застывая, образует вокруг пояска каждого из червей муфточки. Оба червя выделяют из мужского полового отверстия (15-й сегмент) некоторое количество спермы. По особому желобку благодаря сокращению мускулатуры червей сперма одного червя стекает в муфточки и попадает в семяприемники другого червя. Взаимно обменявшись спермой, черви расходятся и выползают из слизистых муфточек. Позднее у каждого червя образуется новая слизистая муфточка, содержащая вещества будущего кокона.
При помощи волнообразных движений переднего конца тела червя муфточка постепенно двигается по направлению к головному концу. Когда муфточка проходит в области 14-го сегмента, на котором находятся женские половые отверстия, в нее откладываются яйца. Немного позже из отверстий семяприемников в муфточку выдавливается сперма. Яйца оплодотворяются в муфточке, которая соскальзывает с червя, стягивается на концах и образует замкнутый кокон. Развитие у олигохет происходит без метаморфоза.
У некоторых олигохет, кроме полового размножения, наблюдается также бесполое. Так, у обычных в пресных водах червей Chaetogaster, Aeolosoma и др. наблюдается размножение при помощи так называемой паратомии. Оно состоит в том, что в каком-либо месте тела червя обособляется участок, в котором сначала развивается головной конец для задней части червя и хвостовой для передней, а после этого происходит деление червя на две особи. При этом еще до разъединения дочерних особей возникают новые участки, в которых происходит тот же процесс, в результате чего образуются целые цепочки дочерних червей.
У других олигохет наблюдается более простой способ бесполого размножения, при котором тело червя сначала распадается на две или несколько частей, а после этого отделившиеся части червя развивают передний и задний концы тела. Такое бесполое размножение называется архитомией. У некоторых олигохет такой способ размножения является обычным, например у некоторых видов пресноводных олигохет (Lumbriculus variegatus и др.).
Понятно, что в тех случаях, когда черви размножаются путем архитомии, размножение сопровождается регенерацией недостающих частей тела. Большинство олигохет обладают сильно развитой способностью к регенерации, причем у форм, способных к бесполому размножению, она более резко выражена.
У некоторых олигохет даже несколько вырезанных члеников регенерируют в целого червя, а у упомянутого выше Lumbriculus variegatus к регенерации способны даже отдельные членики.
Важнейшие представители класса малощетинковых кольчецов и их практическое значение
Олигохеты широко распространены по всему земному шару. Насчитывается свыше 3400 видов, большинство которых составляют разнообразные почвенные олигохеты. Многочисленны и пресноводные формы малощетинковых кольчецов.
В озерах и прудах встречается много олигохет, в том числе виды рода Tubifex, Lumbriculus variegatus, различные виды родов Nais, Stylaria, Aeolosoma (весьма обычные в аквариумах), а также пресноводные представители семейства Enchytraeidae и др.
Почвенные олигохеты представляют собой группу, особенно богатую видами. При этом различные почвы характеризуются наличием определенных видов почвенных олигохет. ВРоссии из почвенных олигохет распространены представители двух семейств — Enchytraeidae и Lumbricidae, причем семейство Lumbricidae представлено почти 50 различными видами, распространенными в различных почвах и в разных районах России.
Среди почвенных олигохет есть очень крупные формы. Таков, например, Megascolides australis, живущий в Австралии и достигающий длины свыше 2 м.
Большинство олигохет, безусловно, полезные животные. Пропуская через кишечник огромное количество ила, пресноводные олигохеты являются существенным фактором, влияющим на грунт водоема. Так как пресноводные олигохеты питаются растительным детритом, они играют огромную роль в очищении водоемов от гниющих остатков растений.
Особенно большое значение в жизни почвы имеют различные виды дождевых червей. Влияние их на почву заключается в том, что они разрыхляют ее на большую глубину, чем содействуют проникновению воздуха (аэрацпя почвы) и влаги, что способствует деятельности почвенных микробов, а также проникновению корней растений в глубокие слои почвы и укреплению их. Кроме того, дождевые черви перепахивают почву. Пропуская почву через кишечник, они выносят постепенно почву из более глубоких слоев на поверхность. Ч. Дарвин, написавший прекрасную работу о роли дождевых червей, указывает, что за один год па площади 1 м 2 дождевыми червями было вынесено па поверхность до 4 кг почвы (сухой вес). Затаскивая в свои норы остатки растений, черви погребают их, чем способствуют накоплению в почве органических веществ и образованию перегноя. По этой же причине черви содействуют погружению камней и древних сооружений под почву. Так, некоторые остатки сооружений древнего Рима оказались под землей благодаря работе дождевых червей.
Дождевые черви используются также в качестве ценного корма для рыб и домашней птицы. В связи с этим в последнее время все шире развертывается искусственное разведение червей.
Класс малощетинковые - полное описание
Малощетинковые черви – обитатели пресных вод и почвы, единично встречающихся в морях. Известно более 5000 видов. Отличительными особенностями внешнего строения малощетинковых червей являются гомономная сегментация тела, отсутствие параподий, наличие железистого пояска в передней трети тела у половозрелых особей. Головной отдел у них не выражен. Головная лопасть, как правило, лишена глаз и придатков. На анальной лопасти (пигидиуме) также нет никаких придатков. По бокам тела расположены щетинки, обычно по четыре пары пучков на каждом сегменте. Это рудименты параподии. Такое упрощение внешнего строения связано с адаптациями к роющему образу жизни. у малощетинковых червей наблюдается конвергентное сходство с роющими полихетами. Это подтверждает причину их морфоэкологического сходства в связи с развитием сходного, роющего образа жизни. Наиболее знакомыми нам олигохетами являются обитающие в почве дождевые черви. Их тело достигает нескольких сантиметров, самые крупные среди них – до 3 м. (в Австралии). В почве обычны и мелкие беловатые кольчатые черви – энхитреиды (5 – 10 мм). Дождевые черви и энхитреиды питаются растительными остатками в почве и играют важную роль в почвообразовании. В пресных водоемах нередко можно видеть олигохет с длинными щетинками или живущих в вертикальных трубках и образующих плотные поселения на дне. Они питаются взвешенными органическими остатками и являются полезными фильтраторами, играющими существенную роль в очистке вод.

Кольчатые черви. Фото: Shanegenziuk
Общая морфофизиологическая характеристика. Длина тела варьирует от нескольких миллиметров до 3 м. Тело длинное, червеобразное, сегментированное. Число сегментов колеблется от 5 – 6 до 600. На первом сегменте тела после головной лопасти находится рот. На анальной лопасти располагается анальное отверстие. Движутся олигохеты сокращая мускулатуру тела. При рытье червь передним концом тела раздвигает почву, опираясь на многочисленные щетинки. Щетинки упираются в стенки прорытого хода, поэтому дождевого червя трудно вытащить из норки.
Кожно-мускульный мешок. Тело покрыто слоем кожного эпителия, часто с большим количеством железистых клеток. Кожа выделяет тонкую кутикулу. Обильное выделение слизи защищает кожу дождевых червей от механических повреждений и высыхания. Под кожей, как и у полихет, у них залегают кольцевые и продольные мышцы, изнутри выстланные целомическим эпителием.
Пищеварительная система. Кишечник проходит по всей длине тела. В переднем отделе кишечника дождевого червя обособлены ротовая полость, мускулистая глотка, относительно узкий пищевод, зоб и желудок. В стенках пищевода имеются три пары известковых желез, секреты которых нейтрализуют гуминовые кислоты в пище дождевых червей. Из желудка пища поступает в среднюю кишку, где происходит всасывание питательных веществ. Непереваренные остатки пищи и минеральные частицы почвы поступают в короткую заднюю кишку и удаляются через анальное отверстие наружу. В средней кишке дорсально расположена внутренняя продольная складка – тифлозоль, свешивающаяся в просвет кишки и увеличивающая всасывательную поверхность кишечника.
Выделительная система представлена метанефридиями. Способность метанефридиев экономить влагу в организме путем реабсорбции воды обеспечивает приспособленность олигохет к жизни на суше. Твердые экскреты накапливаются в хлорагогенных клетках целомического эпителия. Частично эти клетки, наполненные экскретами, удаляются через воронки нефридиев или через особые поры в стенке тела.
Нервная система представлена, как у всех кольчецов, парой надглоточных ганглиев (мозгом) и брюшной нервной цепочкой.
Органы чувств у малощетинковых червей развиты слабее, чем у большинства многощетинковых, в связи с роющим образом жизни. Глаза, как правило, отсутствуют. В коже олигохет имеются многочисленные чувствующие клетки: светочувствительные, осязательные и др. дождевые черви чутко реагируют на факторы света, влажности и температуры. Этим объясняются их вертикальные миграции в почве в течении суток и по сезонам.
Половая система олигохет гермафродитная. Гермафродитные особи олигохет однотипные, в отличии от половозрелых особей полихет с половым диморфизмом. Гермафродитизм в животном мире – это приспособление к увеличению плодовитости, так как все 100% особей в популяции могут откладывать яйца. Рассмотрим строение половой системы на примере дождевого червя. Половые железы у олигохет сосредоточены в передних сегментах тела. Семенники (две пары) расположены в 10-м и 11-м сегментах тела и прикрыты тремя парами семенных мешков. В семенных мешках накапливается сперма, вытекающая из семенников. Здесь происходит созревание сперматозоидов. Спермии поступают в мерцательные воронки семяпроводов. Семяпроводы сливаются попарно левой и правой сторонами тела, и образуются два продольных канала, открывающихся парными мужскими половыми отверстиями на 15-м сегменте тела. Женская половая система представлена парой яичников, расположенных на 13-м сегменте, парой яйцеводов с воронками, открывающихся половыми отверстиями на 14-м сегменте. В 13-м сегменте диссипименты образуют яйцевые мешки, прикрывающие яичники и воронки яйцеводов. К женской половой системе относятся еще особые кожные впячивания на 9-м и 10-м сегментах – две пары семяприемников с отверстиями на брюшной стороне тела.
Развитие у олигохет протекает без метаморфоза, т. е. без личиночных стадий. Из яйцевого кокона вылупляются маленькие червячки, похожие на взрослых. Такое прямое развитие без метаморфоза возникло у олигохет в связи с переходом к жизни на суше или к обитанию в пресных водоемах, которые нередко пересыхают. Эмбриональное развитие зародыша олигохет протекает, как и у большинства полихет, по спиральному типу дробления и с телобластической закладкой мезодермы.
Бесполое размножение известно в некоторых семействах пресноводных олигохет. При этом происходит поперечное деление червя на несколько фрагментов, из которых потом развиваются целые особи, или путем дифференциации червя на цепочку из коротких дочерних особей. В дальнейшем эта цепочка распадается. У дождевых червей крайне редко наблюдается бесполое размножение, зато хорошо выражена способность к регенерации. Перерезанный червь, как правило, не погибает, а каждая его часть восстанавливает недостающие концы. Наиболее легко червь восстанавливает задний конец тела. Головной конец тела восстанавливается редко и с трудом.
Д О К Л А Д
ПО БИОЛОГИИ
средней школы N 8
Снытко Николая
Кольчатые черви ведут свое начало от примитивных червей с нерасчлененным телом. Наиболее древними из кольчатых червей являются морские многощетинковые черви. От них при переходе к пресноводному и наземному образу жизни произошли малощетинковые, а от них пиявки.
Класс Малощетинковые
Его представители — в основном обитатели почвы, но известны и пресноводные формы. Строение малощетинковых в значительной степени определено почвенным образом жизни, благодаря чему организация червей упростилась. Головной отдел имеет простое строение, лишен органов чувств. Параподии (органы передвижения) отсутствуют, щетинки сохранились в ограниченном числе. Длина тела от долей мм до 2,5 м (некоторые тропические дождевые черви). Имеется вторичная полость тела - целом. Сегментация тела хорошо выражена внутри и снаружи.
Большинство малощетинковых червей питается растительным детритом, который поглощают с грунтом.
(Детрит - мертвое органическое или частично минерализованное вещество, взвешенное в толще воды в виде частиц размером от нескольких микронов до нескольких сантиметров. Детрит образуется из отмерших растений и животных или их выделений, причём часто органическое вещество адсорбируется на поверхности минеральных взвесей. В детрите всегда живут микроорганизмы. Детрит - основная пища многих водных животных.)
Размножение
Малощетинковые черви – гермафродиты. Размножаются посредством спаривания. Яйца оплодотворяются одной из спаривающихся особей и откладываются в специфическом коконе, состоящем из слизи, выделяемой железистыми клетками (т.н. пояска на теле).
Почвенные кольчатые черви - полезные животные. Еще Ч. Дарвин отметил их значение для плодородия почвы. Затаскивая в норы опавшие листья, они обогащают почву перегноем, а, прокладывая в почве ходы, рыхлят ее и способствуют проникновению воздуха, воды к корням растений.
Пресноводные малощетинковые играют значительную роль в питании донных рыб.
Известные виды
Описано примерно 3000 видов. Некоторые из них:
Наиболее известные группы:
Дождевые черви - типичные представители этого класса. Дождевые черви живут во влажной, богатой перегноем почве.
Дождевой червь имеет сильно вытянутое, почти круглое в поперечном разрезе тело, способное сокращаться и удлиняться, длинной до 30 см. Кольцевидные перетяжки - главный признак всех кольчатых червей - разделяют тело дождевого червя на 100-180 сегментов. В передней части тела находится утолщение - поясок (его клетки функционируют в период полового размножения и откладки яиц). На брюшной стороне червя развиты тонкие упругие и короткие щетинки. Цепляясь ими за неровности почвы, червь с помощью мышц мощного кожно-мускульного мешка продвигается вперед. Наличие редких щетинок - характерная особенность всего класса малощетинковых.
Дождевой червь имеет красновато-коричневую окраску, при этом его брюшная сторона светлее спинной.
Характерной особенностью внутреннего строения является то, что у дождевых червей хорошо развиты настоящие ткани. Снаружи тело червя покрыто слоем эктодермы, клетки которой образуют покровную ткань. Кожный эпителий богат слизистыми железистыми клетками. Хорошо развитая мускулатура состоит из слоя продольных и кольцевых мышц. При сокращении кольцевых мышц тело червя вытягивается, при сокращении продольных утолщается и раздвигает частички почвы. Чередование действия мышц имеет особое значение при рытье норок.
Системы органов
У дождевого червя органы располагаются в определенной последовательности, связаны между собой, образуя в целом систему.
Пищеварительная система. Дождевой червь питается перегнившими растительными остатками, которые он заглатывает вместе с землей. Он может затаскивать в свою норку опавшие листья и проглатывать их по частям. Пища поступает сначала в переднюю кишку, состоящую из глотки, пищевода, зоба и желудка. От желудка до конца тела тянется кишка. В кишке под действием пищеварительных соков измельченная пища переваривается, а питательные вещества всасываются. Не переваренные остатки и земля выбрасываются наружу через анальное отверстие. Рот, глотка, пищевод, зоб, желудок и кишка - органы, которые в своей совокупности образуют пищеварительную систему.
Выделительная система. Жидкие вредные продукты жизнедеятельности накапливаются в полости тела. В каждом членике дождевого червя расположена пара органов выделения - тонких, петлеобразно извитых трубочек. Одним концом трубочка сообщается с полостью тела, а другим открывается наружу. Через эти трубочки вредные жидкие вещества выводятся из организма дождевого червя.
Нервная система. В каждом членике червя на брюшной стороне находится небольшой нервный узел. Все узлы соединены между собой, образуя брюшную нервную цепочку. В передней части этой цепочки, под глоткой, находится большой подглоточный узел, а над глоткой - самый крупный надглоточный узел. Надглоточный и подглоточный узлы соединены нервными перемычками, окружающими глотку с боков. Оба нервных узла и перемычки образуют окологлоточное нервное кольцо. От всех нервных узлов в различные части тела червя отходят нервы.
Специальных органов чувств у червя нет, но он ощущает прикосновение к его телу, вкус пищи, отличает свет от темноты. Раздражения воспринимаются чувствительными клетками кожи, связанными при помощи нервов с нервными узлами. Особенно много чувствительных клеток на переднем конце тела, от них нервы подходят к узлам окологлоточного нервного кольца. Рефлексы дождевого червя значительно сложнее, чем рефлексы кишечнополостных и плоских червей. В отличие от них дождевой червь при соприкосновении с иглой может извиваться в ту или другую сторону, сокращать только передний или только задний конец тела. Это зависит от более сложного строения его нервной системы.
Для дождевых червей характерна высокая способность к регенерации.
Размножение происходит только половым путем. Дождевые черви — гермафродиты. Оплодотворение перекрестное.
Трубочник обыкновенный
Трубочник обыкновенный - водные, донные черви, красного цвета, длиной 40-60 мм. Обитают они обычно на илистых грунтах, образуя на дне водоемов скопления из множества - сотен и тысяч - особей, похожие на своеобразные красные "подушки". Большая часть тела трубочника погружена в грунт, наружу высовывается лишь его задний конец. Вокруг него образуется короткая, направленная вверх трубочка, состоящая из склеенных слизью частичек ила. При малейшей тревоге черви сразу прячутся в трубочки, но вскоре опять высовываются наружу. Трубочники встречаются даже в сильно загрязненных водоемах - в сточных канавах, прудах и реках в черте города.
Пиявки
Срок жизни пиявок – несколько лет. Все они гермафродиты. Яйца откладываются в коконах, личиночной стадии нет. Большинство пиявок сосёт кровь у различных животных, в том числе человека. Пиявки прокалывают кожные покровы хоботком или зубчиками на челюстях, а специальное вещество – гирудин – препятствует свёртыванию крови. Высасывание крови из одной жертвы может продолжаться месяцы. В кишечнике кровь не портится очень долго: пиявки могут жить без пищи даже два года. Некоторые пиявки – хищники, целиком проглатывающие добычу.
Пиявки обитают в пресных водоёмах, встречаются также в морях и почве. Пиявки служат кормом для рыб. Медицинская пиявка используется человеком в лечебных целях.
Читайте также: