
Увеличивает скорость проведения нервного импульса
В 1830 г. один из крупнейших физиологов XIX века Иоганн Мюллер заявил, что скорость распространения ПД измерить невозможно. По его мнению, поскольку ПД – это электрический импульс, он должен проводиться со скоростью, примерно равной скорости света (3–10 10 см/с); учитывая небольшие размеры биологических объектов, даже с помощью лучших инструментов того времени измерить такую скорость было невозможно.
Спустя 15 лет один из студентов Мюллера Герман фон Гельмгольц с помощью простого и изящного эксперимента, который легко воспроизвести на студенческом лабораторном практикуме (рис. 6–8), измерил скорость распространения импульсов в нерве лягушки. В своих опытах Гельмгольц раздражал нерв в двух участках, отстоящих друг от друга на 3 см, и измерял время от момента подачи стимула до максимума мышечного сокращения. Предположим, что при раздражении дистального (расположенного ближе к мышцам) участка это время уменьшается на 1 мс. Тогда скорость распространения импульсов V равна
| V | = | d | / | t | = | 3 см | / | 1 мс | = | · | 10 3 | см/с |
Эта величина оказалась на семь порядков меньше, чем скорость распространения электрического тока в медном проводнике или в растворе электролита. Отсюда Гельмгольц сделал совершенно правильный вывод, что проведение нервного импульса –это более сложный процесс, чем простое продольное распространение тока в нервном волокне.
Скорость распространения импульсов в различных аксонах варьирует от 120 м/с (в некоторых крупных волокнах) до нескольких сантиметров в секунду (в очень тонких аксонах). Эти различия между скоростью проведения в разных волокнах иллюстрируют табл. 6–1 и рис. 6–9.
Скорость распространения импульса в значительной степени зависит от того, как быстро участок мембраны, расположенный на определенном расстоянии от места подачи стимула, деполяризуется местными токами до порогового уровня. Чем выше постоянная длины волокна, тем дальше могут распространяться эти токи, тем быстрее происходит деполяризация мембраны впереди от места возбуждения и, следовательно, тем выше скорость распространения импульса. Влияние постоянной длины на эту скорость можно продемонстрировать, если поместить аксон в масло или в воздух. При этом на поверхности аксона остается лишь тонкая пленка солевого раствора, и постоянная длины уменьшается из–за увеличения наружного продольного сопротивления [в уравнении (6–2) –r0]. В этих условиях скорость проведения возбуждения будет ниже чем при погружении аксона в солевой раствор.
Таблица 6–1. Классификация нервных волокон лягушки по их диаметру и скорости проведения возбуждения (Erlanger, Gasser, 1937)
| Группа волокон | Диаметр, мкм | Скорость, м/с |
| A α | 18,5 | |
| β | 14,0 | |
| γ | 11,0 | |
| B | — | 4,2 |
| C | 2,5 | 0,4 –0,5 |
В процессе эволюции живые организмы выработали два способа увеличения постоянной длины аксона и тем самым–скорости распространения импульса. Один из них (типичным примером могут быть гигантские аксоны кальмаров, членистоногих кольчатых червей, костистых рыб) – это увеличение диаметра аксона, т. е. уменьшение внутреннего продольного сопротивления [в уравнении (6–2) – ri] Подробнее этот вопрос рассмотрен в дополнении 6–2. Гигантские аксоны развились в процессе эволюции у некоторых видов животных для того, чтобы обеспечивать быструю синхронную активацию двигательных рефлексов, например движений мантии у кальмара и рефлекса отдергивания либо избегания у некоторых членистоногих (раков, тараканов) и кольчатых червей (например, земляных).
Сальтаторное проведение
Второй способ увеличить скорость проведения нервных импульсов, реализовавшийся только у позвоночных животных, состоит в изоляции участков аксона с помощью миелиновой оболочки. При этом постоянная длины соответствующих участков значительно увеличивается, и тем самым существенно облегчается проведение тока в продольном направлении. По мере развития животного миелин откладывается вокруг периферических и центральных аксонов глиальными клетками, расположенными вблизи этих аксонов. В результате вокруг волокон образуется плотная многослойная оболочка из клеточных мембран. К клеткам, синтезирующим миелин, относятся шванновские клетки (в области периферических нервов) и олигодендроциты (рис. 6–10) (в ЦНС). На поперечных срезах миелиновой оболочки видны периодически повторяющиеся промежутки в 12 нм, образующиеся в результате наслоения мембран глиальных клеток. С образованием каждого нового слоя поперечное сопротивление оболочки увеличивается. Поскольку слоев в этой оболочке много, ее емкость гораздо ниже, чем у одиночной мембраны. Многослойная миелиновая оболочка периодически прерывается (так называемые перехваты Ранвье>, и на этих небольших участках возбудимая мембрана аксона контактирует с внеклеточной средой. Между перехватами Ранвье миелиновая оболочка тесно прилегает к мембране аксона, практически вытесняя внеклеточную среду. Кроме того, участки мембраны аксона между перехватами Ранвье, по–видимому, не содержат натриевых каналов.
Благодаря изолирующим свойствам миелиновой оболочки постоянная длины аксона резко возрастает: наличие этой оболочки оказывает такой же эффект, как и увеличение rм [уравнение (6–2)]. Из–за высокого сопротивления миелиновой оболочки местные токи, текущие впереди от волны возбуждения, выходят из аксона почти исключительно в области перехватов Ранвье. Кроме того, поскольку емкость толстой миелиновой оболочки мала, на перезарядку этой емкости в участках между перехватами расходуется лишь очень небольшой ток. Благодаря этим особенностям ПД, возникающий в каком–либо перехвате, электротонически деполяризует лишь мембрану, расположенную в области следующего перехвата, и поэтому импульсы в таких аксонах не распространяются по всей их длине, как в немиелинизированных нервных волокнах (например, в аксоне кальмара). Они возникают лишь в небольших участках мембраны –перехватах Ранвье. Все это обусловливает сальтаторное (скачкообразное) проведение, при котором импульсы распространяются прерывисто от перехвата к перехвату (рис. 6–11). Скорость распространения. ПД при этом резко увеличивается, поскольку электротоническое проведение местных токов между перехватами осуществляется очень быстро. Таким образом, у позвоночных животных Природа решила проблему быстрого распространения нервных импульсов, не прибегая к созданию таких громоздких структур, как гигантские аксоны.
Нервная система человека выступает своеобразным координатором в нашем организме. Она передаёт команды от мозга мускулатуре, органам, тканям и обрабатывает сигналы, идущие от них. В качестве своеобразного носителя данных используется нервный импульс. Что он собой представляет? С какой скоростью работает? На эти, а также на ряд других вопросов можно будет найти ответ в этой статье.
Чем является нервный импульс?
Исследование строения и работы
Впервые прохождение нервного импульса было продемонстрировано немецкими учеными Э. Герингом и Г. Гельмгольцем на примере лягушки. Тогда же и было установлено, что биоэлектрический сигнал распространяется с указанной ранее скоростью. Вообще, такое является возможным благодаря особенному построению нервных волокон. В некотором роде они напоминают электрический кабель. Так, если проводить параллели с ним, то проводниками являются аксоны, а изоляторами – их миелиновые оболочки (они являют собой мембрану шванновской клетки, которая намотана в несколько слоев). Причем скорость нервного импульса зависит в первую очередь от диаметра волокон. Вторым по важности считается качество электрической изоляции. Кстати, в качестве материала организмом используется липопротеид миелин, который обладает свойствами диэлектрика. При прочих равных условиях, чем больше будет его слой, тем быстрее будут проходить нервные импульсы. Даже на данный момент нельзя сказать, что эта система полноценно исследована. Многое, что относится к нервам и импульсам, ещё остаётся загадкой и предметом исследования.
Особенности строения и функционирования
Где они создаются?
Типы клеток
- Рецепторные (чувствительные). Ими кодируются и превращаются в нервные импульсы все температурные, химические, звуковые, механические и световые раздражители.
- Вставочные (также называются кондукторными или замыкательными). Они служат для того, чтобы перерабатывать и переключать импульсы. Наибольшее их число находится в головном и спинном мозге человека.
- Эффекторные (двигательные). Они получают команды от центральной нервной системы на то, чтобы были совершены определённые действия (при ярком солнце закрыть рукой глаза и так далее).
Каждый нейрон имеет тело клетки и отросток. Путь нервного импульса по телу начинается именно с последнего. Отростки бывают двух типов:
- Дендриты. На них возложена функция восприятия раздражения расположенных на них рецепторов.
- Аксоны. Благодаря им нервные импульсы передаются от клеток к рабочему органу.
Интересный аспект деятельности
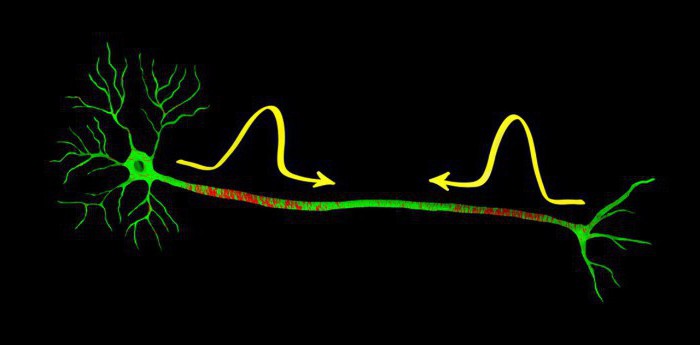
О потенциале действия
Как всё работает в мозгу?
Работа нейромедиаторов
Когда они передают нервные импульсы, то существует несколько вариантов, что произойдёт с ними:
- Они будут диффундированы.
- Подвергнутся химическому расщеплению.
- Вернутся назад в свои пузырьки (это называется обратным захватом).
В конце 20-го века сделали поразительное открытие. Ученые узнали, что лекарства, что влияют на нейромедиаторы (а также их выброс и обратный захват), могут изменять психическое состояние человека коренным образом. Так, к примеру, ряд антидепрессантов вроде "Прозака" блокируют обратный захват серотонина. Есть определённые причины считать, что в болезни Паркинсона виноват дефицит в головном мозге нейромедиатора дофамина.
Если кратко, то они могут работать с тысячами нейромедиаторов, которые посылаются их соседями. Детали относительно обработки и интеграции данного типа импульсов нам почти не известны. Хотя над этим работает много исследовательских групп. На данный момент получилось узнать, что все полученные импульсы интегрируются, а нейрон выносит решение – необходимо ли поддерживать потенциал действия и передавать их дальше. На этом фундаментальном процессе базируется функционирование головного мозга человека. Ну что ж, тогда это неудивительно, что мы не знаем ответа на эту загадку.
Некоторые теоретические особенности
Где же создаются нервные импульсы?
Откуда они начинают свой путь? Ответ на этот вопрос может дать любой студент, который прилежно изучал физиологию возбуждения. Есть четыре варианта:
- Рецепторное окончание дендрита. Если оно есть (что не факт), то возможным является наличие адекватного раздражителя, что создаст сначала генераторный потенциал, а потом уже и нервный импульс. Подобным образом работают болевые рецепторы.
- Мембрана возбуждающего синапса. Как правило, такое возможно только при наличии сильного раздражения или их суммирования.
- Триггерная зона дентрида. В этом случае локальные возбуждающие постсинаптические потенциалы формируются как ответ на раздражитель. Если первый перехват Ранвье миелинизирован, то они на нём суммируются. Благодаря наличию там участка мембраны, которая обладает повышенной чувствительностью, здесь возникает нервный импульс.
- Аксонный холмик. Так называют место, где начинается аксон. Холмик – это наиболее частый создать импульсов на нейроне. Во всех остальных местах, которые рассматривались ранее, их возникновение гораздо менее вероятное. Это происходит из-за того, что здесь мембрана имеет повышенную чувствительность, а также пониженный критический уровень деполяризации. Поэтому, когда начинается суммирование многочисленных возбуждающих постсинаптических потенциалов, то раньше всего на них реагирует холмик.
Пример распространяющегося возбуждения
Вспомните сводки из новостей прошлого лета (также это скоро можно будет услышать опять). Пожар распространяется! При этом деревья и кустарники, которые горят, остаются на своих местах. А вот фронт огня идёт всё дальше от места, где был очаг возгорания. Аналогичным образом работает нервная система.
Часто бывает необходимо успокоить начавшееся возбуждение нервной системы. Но это не так легко сделать, как и в случае с огнем. Для этого совершают искусственное вмешательство в работу нейрона (в лечебных целях) или используют различные физиологические средства. Это можно сравнить с заливанием пожара водой.
Фиксация напряжения на мембране.
Внутриклеточное отведение.
Экспертный метод
Аукционный метод.
На основе закрытых торгов.
Рыночные методы.
На основе уровня текущих цен.При использовании этого метода цены устанавливаются на основе цен, принятых на данном рынке. Здесь применяются цены, которые сохраняются на стабильном уровне в отношении определенных товаров в течение длительного времени на конкретном рыночном пространстве.
Один электрод погружают в межклеточную жидкость, другой (микроэлектрод) - вводится в цитоплазму. Между ними – измерительный прибор.
Микроэлектрод представляет собой полую трубку, кончик которой оттянут до диаметра в доли микрона, а пипетка наполнена хлоридом калия. При введении микроэлектрода мембрана плотно охватывает кончик, и повреждения клетки почти не происходит.
Для создания потенциала действия в эксперименте клетка стимулируется надпороговыми токами, т.е. ещё одна пара электродов связана с источником тока. На микроэлектрод подаётся положительный заряд.
С их помощью можно регистрировать биопотенциалы как крупных, так и мелких клеток, а также биопотенциалы ядер. Но наиболее удобным, классическим объектом исследований, являются биопотенциалы крупных клеток. Например,
Nitella ПП 120 мВ (120 * 10 3 В)
Гигантский аксон кальмара ПП 60мВ
Клетки миокарда человека ПП 90 мВ
В определённый момент развитие потенциала действия искусственно прерывается с помощью специальных электронных схем.
При этом фиксируется значение мембранного потенциала и величины ионных потоков через мембрану в данный момент, следовательно, есть возможность их измерения.
Перфузия – замена оксоплазмы искусственными растворами различного ионного состава. Таким образом, можно определить роль конкретного иона в генерации биопотенциалов.
Проведение возбуждения по нервным волокнам.
Роль потенциала действия в жизнедеятельности.
Раздражимость – способность живых клеток под влиянием раздражителей (определённых факторов внешней или внутренней среды) переходить из состояния покоя в состояние активности. При этом всегда меняется электрическое состояние мембраны.
Возбудимость – способность специализированных возбудимых клеток в ответ на действие раздражителя генерировать особую форму колебания мембранного потенциала – потенциал действия.
В принципе возможно несколько видов ответов возбудимых клеток на раздражение, в частности – локальный ответ и потенциал действия.
Потенциал действия возникает, если действует пороговый или надпороговый раздражитель. Он вызывает уменьшение мембранного потенциала до критического уровня. Тогда происходит открытие дополнительных натриевых каналов, резкое увеличение натриевой проницаемости и развитие процесса по механизму положительной обратной связи.
Локальный ответ возникает, если действует подпороговый раздражитель, составляющий 50-70% от порогового. Деполяризация мембраны при этом меньше критической, наступает лишь кратковременное, небольшое увеличение натриевой проницаемости, механизм положительной обратной связи не включается, и потенциал быстро возвращается к исходному состоянию.
В процессе развития потенциала действия возбудимость меняется.
Снижение возбудимости – относительная рефрактерность.
Полная утрата возбудимости – абсолютная рефрактерность.
По мере приближения к уровню критической деполяризации возбудимость повышается, так как для достижения этого уровня и развития потенциала действия становится достаточно и небольшого изменения мембранного потенциала. Именно так меняется возбудимость в начале фазы деполяризации, а также при локальном ответе клетки на раздражение.
При удалении мембранного потенциала от критической точки возбудимость снижается. На пике фазы деполяризации, когда клетка уже не может реагировать на раздражение открытием дополнительных натриевых каналов, наступает состояние абсолютной рефрактерности.
Во время фазы гиперполяризации возбудимость снова снижена.
Возбуждение – ответ специализированных клеток на действие пороговых и надпороговых раздражителей – это сложный комплекс физико-химических и физиологических изменений, в основе которого лежит потенциал действия.
Результат возбуждения зависит от того, в какой ткани оно развивалось (где возник потенциал действия).
К специализированным возбудимым тканям относятся: нервная, мышечная, железистая.
Потенциалы действия обеспечивают проведение возбуждения по нервным волокнам и инициируют процессы сокращения мышечных и секреции железистых клеток.
Потенциал действия, возникающий в нервном волокне – нервный импульс.
Об аксонах.
Аксоны (нервные волокна) – длинные отростки нервных клеток (нейронов).
Афферентные пути – от органов чувств к ЦНС
Эфферентные пути – от ЦНС к мышцам.
Протяжённость – метры.
Диаметр в среднем от 1 до 100 мкм (у гигантского аксона кальмара – до 1 мм).
По наличию или отсутствию миелиновой оболочки различают аксоны: миелинизированные (миелиновые, мякотные) – есть миелиновая оболочка, немиелинизированные (амиелиновые, безмякотные) – не имеют миелиновые оболочки.
Миелиновая оболочка – окружающая аксон дополнительная многослойная (до 250 слоёв) мембрана, образующаяся при внедрении аксона в шванновскую клетку (леммоцит, олигодендроцит), и многократном наматывании мембраны этой клетки на аксон.
Миелин – очень хороший изолятор.
Через каждые 1-2 мм миелиновая оболочка прерывается перехватами Ранвье, протяжённостью около 1 мкм каждый.
Только в области перехватов возбудимая мембрана контактирует с внешней средой.
Кабельная теория проведения.
Аксон по ряду свойств подобен кабелю: это полая трубка, внутренне содержимое – аксоплазма – проводник (как и межклеточная жидкость), стенка – мембрана – изолятор.
Механизм проведения возбуждения (распространения нервного импульса) включает 2 ступени:
1. Возникновение локальных токов и распространение волны деполяризации вдоль волокна.
2. Формирование потенциалов действия на новых участках волокна.
Локальные токи циркулируют между возбужденным и невозбуждённым участками нервного волокна ввиду разной полярности мембраны на этих участках.
Внутри клетки они текут от возбуждённого участка к невозбуждённому. Снаружи – наоборот.
Локальный ток вызывает сдвиг мембранного потенциала соседнего участка, и начинается распространение волны деполяризации по волокну, как тока по кабелю.
Когда деполяризация очередного участка достигает критической величины, происходит открытие дополнительных натриевых, потом калиевых каналов, возникновение потенциала действия.
В разных участках волокна потенциал действия формируется независимыми ионными потоками, перпендикулярными к направлению распространения.
При этом на каждом участке происходит энергетическая подпитка процесса, так как градиенты ионов, идущих по каналам, создаются насосами, работа которых обеспечивается энергией гидролиза АТФ.
Роль локальных токов – лишь инициация процесса путём деполяризации всё новых участков мембраны до критического уровня.
Благодаря энергетической подпитке нервный импульс распространяется вдоль волокна без затухания (с неизменной амплитудой).
Направление и скорость проведения.
Одностороннее проведение нервного импульса обеспечивают:
наличие в нервной системе синапсов с односторонним проведением, свойство рефрактерности нервного волокна, что делает невозможным обратный ход возбуждения.
Скорость проведения тем выше, чем более выражены кабельные свойства волокна. Для их оценки применяют константу длины нервного волокна:
D – диаметр волокна
bm – толщина мембраны
Физический смысл константы: она численно равна расстоянию, на котором подпороговый потенциал уменьшился бы в e раз. С увеличением константы длины нервного волокна увеличивается и скорость проведения.
Беспозвоночные: увеличение диаметра волокна.
Позвоночные: увеличение толщины и удельного сопротивления мембраны путём приобретения миелиновой оболочки, следовательно, происходит качественное изменение способа проведения.
Непрерывное и сальтаторное проведение.
Проведение возбуждения по амиелиновым волокнам – непрерывное: в своё время, последовательно, один за другим каждый участок волокна переходит в состояние возбуждения.
Проведение по миелиновым волокнам – сальтаторное (скачкообразное).
Возбудимая мембрана мякотного аксона обнажена только перехватами Ранвье, только там открываются натриевые и калиевые каналы, только в них происходит генерация потенциала действия.
По миелинизированному волокну импульс распространяется на порядок быстрее. Это обеспечивает позвоночным эволюционное преимущество.
В основе активного транспорта лежит сопряжение противоградиентных потоков вещества с гидролизом АТФ.
Молекулярный механизм, локализованный в мембране и способный транспортировать вещества за счёт энергии гидролиза АТФ, называется биологическим насосом.
Молекулярная организация и этапы работы Калий - Натриевого насоса.
В основе устройства лежит фермент: калий, натрий – активируемая АТФ-аза.
Нам важно ваше мнение! Был ли полезен опубликованный материал? Да | Нет
Нервный импульс (лат. nervus нерв; лат. impulsus удар, толчок) — волна возбуждения, распространяющаяся по нервному волокну; единица распространяющегося возбуждения.
Нервный импульс обеспечивает передачу информации от рецепторов к нервным центрам и от них к исполнительным органам — скелетной мускулатуре, гладким мышцам внутренних органов и сосудов, железам внутренней и внешней секреции и т. д.
Распространение Нервных импульсов отождествляется с проведением потенциалов действия (см. Биоэлектрические потенциалы). Возникновение возбуждения может быть результатом раздражения (см.), напр, воздействие света на зрительный рецептор, звука на слуховой рецептор, или процессов, протекающих в тканях (спонтанное возникновение Н. и.). В этих случаях Н. и. обеспечивают согласованную работу органов при протекании какого-либо физиологического процесса (напр., в процессе дыхания Н. и. вызывают сокращение скелетных мышц и диафрагмы, результатом чего являются вдох и выдох, и т. д.).
В живых организмах передача информации может осуществляться и гуморальным путем, посредством выброса в русло крови гормонов, медиаторов и т. п. Однако преимущество информации, передаваемой при помощи Н. и., состоит в том, что она более целенаправленна, передается быстро и может быть точнее закодирована, чем сигналы, посылаемые гуморальной системой.
Факт, что нервные стволы являются путем, по к-рому передаются влияния от мозга к мышцам и в обратном направлении, был известен еще в эпоху античности. В средние века и вплоть до середины 17 в. считалось, что по нервам распространяется некая субстанция, подобная жидкости или пламени. Идея о электрической природе Н. и. возникла в 18 в. Первые исследования электрических явлений в живых тканях, связанных с возникновением и распространением возбуждения, были осуществлены Л. Гальвани. Г. Гельмгольц показал, что скорость распространения Н. и., к-рую ранее считали близкой к скорости света, имеет конечное значение и может быть точно измерена. Германн (L. Hermann) ввел в физиологию понятие потенциала действия. Объяснение механизма возникновения и проведения возбуждения стало возможным после создания С. Аррениусом теории электролитической диссоциации. В соответствии с этой теорией Бернштейн (J. Bernstein) предположил, что возникновение и проведение Н. и. обусловлено перемещением ионов между нервным волокном и окружающей средой. Англ. исследователи А. Ходжкин, Б. Катц и Э. Хаксли детально исследовали трансмембранные ионные токи, лежащие в основе развития потенциала действия. Позже стали интенсивно изучаться механизмы работы ионных каналов, по к-рым происходит обмен ионами между аксоном и окружающей средой, и механизмы, обеспечивающие способность нервных волокон проводить ряды Н. и. разного ритма и продолжительности.
Н. и. распространяется за счет местных токов, возникающих между возбужденным и невозбужденным участками нервного волокна. Ток, выходящий из волокна наружу в покоящемся участке, служит раздражителем. Наступающая после возбуждения в данном участке нервного волокна рефрактерность обусловливает поступательное движение Н. и.
Количественно соотношения разных фаз развития потенциала действия можно охарактеризовать, сопоставляя их по амплитуде и длительности во времени. Так, напр., для миелиновых нервных волокон группы А млекопитающих диаметр волокна находится в пределах 1—22 мк, скорость проведения — 5—120 м/сек, длительность и амплитуда высоковольтной части (пика, или спайка) — 0,4—0,5 мсек и 100—120 мв соответственно, следовой негативный потенциал — 12—20 мсек (3—5% от амплитуды спайка), следовой позитивный потенциал — 40—60 мсек (0,2% от амплитуды спайка).
Возможности передачи разнообразной информации расширяются за счет повышения скорости развития потенциала действия, скорости распространения, а также за счет повышения лабильности (см.) — т. е. способности возбудимого образования воспроизводить в единицу времени высокие ритмы возбуждения.
Возникновение Н. и. в нервных клетках (см.) или рецепторах (см.) связано с деполяризацией мембраны, т. е. со снижением величины электрического потенциала на мембране (потенциала покоя, или мембранного потенциала). Если величина мембранного потенциала снижается на 10—20% (пороговый критический уровень), то местный процесс переходит в распространяющийся — возникает потенциал действия (см. Возбуждение).
Конкретные особенности распространения Н. и. связаны со строением нервных волокон (см.). Сердцевина волокна (аксоплазма) обладает низким сопротивлением и, соответственно, хорошей проводимостью, а окружающая аксоплазму плазматическая мембрана — большим сопротивлением. Особенно велико электрическое сопротивление наружного слоя у миелинизированных волокон, у к-рых свободны от толстой миелиновой оболочки только перехваты Ранвье. В безмиелиновых волокнах Н. и. движется непрерывно, а в миелиновых — скачкообразно (сальтаторное проведение).
Различают декрементное и бездекрементное распространение волны возбуждения. Декрементное проведение, т. е. проведение возбуждения с угасанием, наблюдается в безмиелиновых волокнах. В таких волокнах скорость проведения Н. и. невелика и по мере отдаления от места раздражения раздражающее действие местных токов постепенно уменьшается вплоть до полного угасания. Декрементное проведение свойственно волокнам, иннервирующим внутренние органы, обладающие низкой функц, подвижностью. Без декрементное проведение характерно для миелиновых и тех безмиелиновых волокон, к-рые передают сигналы к органам, обладающим высокой реактивностью (напр., сердечной мышце). При бездекрементном проведении Н. и. проходит весь путь от места раздражения до места реализации информации без затухания.
Передача Н. и. с нервного волокна на мышечное или какой-либо другой эффектор осуществляется через синапсы (см.). У позвоночных животных в подавляющем большинстве случаев передача возбуждения на эффектор происходит при помощи выделения ацетилхолина (нервно-мышечные синапсы скелетной мускулатуры, синаптические соединения в сердце и др.). Для таких синапсов характерно строго одностороннее проведение импульса и наличие временной задержки передачи возбуждения.
В синапсах, в синаптической щели которых сопротивление электрическому току благодаря большой площади контактирующих поверхностей мало, происходит электрическая передача возбуждения. В них нет синаптической задержки проведения и возможно двустороннее проведение. Такие синапсы свойственны беспозвоночным животным.
Регистрация Н. и. нашла широкое применение в биол, исследованиях и клин, практике. Для регистрации используют шлейфные и чаще катодные осциллографы (см. Осциллография). При помощи микроэлектродной техники (см. Микроэлектродный метод исследования) регистрируют Н. и. в одиночных возбудимых образованиях — нейронах и аксонах. Возможности исследования механизма возникновения и распространения Н. и. значительно расширились после разработки метода фиксации потенциала. Этим методом были получены основные данные о ионных токах (см. Биоэлектрические потенциалы).
Нарушение проведения Н. и. происходит при повреждении нервных стволов, напр, при механических травмах, сдавливании в результате разрастания опухоли или при воспалительных процессах. Такие нарушения проведения Н. и. зачастую бывают необратимы. Следствием прекращения иннервации могут быть тяжелые функциональные и трофические расстройства (напр., атрофия скелетных мышц конечностей после прекращения поступления Н. и. вследствие необратимой травмы нервного ствола). Обратимое прекращение проведения Н. и. может быть вызвано специально, в терапевтических целях. Напр., с помощью анестезирующих средств блокируют импульсацию, идущую от болевых рецепторов в ц. н. с. Обратимое прекращение проведения Н. и. вызывает и новокаиновая блокада. Временное прекращение передачи Н. и. по нервным проводникам наблюдается и во время общего наркоза.
Библиография: Бpеже М. А. Электрическая активность нервной системы, пер. с англ., М., 1979; Жуков Е. К. Очерки по нервно-мышечной физиологии, Л., 1969; Коннели К. Восстановительные процессы и обмен веществ в нерве, в кн.: Совр, пробл. биофизики, пер. с англ., под ред. Г. М. Франка и А. Г. Пасынского, т. 2, с. 211, М., 1961; Костюк П. Г. Физиология центральной нервной системы, Киев, 1977; Латманизова Л. В. Очерк физиологии возбуждения, М., 1972; Общая физиология нервной системы, под ред. П. Г. Костюка, Л., 1979; Тасаки И. Нервное возбуждение, пер. с англ., М., 1971; Ходжкин А. Нервный импульс, пер. с англ., М., 1965; Ходоров Б. И. Общая физиология возбудимых мембран, М., 1975.
Читайте также: