
Законы проведения пд в нервах классификация теплопродукция и утомление
· Закон физиологической и анатомической непрерывности — возбуждение может распространяться по нерву только при сохранении его морфологической и функциональной целостности. Травматическое повреждение нерва нарушают или полностью прекращают проведение возбуждения.
· Закон изолированного проведения — возбуждение, распространяющееся в одной группе волокон (например, A«), не передается на волокна другой группы (например, В) того же ствола. Вследствие этого информация, передаваемая по разным типам волокон, носит строго направленный специфический характер.
· Закон двустороннего проведения — возбуждение, возникающее в каком-либо участке нерва, распространяется в обе стороны от очага возникновения.
Нерв имеет самую высокую функциональную лабильность, самый короткий период абсолютной рефрактерности и практически неутомляем.
Проведение возбуждения по нервным волокнам.
Безмиелиновые волокнана всем протяжении имеют одинаковую электропроводность и сопротивление. Вследствие деполяризации участка мембраны возникающий в нем локальный (местный) ток распространяется только на рядом расположенный невозбужденный. Волна деполяризации идет последовательно, не имея возможности миновать ни один из невозбужденных участков волокна.
Миелиновые волокнаимеют изолирующий слой, резко уменьшающий емкость мембраны нервного волокна и практически полностью предотвращающий утечку тока из него. Перехваты узла лишенные миелина, в отличие от миелиновых участков, имеют очень низкое сопротивление и поэтому являются центрами электрической активности. Практически все натриевые каналы сосредоточены в области перехватов — до нескольких тысяч на 1 мкм 2 , тогда как в миелиновых участках их вообще нет.
Итак, миелиновые волокна имеют очевидные преимущества:
· быстро, точно и дифференцированно проводят различные виды чувствительности, обеспечивая максимально быстрые, адекватные реакции.
В процессе эволюции высших организмов скачок в развитии нервной системы был, по-видимому, связан с началом миелинизации нервных волокон. В онтогенезе, особенно у человека, отмечена корреляция между ми-елинизацией некоторых проводящих путей и усложнением рефлекторного и целостного приспособительного поведения.
№ 5 Проведение возбуждения между клетками возбудимых тканей. Синапсы: строение, классификация и механизм проведения возбуждения в них. Функциональные свойства синапсов. Понятие о нейромедиаторах и нейромодуляторах.
Синапс — специализированный контакт между нервными клетками или нервными клетками и другими возбудимыми образованиями, обеспечивающий передачу возбуждения с сохранением его информационной значимости. С помощью синапсов осуществляется взаимодействие разнородных по функциям тканей организма, например нервной и мышечной, нервной и секреторной.
Структура синапса.
Пресинаптическое окончание аксона нейронапри подходе к иннервируемой клетке теряет миелиновую оболочку, что несколько снижает скорость распространения волны возбуждения. Небольшое утолщение на конце волокна, называемое синоптической бляшкой, содержит синаптические пузырьки размером 20—60 нм с медиатором — веществом, способствующим передаче возбуждения в синапсе.
Синаптическая щель— пространство между пресинаптическим окончанием и участком мембраны эффекторной клетки является непосредственным продолжением межклеточного пространства.
Постсинаптическая мембрана— участок эффекторной клетки, контактирующий с пресинаптической мембраной через синаптическую щель.
Классификация синапсов.
В соответствии с морфологическим принципом синапсы подразделяют на:
• аксо-аксональные (между двумя аксонами);
• аксодендритические (между аксоном одного нейрона и дендритом другого);
• аксосоматические (между аксоном одного нейрона и телом другого);
• дендродендритические (между дендритами двух или нескольких нейронов);
• нервно-мышечные (между аксоном мотонейрона и исчерченным мышечным волокном);
• аксоэпителиальные (между секреторным нервным волокном и грану-лоцитом);
• межнейронные (общее название синапсов между какими-либо элементами двух нейронов).
Все синапсы делят на центральные (в головном и спинном мозге) и периферические (нервно-мышечные, аксоэпителиальные и синапсы вегетативных ганглиев).
В соответствии с нейрохимическим принципомсинапсы классифицируют по виду химического вещества — медиатора, с помощью которого происходит возбуждение и торможение эффекторной клетки.
По конечному физиологическому эффекту , а также по изменению потенциала постсинаптической мембраны, различают возбуждающие и тормозные синапсы.
Механизм проведения возбуждения в синапсах. Передача возбуждения в химическом синапсе — сложный физиологический процесс, протекающий в несколько стадий. Он включает синтез и секрецию медиатора; взаимодействие медиатора с рецепторами постсинаптической мембраны; инактивирование медиатора. В целом синапс осуществляет последовательную трансформацию электрического сигнала, поступающего по нервному волокну, в энергию химических превращений на уровне синаптической щели и постсинаптической мембраны, которая затем снова трансформируется в энергию распространяющегося возбуждения в эффекторной клетке.
Свойства синапсов.
· Пластичность синапса.
· Одностороннее проведение возбуждения.
· Низкая лабильность и высокая утомляемость синапсаобусловлены временем распространения предыдущего импульса и наличием у него периода абсолютной рефрактерности.
· Высокая избирательная чувствительность синапсак химическим веществам обусловлена специфичностью хеморецепторов постсинаптической мембраны.
· Способность синапса трансформировать возбуждениесвязана с его низкой функциональной лабильностью и спецификой протекающих в нем химических процессов.
· Синаптическая задержка, т.е. время между приходом импульса в преси-наптическое окончание и началом ответа, составляет 1—3 мс. Суммация возбужденийопределяется переходом местного возбуждения в распространяющееся в результате временного взаимодействия серии возбуждающих постсинаптических потенциалов.
· Трофическая функция синапсов
Нейромедиаторы - физиологически активные вещества, вырабатываемые нервными клетками. С помощью нейромедиаторов нервные импульсы передаются от одного нервного волокна другому волокну или другим клеткам через синаптическую щель.
Нейромодуляторы - химические вещества, которые действуют как нейромедиаторы, но не ограничиваются синаптической щелью, а рассредотачиваются повсюду, модулируя действие многих нейронов в определенной области.
№ 7 Кровь как внутренняя среда организма: состав крови и количество крови в организме и ее функции. Основные гомеостатические показатели крови и их характеристика.
Кровь — важнейшая внутренняя жидкая среда организма, относительное постоянство состава которой обеспечивает оптимальные условия протекания клеточного метаболизма.
Вследствие наличия гистогематических барьеров истинной средой, взаимодействующей с клетками, является межклеточная жидкость. Вместе с другими жидкими средами организма она участвует в непрерывном обмене продуктов клеточного метаболизма, поступающих в кровь и лимфу, с извлекаемыми из крови веществами, необходимыми для энергетического и пластического обмена.
В конечном счете состав всех жидких сред организма определяется состоянием системы крови.
Система крови — совокупность образований, участвующих в поддержании гомеостаза тканей и органов:
• собственно кровь как жидкая разновидность соединительной ткани;
• органы кроветворения и кроворазрушения: костный мозг, вилочко-вая железа, лимфатические узлы, селезенка, печень;
• нейрогуморальный аппарат регуляции.
Функции крови.
Кроветворениепроисходит непрерывно в красном костном мозге. В среднем у человека в течение жизни образуется около 450 кг эритроцитов, 5400 кг гранулоцитов, 275 кг лимфоцитов и 40 кг тромбоцитов.
Кроворазрушениетакже протекает непрерывно в самом сосудистом русле, в селезенке и печени в количествах, эквивалентных вновь образующимся форменным элементам.
Синтез белков плазмыпроисходит непрерывно в печени.
Трофическая функция крови по отношению к клеткам заключается в переносе к ним от кишечника питательных веществ — аминокислот, липидов,
моно- и дисахаридов, витаминов, микроэлементов и др.
Экскреторная функция крови способствует выведению через почки, легкие, потовые железы и пищеварительный тракт токсичных продуктов метаболизма (мочевина, аммиак, билирубин, уробилин, двуокись углерода и др.), а также избытка воды и солей.
Защитная функция — одна из важнейших функций крови — реализуется в двух формах — иммунных реакциях (гуморальный и клеточный иммунитет) и свертывании (тромбоцитарный и коагуляционный гемостаз). Частным случаем защитной функции являются противосвертывающие механизмы системы крови.
Терморегуляторная функция способствует поддержанию температуры тела, особенно в условиях повышенной или пониженной температуры окружающей среды. Вследствие большой теплоемкости кровь переносит тепло от более нагретых к менее нагретым участкам тела и органам, регулируя таким образом физическую теплоотдачу.
Классификация нервных волокон по Эрлангеру-Гассеру
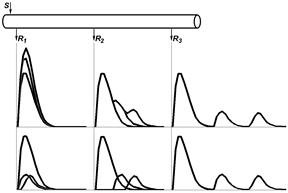
В 1939 г. американские[Мф54] физиологи Джозеф Эрлангер [Б55] и Герберт С.Гассер [Б56] [Б57] зарегистрировали токи действия от целого нервного ствола седалищного нерва лягушки на разных расстояниях от стимулирующего электрода (рис. 210041905).[Б58]
Было установлено, что регистрируемый суммарный потенциал имеет ряд пиков, которые были обозначены буквами латинского алфавита A, B, C (рис. , I). Пик A имел дополнительные пики, помеченные греческими буквами α, β, γ, δ (рис. , II). [Б59] В 1944 г. работа Дж.Эрлангера и Г.С.Гассера была оценена присуждением Нобелевской премии[Б60] .
Нервы у позвоночных состоят из трех основных групп волокон (А, В и С), различающихся по степени миелизации, диаметру волокна, длительности пика ПД (скорости развития ПД), электровозбудимости, его компенсации и скорости проведения (все эти показатели в ряду А — В — С падают).
Группа А включает наиболее толстые хорошо миелинизированные моторные и чувствительные волокна; группа В — слабомиелинизированные, преганглионарные волокна автономной нервной системы; группа С - немиелинизированные, постганглионарные (симпатические) волокна.
В группе А в ряду a, b, g, d названные показатели тоже падают. Соотношения свойств этих групп волокон демонстрируются в табл. .
Необходимо заметить, что указанные соотношения порогов электрического раздражения групп волокон не отражают точного соотношения электровозбудимости их мембран. Относительно высокие пороги тонких волокон при их раздражении в нервном стволе определяются в основном тем обстоятельством, что тонкие волокна по сравнению с толстыми обладают более высоким входным сопротивлением. В них входит такая малая часть раздражающего тока, что при пороговой силе для Аa-волокон она совершенно недостаточна для создания на мембране более тонких волокон сколько-нибудь существенной деполяризации. По этой же причине (высокое RI) отводимые от ствола (внеклеточно) ПД тонких волокон предстают значительно меньшими, чем ПД толстых волокон.

Рис. . Составные части потенциала действия смешанного нерва.
I – при относительно медленной скорости записи.
II – при относительно высокой скорости записи.
Объяснение в тексте. По оси абсцисс – время, по оси ординат амплитуда составного потенциала в мВ.
Суммарная электрическая активность нерва создается его волокнами, каждое из которых генерирует свой стандартный по амплитуде и временным параметрам ПД, распространяющийся в обе стороны от точки, к которой приложено раздражение. Суммарный электрический сигнал нерва зависит от числа активных волокон, синхронности их активности, способа отведения и других обстоятельств.
При дальнейшем увеличении силы стимула этот ПД несколько удлиняется во времени. Все изменения амплитуды и длительности пика ПД нерва при усилении стимула определяются ростом числа активных волокон, подключением к низкопороговым и быстрым А(альфа)-волокнам более высокопороговых медленных бета-, гамма-, дельта-волокон группы А, затем В- и, наконец, С-группы.
| Группы волокон (по Эрлангеру и Гассеру) | Диаметр, мкм | Скорость проведения, м/с |
| Aa | 13 - 22 | 70 - 120 |
| Ab | 8 -13 | 40 - 70 |
| Ag | 4 - 8 | 15 – 40 |
| Ad | 1 – 4 | 5 – 15 |
| B | 1 - 3 | 3 – 14 |
| C | 0,5 – 1,0 | 0,5 - 2 |
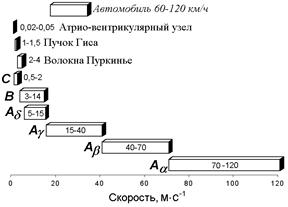
Таблица . Классификация нервных волокон по Дж.Эрлангеру и Х.Гассеру
| Группы волокон (по Эрлангеру и Гассеру) | Диаметр, мкм | Пороги электрического раздраженния (относительно Aa) | Длительность пика ПД 1 | Отрицательный следовой потенциал (ОСП) | Положительный следовой потенциал [Б61] | Скорость проведения, м/с | |
| Длительность, мс | Амплитуда СП, % к амплитуде ПД | Длительность, мс | Амплитуда СП, % к амплитуде ПД | ||||
| Aa | 13 - 22 | 1,0 | 0,4 | 15 – 20 | 40 - 60 | 0,2 | 70 - 120 |
| Ab | 8 -13 | 40 - 70 | |||||
| Ag | 4 - 8 | 15 – 40 | |||||
| Ad | 1 – 4 | 5 – 15 | |||||
| B | 1 - 3 | 11,7 | 1,2 | ОСП нет | 100 – 300 | 3 – 14 | |
| C | 0,5 – 1,0 | 100,0 | 2,0 | 50 – 60 | 300 - 1000 | 0,5 - 2 |
1 Приблизительно ту же величину имеют и абсолютные рефрактерные фазы
Основные свойства автоволн, касающиеся их распространения, распространяются и на потенциалы действия нервных волокон:
1. распространяется без затухания как по длине волокна, так и при его разветвлении (рис. 709170042).
2. не интерферируют (рис. 709170043).
3. не отражаются от препятствий (рис. 709170044).
4. направление распространения определяется зонами рефрактерности и покоя, обеспечивается двустороннее проведение возбуждения (рис. 709170045, 709170046).
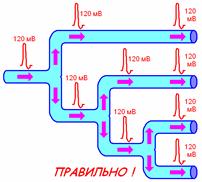
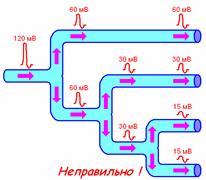
Рис. 709170042. Распространение ПД при разветвлении нервных волокон. Показан правильный вариант и ошибочный вариант, который часто встречается при ответах студентов.
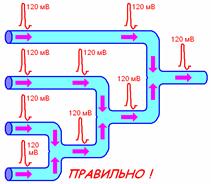
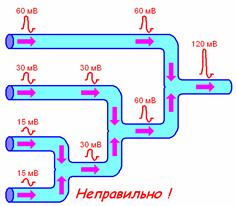
Рис. 709170043. Распространение ПД при схождении нервных волокон. Показан правильный вариант и ошибочный вариант, который часто встречается при ответах студентов.
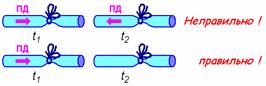
Рис. 709170044. ПД не отражаются от препятствий Показан правильный вариант и ошибочный вариант, который часто встречается при ответах студентов.
ПД проходит равные расстояния (L) от места действия стимула за одно и то же время (t) и сохраняет амплитуду при прочих равных условиях.
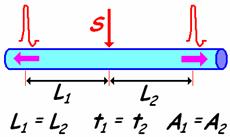
Рис. 709170045. Двустороннее проведение по нервным волокнам. L – расстояние от места действия стимула, t – время проведения ПД от места действия стимула до места расположения регистрирующих электродов, A – амплитуда ПД.
Обычно подчёркивается условие сохранения анатомической и физиологической непрерывности волокна
Анатомическая и физиологическая непрерывность волокна
Проведение импульсов возможно лишь при условии анатомической целостности волокна, поэтому как перерезка нервных волокон, так и любая травма поверхностной мембраны нарушают проводимость. Непроводимость наблюдается также при нарушении физиологической целостности волокна блокада натриевых каналов возбудимой мембраны тетродотоксином или местными aнестетиками, резкое охлаждение и т.п.). Проведение нарушается и при стойкой деполяризации мембраны нервного волокна К + , накапливающимися при ишемии в межклеточных щелях. Механическая травма, сдавливание нерва при воспалительном отеке тканей могут сопровождаться частичным или полным нарушением функции проведения.
Нервн. волокна (НВ) - это отростки нейронов, с помощью кот. осуще-ся связь между нейронами, а также нейронов с исполнит-ми кл. В состав НВ входят осевой цилиндр (нервный отросток) и глиальная оболочка. По взаимоотнош. осевых цилиндров с глиальными кл выделяют два типа нервных волокон: безмиелиновые и миелиновые. Оболочку безмиелиновых волокон образ шванновские клетки (леммоциты). При этом осевые цилиндры прогибают клеточную оболочку леммоцитов и погружаются в них. Клеточная мембрана обычно полностью окруж. каждый осевой цилиндр и смыкается над ним, образуя сдвоенную мембрану (мезаксон).
Оболочку миелиновых волокон образуют в периферич. нервной системе также шванновские клетки, а в ЦНС — олигодендроциты. В отличие от безмиелиновых волокон в миелиновых волокнах мезаксон удлиняется и спирально закручивается вокруг осевого цилиндра, образуя слой миелина (липидный футляр) вокруг осевого цилиндра. Миелиновая оболочка через равные уч-ки прерыв-ся, образ. свободн. от миелина небольшие уч-ки — узловые перехваты Ранвье. Участки волокон между перехватами наз-ся межузловыми сегментами, они образованы слоем миелина.Миелин сост. на 78 % из липидов (фосфолипид 42 %, цереброзидов — 28 %, холестерина — 25 %.) НО миелин требует затраты энергии и обеспеч. О2 и пит. в-вами(или - деструкция :( ). Нервные волокна имеют две основные ф-ции — изолирующ. ф-ция, способствующ. лучшему проведению биопотенциалов по отросткам нейронов. и транспорт в-в, обеспеч. трофическую ф-цию.
Проведение возбужд. по нервам подчиняется следующим законам: 1. Закон анатомической и физиологической целостности нерва. Первая нарушается при перерезе, вторая - действии веществ блокирующих проведение, например новокаина. 2. Закон двустороннего проведения возбуждения. Оно распространяется в обе стороны от места раздражения. В организме чаше всего возбуждение по афферентным путям оно идет к нейрону, а по эфферентным - от нейрона. Такое распространение называется ортодромным. Очень редко возникает обратное или антидромное распространение возбуждения. 3. Закон изолированного проведения. Возбуждение не передается с одного нервного волокна на другое, входящее в состав этого же нервного ствола. 4. Закон бездекрементного проведения. Возбуждение проводится по нервам без декремента, т.е. затухания. Следовательно, нервные импульсы не ослабляются, проходя по ним. 5. Скорость проведения прямопропорциональна диаметру нерва. (Нервные волокна обладают свойствами центрического кабеля, у которого не очень хорошая изоляция.
40. Механизм проведения потенциала действия в мякотных и безмякотных волокнах. Неутомляемость нервного волокна. Скорость проведения в различных нервах.
Функцию быстрой передачи возбуждения к нервной клетке и от нее выполняют ее отростки - дендриты и аксоны, т.е нервные волокна. В зависимости от структуры их делят на мякотные, имеющие миелиновую оболочку, и безмякотные. Эта оболочка формируется шванновскими клетками, являющиеся видоизмененными глиальными клетками. Они содержат миелин, который в основном состоит из липидов. Он выполняет изолирующую и трофическую функции. Участки, где оболочка прерывается, т.е. не покрыты миелином называют перехватами Ранвье. Функционально все нервные волокна делят на три группы:
1 Волокна типа А - это толстые волокна, имеющие миелиновую оболочку. В эту группу входят 4 подтипа: (альфа) двигательные волокна скелетных мышц и афферентные нервы (рецепторов растяжения). Скорость проведения по ним максимальна (бета) - афферентные волокна, идущие от рецепторов давления и прикосновения кожи. (гамма)- эфферентные волокна, идущие к мышечным веретенам (15-30 м/сек). (сигма) - афферентные волокна от температурных и болевых рецепторов кожи (12-30 м/сек).
2 Волокна группы В - тонкие миелинизированные волокна, являющиеся преганглионарными волокнами вегетативных эфферентных путей.
3 Волокна группы С, безмиелиновые постганглионарные волокна вегетативной нервной системы.
Проведение возбуждения по нервам подчиняется следующим законам: 1. Закон анатомической и физиологической целостности нерва. Первая нарушается при перерезе, вторая - действии веществ, блокирующих проведение, например новокаина. 2. Закон двустороннего проведения возбуждения. Оно распространяется в обе стороны от места раздражения. В организме чаше всего возбуждение по афферентным путям оно идет к нейрону, а по эфферентным - от нейрона. Такое распространение называется ортодромным. Очень редко возникает обратное или антидромное распространение возбуждения. 3. Закон изолированного проведения. Возбуждение не передается с одного нервного волокна на другое, входящее в состав этого же нервного ствола. 4. Закон бездекрементного проведения. Возбуждение проводится по нервам без декремента, т.е. затухания. Следовательно, нервные импульсы не ослабляются, проходя по ним.
5. Скорость проведения прямопропорциональна диаметру нерва. (Нервные волокна обладают свойствами центрического кабеля, у которого не очень хорошая изоляция. В основе механизма проведения возбуждения лежит возникновение местных токов: В результате генерации ПД в аксоном холмике и реверсии мембранного потенциала, мембрана аксона приобретает противоположный заряд. Снаружи она становится отрицательной, внутри положительной. Мембрана нижележащего, невозбужденного участка аксона заряжена противоположным образом. ''Поэтому между этими участками, по наружной и внутренней поверхностям мембраны начинают проходить местные, токи. Эти токи деполяризуют мембрану нижележащего невозбужденного участка нерва до критического уровня к в нем также генерируется ПД. Затем процесс повторяется и возбуждается более отдаленный участок нерва и т.д. Т.к. по мембране безмякотного волокна местные токи текут не прерываясь, поэтому такое проведение называется непрерывным. При непрерывном проведении местные токи захватывают большую поверхность волокна, поэтому им " требуется, длительное время для прохождения по участку волокна. В результате дальность, и. скорость проведения возбуждения по безмякотным волокнам небольшая. В мякотных волокнах участки, покрытые миелином обладают большим электрическим сопротивлением. Поэтому непрерывное проведение ПД Невозможно. При генерации ПД местные токи текут лишь между соседними, перехватами По закону “все или ничего" возбуждается ближайший к аксонному холмику перехват Ранвье, затем соседний нижележащий перехват и т.д. Такое проведение называется сальтаторным (прыжком). При этом механизме ослабления местных токов не происходит, и нервные импульсы распространяются на большое расстояние и с большой скоростью.
Скорость проведения потенциала действия в разных нервах. Скорость потенциала действия нервного ствола. Химические изменения в нерве при потенциале действия. Теплопродукция и утомление.
Энергозатраты нервного волокна на проведение ПД относительно невелики, поскольку возбуждаются только перехваты Ранвье, площадь которых составляет менее 1% общей поверхности мембраны аксона. Поэтому даже после длительных ритмических пачек ПД трансмембранный градиент концентраций ионов практически не изменяется.
В физиологических условиях ПД движутся в одном направлении от места раздражения (ортодромное проведение). ПД, проходящий по нервному волокну, возбуждает следующий, но не предыдущий участок мембраны. Это связано с рефрактерностью предыдущего участка после возбуждения. Проведение в противоположном направлении (антидромное проведение) возможно при травматическом поражении нервных волокон и в редких случаях (аксон–рефлекс).
В периферической нервной системе волокна объединены с помощью соединительнотканных оболочек в нервные стволы (нервы). В одном нерве могут быть тысячи нервных волокон: например, в срединном и мышеч-но-кожном нервах имеется 27—37 тыс. нервных волокон. Волокна в нервах могут быть миелиновыми и безмиелиновыми, афферентными и эфферентными. В естественных условиях каждое волокно нерва возбуждается от своего источника (например, эфферентное — от аксонного холмика, афферентное — от рецептора), и ПД в них проводятся асинхронно. Кроме того, чувствительные и двигательные волокна проводят импульсы в противоположных направлениях. Суммарная электрическая активность нерва создается электрической активностью составляющих его волокон и зависит от числа возбужденных волокон, степени шунтирования местных токов невозбужденными волокнами, синхронности проведения ПД в волокнах. В связи с этим анализ суммарной электрической активности нерва (нейрограммы) представляет трудную задачу.
3. Большая скорость проведения возбуждения. Скорость проведения ПД в различных типах волокон нерва равна 0,5—120 м/с (см. табл. 5.2). Она значительно выше в миелиновых волокнах в связи с сальтаторным типом проведения ПД (см. раздел 5.2.3), а среди ми-елиновых волокон прямо пропорциональна диаметру волокна. Скорость проведения возбуждения в миелиновых нервных волокнах значительно выше, чем в других удлиненных возбудимых структурах, — в гладких миоци-тах (0,02—0,10 м/с), рабочих кардиомиоцитах (около 1 м/с), и только в миоцитах проводящей системы сердца и скелетных миоцитах скорость проведения ПД (2—5 м/с) достигает величин распространения ПД в низкоскоростных нервных волокнах (тип С и В). Передача возбуждения по нервным волокнам является наиболее скоростным из известных способов передачи информации на значительные расстояния в организме. Для сравнения отметим, что скорость передачи гуморальных влияний ограничена скоростью кровотока, которая равна от 0,5 мм/с в капиллярах до 0,25 м/с в аорте (средняя скорость).
Дата добавления: 2015-12-15 | Просмотры: 2165 | Нарушение авторских прав
Законы проведения возбуждения в нервах
При изучении проведения возбуждения в нервных волокнах уста лено несколько правил, или законов , протекания этого процесса.
Закон физиологической непрерывности нерва . Обязательным условием проведения возбуждения по нервному волокну является анатомия и функциональная целость возбудимой мембраны осевого цилиндра.
Поэтому не только перерезка нерва, но и любое воздействие, нарушающее целость мембраны осевого цилиндра, например перевязка нерва, чрезмерное натяжение нервных волокон, создают непроводимость. Непроводимость наступает также при воздействиях, нарушающих генерацию нервного импульса, чрезмерное охлаждение или согревание, прекращение кровоснабжения, различные химические агенты, в частности местные обезболивающие — новокаин, кокаин, дикаин, прекращают проведение по нерву.
На основании подобных фактов сформулирован закон, гласящий, что проведение возможно только лишь при условии физиологической непрерывностн нервных волокон.
Закон двустороннего проведения . При нанесении раздражения на нервное волокно возбуждение распространяется по нему двусторонне, т.е. и в центробежном и в центростремительном направлениях. Это доказывают следующим опытом.
К нервному волокну, безразлично к двигательному или чувствительному, прикладывают две пары электродов, связанных с двумя электроизмерительными приборами А и Б ( рис. 154 ). Раздражение наносят между этими электродами. В результате двустороннего проведения возбуждения приборы зарегистрируют прохождение импульса как под электродами А, так и под электродами Б.
Рис. 154. Доказательство двустороннего проведения импульса в нерве (объяснение в тексте).
Доказательство двустороннего проведения возбуждения может быть получено и без помощи специальной электрофизиологической аппаратуры. Одна из бедренных мышц лягушки — m. gracilis — иннервирована таким образом, что к двум ее половинам подходят разветвления аксонов одних и тех же двигательных нервных волокон. На этой мышце В. Кюне проделал следующий опыт: он разрезал мышцу на две части, соединенные только посредством разветвлений нервных волокон, а затем одну часть мышцы он раздражал электрическим током. Это приводило к сокращению обеих половин мышцы, так как возбуждение распространялось по веточкам одного и того же аксона сначала центростремительно, а затем центробежно.
Закон двустороннего проведения нерва не противоречит тому факту, что в рефлекторных дугах центральной нервной системы и в нервных окончаниях на периферии возбуждение распространяется односторонне. Эта односторонность есть свойство не нервного волокна, а синапса, т. е. места перехода возбуждения с одного нейрона на другой или с нейрона на иннервированную им ткань.
Закон изолированного проведения по нерву . Всякий периферический нервный ствол состоит из большого числа нервных волокон — двигательных, чувствительных и вегетативных. Волокна нервных стволов снабжают большое число иногда далеко отстоящих друг от друга периферических образований. Так, блуждающий нерв иннервирует все органы грудной и значительную часть органов брюшной полости, седалищный нерв иннервирует всю мускулатуру, костный аппарат, сосуды и кожу нижней конечности.
Понятно, что нормальное функционирование всех периферических органов и тканей возможно лишь в том случае, если импульсы распространяются по каждому волокну нерва изолированно, не переходя с одного волокна на другое и оказывая действие только на те клетки, на которых нервные волокна оканчиваются.
Изолированное проведение в отдельных волокнах смешанного нерва может быть доказано опытом на скелетной мышце, иннервированной смешанным нервом, в образовании которого участвует несколько спинномозговых корешков. Если раздражать один из этих корешков, то сокращается не вся мышца, как это было бы, если бы возбуждение переходило с одних нервных волокон на другие, а только те группы мышечных волокон, которые иннервированы раздражаемым корешком. Еще более строгое доказательство изолированного проведения возбуждения может быть получено ври отведении потенциалов действия от различных нервных волокон нервного ствола.
Читайте также: