
Складки кори мозку називаються
Кора́ больших полуша́рий головного мозга
Слой серого вещества толщиной 1—5 мм, покрывающий полушария большого мозга млекопитающих животных и человека. Эта часть головного мозга (См. Головной мозг), развившаяся на поздних этапах эволюции животного мира, играет исключительно важную роль в осуществлении психической, или высшей нервной деятельности (См. Высшая нервная деятельность), хотя эта деятельность является результатом работы мозга как единого целого. Благодаря двусторонним связям с нижележащими отделами нервной системы, кора может участвовать в регуляции и координации всех функций организма. У человека кора составляет в среднем 44% от объёма всего полушария в целом. Её поверхность достигает 1468—1670 см 2 .
Строение коры. Характерной особенностью строения коры является ориентированное, горизонтально-вертикальное распределение составляющих её нервных клеток по слоям и колонкам; таким образом, корковая структура отличается пространственно упорядоченным расположением функционирующих единиц и связей между ними (рис. 1). Пространство между телами и отростками нервных клеток коры заполнено нейроглией (См. Нейроглия) и сосудистой сетью (капиллярами). Нейроны коры подразделяются на 3 основных типа: пирамидные (80—90% всех клеток коры), звездчатые и веретенообразные. Основные функциональный элемент коры — афферентно-эфферентный (т. е. воспринимающий центростремительные и посылающий центробежные стимулы) длинноаксонный пирамидный нейрон (рис. 2). Звездчатые клетки отличаются слабым развитием Дендритов и мощным развитием Аксонов, которые не выходят за пределы поперечника коры и охватывают своими разветвлениями группы пирамидных клеток. Звездчатые клетки выполняют роль воспринимающих и синхронизирующих элементов, способных координировать (одновременно тормозить или возбуждать) пространственно близкие группы пирамидных нейронов. Корковый нейрон характеризуется сложным субмикроскопическим строением (см. Клетка). Различные по топографии участки коры отличаются плотностью расположения клеток, их величиной и другими характеристиками послойной и колончатой структуры. Все эти показатели определяют архитектуру коры, или её цитоархитектонику (см. рис. 1 и 3).
Наиболее крупные подразделения территории коры — древняя (палеокортекс), старая (архикортекс), новая (неокортекс) и межуточная кора. Поверхность новой коры у человека занимает 95,6%, старой 2,2%, древней 0,6%, межуточной 1,6%.
Если представить себе кору мозга в виде единого покрова (плаща), одевающего поверхность полушарий, то основная центральная часть его составит новая кора, в то время как древняя, старая и межуточная займут место на периферии, т. е. по краям этого плаща. Древняя кора у человека и высших млекопитающих состоит из одного клеточного слоя, нечетко отделённого от нижележащих подкорковых ядер; старая кора полностью отделена от последних и представлена 2—3 слоями; новая кора состоит, как правило, из 6—7 слоев клеток; межуточные формации — переходные структуры между полями старой и новой коры, а также древней и новой коры — из 4—5 слоев клеток. Неокортекс подразделяется на следующие области: прецентральную, постцентральную, височную, нижнетеменную, верхнетеменную, височно-теменно-затылочную, затылочную, островковую и лимбическую. В свою очередь, области подразделяются на подобласти и поля. Основной тип прямых и обратных связей новой коры — вертикальные пучки волокон, приносящие информацию из подкорковых структур к коре и посылающие её от коры в эти же подкорковые образования. Наряду с вертикальными связями имеются внутрикортикальные — горизонтальные — пучки ассоциативных волокон, проходящие на различных уровнях коры и в белом веществе под корой. Горизонтальные пучки наиболее характерны для I и III слоев коры, а в некоторых полях для V слоя. Горизонтальные пучки обеспечивают обмен информацией как между полями, расположенными на соседних извилинах, так и между отдалёнными участками коры (например, лобной и затылочной).
Функциональные особенности коры обусловливаются упомянутым выше распределением нервных клеток и их связей по слоям и колонкам. На корковые нейроны возможна конвергенция (схождение) импульсов от различных органов чувств. Согласно современным представлениям, подобная конвергенция разнородных возбуждений — нейрофизиологический механизм интегративной деятельности головного мозга, т. е. анализа и синтеза ответной деятельности организма. Существенное значение имеет и то, что нейроны сведены в комплексы, по-видимому, реализующие результаты конвергенции возбуждений на отдельные нейроны. Одна из основных морфо-функциональных единиц коры — комплекс, называемый колонкой клеток, который проходит через все корковые слои и состоит из клеток, расположенных на одном перпендикуляре к поверхности коры. Клетки в колонке тесно связаны между собой и получают общую афферентную веточку из подкорки. Каждая колонка клеток отвечает за восприятие преимущественно одного вида чувствительности. Например, если в корковом конце кожного анализатора (См. Кожный анализатор) одна из колонок реагирует на прикосновение к коже, то другая — на движение конечности в суставе. В зрительном анализаторе (См. Зрительный анализатор) функции восприятия зрительных образов также распределены по колонкам. Например, одна из колонок воспринимает движение предмета в горизонтальной плоскости, соседняя — в вертикальной и т. п.
Локализация функций в коре характеризуется динамичностью в силу того, что, с одной стороны, имеются строго локализованные и пространственно отграниченные зоны коры, связанные с восприятием информации от определенного органа чувств, а с другой — кора является единым аппаратом, в котором отдельные структуры тесно связаны и в случае необходимости могут взаимозаменяться (т. н. пластичность корковых функций). Кроме того, в каждый данный момент корковые структуры (нейроны, поля, области) могут образовывать согласованно действующие комплексы, состав которых изменяется в зависимости от специфических и неспецифических стимулов, определяющих распределение торможения (См. Торможение) и возбуждения (См. Возбуждение) в коре. Наконец, существует тесная взаимозависимость между функциональным состоянием корковых зон и деятельностью подкорковых структур. Территории коры резко различаются по своим функциям. Большая часть древней коры входит в систему обонятельного анализатора. Старая и межуточная кора, будучи тесно связанными с древней корой как системами связей, так и эволюционно, не имеют прямого отношения к обонянию. Они входят в состав системы, ведающей регуляцией вегетативных реакций и эмоциональных состояний организма (см. Ретикулярная формация, Лимбическая система). Новая кора — совокупность конечных звеньев различных воспринимающих (сенсорных) систем (корковых концов Анализаторов).
Кроме того, локализация функций в первичных зонах многократно дублируется по механизму, напоминающему голографию (См. Голография), когда каждый самый маленький участок запоминающего устройства содержит сведения о всём объекте. Поэтому достаточно сохранности небольшого участка первичного сенсорного поля, чтобы способность к восприятию почти полностью сохранилась. Вторичные поля получают проекции от органов чувств через дополнительные переключения в подкорке, что позволяет производить более сложный анализ того или иного образа. Наконец, третичные поля, или ассоциативные зоны, получают информацию от неспецифических подкорковых ядер, в которых суммируется информация от нескольких органов чувств, что позволяет анализировать и интегрировать тот или иной объект в ещё более абстрагированной и обобщённой форме. Эти области называются также зонами перекрытия анализаторов. Первичные и отчасти вторичные поля — возможный субстрат первой сигнальной системы (См. Первая сигнальная система), а третичные зоны (ассоциативные) — второй сигнальной системы (См. Вторая сигнальная система), специфичной для человека (И. П. Павлов). Эти межанализаторные структуры определяют сложные формы мозговой деятельности, включающие и профессиональные навыки (нижнетеменная область), и мышление, планирование и целенаправленность действий (лобная область), и письменную и устную речь (нижняя лобная подобласть, височная, височно-теменно-затылочная и нижнетеменная области). Основные представители первичных зон в затылочной области — поле 17, где спроецирована сетчатка, в височной — поле 41, где спроецирован Кортиев орган, в прецентральной области — поле 4, где осуществляется проекция проприорецепторов в соответствии с расположением мускулатуры, в постцентральной — поля 3 и 1, где спроецированы экстерорецепторы в соответствии с их распределением в коже. Вторичные зоны представлены полями 8 и 6 (Двигательный анализатор), 5 и 7 (кожный анализатор), 18 и 19 (зрительный анализатор), 22 (Слуховой анализатор). Третичные зоны представлены обширными участками лобной области (поля 9, 10, 45, 44 и 46), нижнетеменной (поля 40 и 39), височно-теменно-затылочной (поле 37).
Корковые структуры играют первостепенную роль в обучении животных и человека. Однако образование некоторых простых условных рефлексов (См. Условные рефлексы), главным образом с внутренних органов, может быть обеспечено подкорковыми механизмами. Эти рефлексы могут образовываться и на низших уровнях развития, когда ещё нет коры. Сложные условные рефлексы, лежащие в основе целостных актов поведения (См. Поведение), требуют сохранности корковых структур и участия не только первичных зон корковых концов анализаторов, но и ассоциативных — третичных зон. Корковые структуры имеют прямое отношение и к механизмам памяти (См. Память). Электрораздражение отдельных областей коры (например, височной) вызывает у людей сложные картины воспоминаний.
Характерная особенность деятельности коры — её спонтанная электрическая активность, регистрируемая в виде электроэнцефалограммы (ЭЭГ). В целом кора и её нейроны обладают ритмической активностью, которая отражает происходящие в них биохимические и биофизические процессы. Эта активность имеет разнообразную амплитуду и частоту (от 1 до 60 гц) и изменяется под влиянием различных факторов.
Ритмическая активность коры нерегулярна, однако можно по частоте потенциалов выделить несколько разных типов её (альфа-, бета-, дельта- и тета-ритмы). ЭЭГ претерпевает характерные изменения при многих физиологических и патологических состояниях (различных фазах сна (См. Сон), при опухолях, судорожных припадках и т. и.). Ритм, т. е. частота, и амплитуда биоэлектрических потенциалов (См. Биоэлектрические потенциалы) коры задаются подкорковыми структурами, которые синхронизируют работу групп корковых нейронов, что и создаёт условия для их согласованных разрядов. Этот ритм связан с апикальными (верхушечными) дендритами пирамидных клеток. На ритмическую деятельность коры накладываются влияния, идущие от органов чувств. Так, вспышка света, щелчок или прикосновение к коже вызывают в соответствующих зонах т. н. первичный ответ, состоящий из ряда позитивных волн (отклонение электронного луча на экране осциллографа вниз) и негативной волны (отклонение луча вверх). Эти волны отражают деятельность структур данного участка коры и меняются в её различных слоях.
Филогенез и онтогенез коры. Кора — продукт длительного эволюционного развития, в процессе которого сначала появляется древняя кора, возникающая в связи с развитием обонятельного анализатора у рыб. С выходом животных из воды на сушу начинает интенсивно развиваться т. н. плащевидная, полностью обособленная от подкорки часть коры, которая состоит из старой и новой коры. Становление этих структур в процессе приспособления к сложным и разнообразным условиям наземного существования связано (совершенствованием и взаимодействием различных воспринимающих и двигательных систем. У земноводных кора представлена древней и зачатком старой коры, у пресмыкающихся хорошо развиты древняя и старая кора и появляется зачаток новой коры. Наибольшего развития новая кора достигает у млекопитающих, а среди них у приматов (обезьяны и человек), хоботных (слоны) и китообразных (дельфины, киты). В связи с неравномерностью роста отдельных структур новой коры её поверхность становится складчатой, покрываясь бороздами и извилинами. Совершенствование коры конечного мозга у млекопитающих неразрывно связано с эволюцией всех отделов центральной нервной системы. Этот процесс сопровождается интенсивным ростом прямых и обратных связей, соединяющих корковые и подкорковые структуры. Т. о., на более высоких этапах эволюции функции подкорковых образований начинают контролироваться корковыми структурами. Данное явление получило название кортиколизации функций. В результате кортиколизации ствол мозга образует с корковыми структурами единый комплекс, а повреждение коры на высших этапах эволюции приводит к нарушению жизненно важных функций организма. Наибольшие изменения и увеличение в процессе эволюции новой коры претерпевают ассоциативные зоны, в то время как первичные, сенсорные поля уменьшаются по относительной величине. Разрастание новой коры приводит к вытеснению старой и древней на нижнюю и срединную поверхности мозга.
Корковая пластинка появляется в процессе внутриутробного развития человека сравнительно рано — на 2-м месяце. Раньше всего выделяются нижние слои коры (VI—VII), затем — более высоко расположенные (V, IV, III и II; см. рис. 1). К 6 месяцам у эмбриона уже имеются все цитоархитектонические поля коры, свойственные взрослому человеку. После рождения в росте коры можно выделить три переломных этапа: на 2—3-м месяце жизни, в 2,5—3 года и в 7 лет. К последнему сроку цитоархитектоника коры полностью сформирована, хотя тела нейронов продолжают увеличиваться до 18 лет. Корковые зоны анализаторов завершают своё развитие раньше, и степень их увеличения меньше, чем у вторичных и третичных зон. Отмечается большое разнообразие в сроках созревания корковых структур у разных индивидуумов, что совпадает с разнообразием сроков созревания функциональных особенностей коры. Т. о., индивидуальное (Онтогенез) и историческое (Филогенез) развитие коры характеризуется сходными закономерностями.
Лит.: Орбели Л. А., Вопросы высшей нервной деятельности, М.— Л., 1949; Цитоархитектоника коры большого мозга человека. Сб. ст., М., 1949; Филимонов И. Н., Сравнительная анатомия коры большого мозга млекопитающих, М., 1949; Павлов И. П., Двадцатилетний опыт объективного изучения высшей нервной деятельности животных, Полн. собр. соч., 2 изд., т. 3, кн. 1—2, М., 1951; Брейзье М., Электрическая активность нервной системы, пер. с англ., М., 1955; Сепп Е. К., История развития нервной системы позвоночных, 2 изд., М., 1959; Лурия А. Р., Высшие корковые функции человека и их нарушения при локальных поражениях мозга, М., 1962; Воронин Л. Г., Курс лекций по физиологии высшей нервной деятельности, М., 1965; Поляков Г. И., О принципах нейронной организации мозга, М., 1965; Корковая регуляция деятельности подкорковых образований головного мозга. Сб. ст., Тб., 1968; Анохин П. К., Биология и нейрофизиология условного рефлекса, М., 1968; Беритов И. С., Структура и функции коры большого мозга, М., 1969.
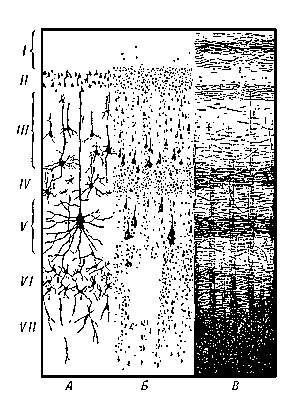
Рис. 1. Схема строения коры головного мозга человека: I — зональный слой, II — наружный зернистый слой, III — пирамидный слой, IV — внутренний зернистый слой, V — ганглионарный слой, VI — слой треугольных клеток, VII — слой веретеновидных клеток. А — нейронное строение, Б — цитоархитектоника, В — волоконная структура.
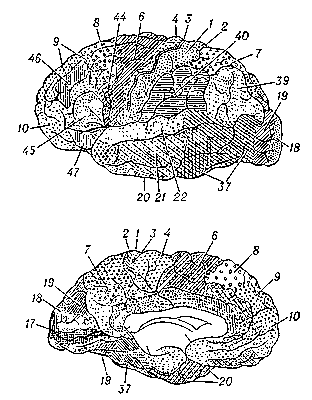
Рис. 3. Карта цитоархитектонических полей коры головного мозга человека: А — наружная поверхность полушария, Б — внутренняя поверхность полушария. Номерами и различной штриховкой обозначены цитоархитектонические поля коры.
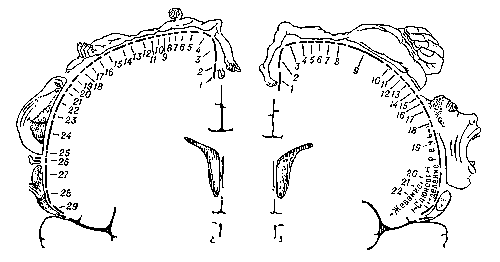
Рис. 4. Представительство чувствительных функций тела в задней центральной извилине (А) и двигательных функций — в передней центральной извилине (Б). А: 1 — половые органы; 2 — пальцы; 3 — ступня; 4 — голень; 5 — бедро; 6 — туловище; 7 — шея; 8 — голова; 9 — плечо; 10 — рука; 11 — локоть; 12 — предплечье; 13 — запястье; 14 — кисть; 15 — мизинец; 16 — безымянный палец; 17 — средний палец; 18 — указательный палец; 19 — большой палец; 20 — глаз; 21 — нос; 22 — лицо; 23 — верхняя губа; 24 — губы; 25 — нижняя губа; 26 — зубы, дёсны и челюсть; 27 — язык; 28 — глотка; 29 — внутренние органы. Б: 1 — пальцы; 2 — лодыжка; 3 — колено; 4 — бедро; 5 — туловище; 6 — плечо; 7 — локоть; 8 — запястье; 9 — кисть; 10 — мизинец; 11 — безымянный палец; 12 — средний палец; 13 — указательный палец; 14 — большой палец; 15 — шея; 16 — бровь; 17 — веко и глазное яблоко; 18 — лицо; 19 — губы; 20 — челюсть; 21 — язык; 22 — глотание. Размеры частей тела, изображенного на рисунке, соответствуют представительству двигательных и чувствительных функций организма в передней и задней центральных извилинах коры.
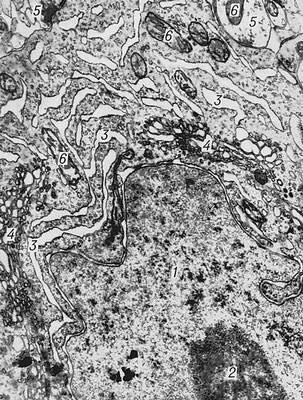
Рис. 2. Электронномикроскопическое строение пирамидной клетки коры головного мозга белой крысы: 1 — ядро; 2 — ядрышко; 3 — канальцы эндоплазматической сети; 4 — комплекс Гольджи; 5 — синапсы; 6 — митохондрии.
Более того, у человека наивысший уровень нервной системы стал доминирующим не только согласно нашему о нем мнению, но и по реальной массе. Центральная нервная система среднестатистического человека весит 1480 г. Из этого веса на долю спинного мозга (то есть на долю низшего и самого примитивного уровня) приходится около 30 г, то есть около 2%. Что же касается головного мозга, то в его массе превалирует масса конечного мозга, вес которого составляет 5/6 общего веса головного мозга.
Кора сложным образом свернута в складки, как я уже говорил в предыдущей главе. Линии, которые разделяют между собой складки, называются бороздами. Самые глубокие борозды называются щелями. Возвышения мозговой ткани между бороздами, которые выглядят как слегка уплощенные давлением крышки черепа цилиндры, называются извилинами. Борозды и извилины, создавая складчатость поверхности полушарий мозга, втрое увеличивают площадь серого вещества мозга. Количество серого вещества в глубине складок коры вдвое превышает его количество на уплощенной поверхности извилин.
Борозды и извилины - стандартные части мозга, и самые заметные из них, расположенные в одних и тех же местах у разных людей, имеют свои наименования.
Две самые постоянные борозды - это центральная и латеральная (то есть боковая) борозды, расположенные, естественно, в каждом полушарии большого мозга. (Полушария головного мозга по своему строению представляют собой зеркальные отражения друг друга.) Центральная борозда начинается на вершине мозга, непосредственно в его середине, и, немного изгибаясь, направляется вперед и вниз. Иногда эту борозду называют роландовой, по имени описавшего ее итальянского анатома XVIII века Луиджи Роландо. Латеральная борозда начинается у нижней поверхности полушария, отступя на одну треть расстояния от его переднего края, и направляется назад и немного вверх параллельно линии основания мозга. Закапчивается эта борозда, не дойдя полпути до заднего края большого мозга. Это самая заметная из всех борозд. Иногда ее называют сильвиевой бороздой по имени Сильвия (профессиональный псевдоним описавшего ее французского анатома XVII века).
Однако, несмотря на то что Галль и, в особенности, его ученики свернули с истинного пути, в их наблюдениях, тем не менее, присутствовало рациональное зерно. В 1861 году французский хирург Пьер Поль Брока при тщательном исследовании мозга умерших сумел показать, что у больных, страдавших неспособностью говорить и понимать речь (афазией), был поврежден определенный участок головного мозга. Эта область располагается в третьей левой лобной извилине, которая с тех пор называется областью Брока.
Этими исследованиями было установлено, что полоса коры, расположенная перед центральной бороздой, отвечает за стимуляцию двигательной активности скелетных мышц и называется поэтому двигательной областью коры. Тело представлено в двигательной области в перевернутом виде. Так, мышцы самой нижней части ног представлены в самой верхней части моторной коры, дальше книзу находятся области представительства бедра, потом туловища и рук, а в самом низу расположены участки, отвечающие за движения головы и шеи.
Кора головного мозга в двигательной зоне, так же как и в других участках, разделена на несколько слоев, которые анатомы четко отличают друг от друга. В одном из таких слоев, в обоих полушариях, расположены по 30 000 необычайно крупных клеток. Из-за своей формы они получили название пирамидных клеток, или клеток Беца, по имени русского анатома Владимира Беца, который впервые описал их в 1874 году. Волокна этих клеток управляют сокращениями мышц, причем каждая пирамидная клетка соединена волокнами со строго определенными частями какой-либо мышцы. Раздражение волокон более мелких клеток, слой которых расположен в коре выше клеток Беца, не приводит к сокращению мышц, но делает мышечные волокна чувствительными к стимуляции со стороны пирамидных клеток.
Волокна, исходящие из двигательной области коры, образуют пучок, который называется пирамидным путем, или пирамидным трактом. Этот тракт проходит через различные участки головного мозга, лежащие ниже коры, и выходит в спинной мозг. Поскольку пирамидный путь связывает кору (кортекс) со спинным мозгом, его называют еще кортикоспинальным трактом. Два пирамидных тракта, но одному из каждого полушария большого мозга, перекрещиваются в нижней части головного мозга и в верхних частях спинного мозга. В результате стимуляция двигательной области левого полушария приводит к сокращению мышц правой половины тела и наоборот.
Само существование пирамидной системы указывает нам способ объединения нервной системы в функциональную единицу. Головной мозг разделен на изолированные анатомические части - большой мозг, мозжечок и другие, которые будут описаны ниже, но это вовсе не значит, что каждая из них функционирует в отрыве от других частей. Напротив, пирамидная система контролирует деятельность двигательных участков других отделов центральной нервной системы от коры до спинного мозга. Есть нервные волокна, контролирующие двигательную активность нейронов, не являющихся пирамидными клетками и представляющих экстрапирамидную систему, которая также соединяет между собой все части центральной нервной системы. В анатомическом плане нервную систему лучше всего характеризовать по последовательным горизонтальным срезам, но в функциональном плане ее лучше всего исследовать по срезам вертикальным.
Спускаясь вниз от коры через нижележащие области пирамидного и экстрапирамидного трактов до собственно мышечных волокон, мы можем наблюдать умножение эффектов. Волокно единственной пирамидной клетки оказывает воздействие на несколько клеток спинного мозга. Каждая из этих последних управляет деятельностью многих нейронов периферической нервной системы (то есть той части нервной системы, которая расположена за пределами головного и спинного мозга), а каждый из этих нейронов управляет активностью нескольких мышечных волокон. В итоге получается, что одна пирамидная клетка может косвенно контролировать работу приблизительно до 150 000 мышечных волокон. Такое положение помогает осуществлять координацию мышечной активности.
Изменяя степень такой дивергенции, организм может при необходимости регулировать топкую настройку движений. Например, движения туловища могут регулироваться сравнительно небольшим количеством пирамидных клеток, так степень свободы движения торсом сильно ограничена. Совершенно иная ситуация складывается при движениях пальцами рук, которые призваны выполнять более разнообразные движения. Здесь дивергенция намного меньше, и каждая пирамидная клетка контролирует деятельность меньшего числа мышечных волокон.
Но кора головного мозга не просто контролирует ответные реакции. Для того чтобы реакция оказалась полезной и целенаправленной, кора головного мозга должна получать сигналы об ощущениях. В теменной доле, непосредственно позади центральной борозды, находится область коры, которая называется сенсорной.
Несмотря на такое обобщающее наименование, этот участок коры воспринимает отнюдь не все ощущения. Чувствительные волокна, берущие начало от кожи и внутренних органов тела, направляются в составе нервных пучков в головной мозг по проводящим путям спинного мозга. Некоторые из этих волокон остаются в составе спинного мозга, некоторые отходят от основного пути в нижележащие области мозга головного. В большинства своем эти волокна все же достигают коры. Эти достигшие коры волокна несут прежде всего информацию о прикосновениях и температуре, наряду с импульсами, возникающими в мышцах. Эти последние несут информацию, касающуюся положения тела в пространстве и взаимного расположения частей тела, обеспечивая сохранение равновесия. Существуют также генерализованные ощущения, которые не требуют для своего восприятия каких-либо специализированных сенсорных органов. (Эти и другие ощущения будут описаны в 10, 11 и 12-й главах.) В более узком смысле сенсорную область коры головного мозга часто называют соматосенсорной областью (то есть областью телесной чувствительности). Но даже и это слишком обобщающее наименование, потому что один из важнейших соматосенсорных стимулов - боль - не представлен в этой части коры. Боль воспринимается и обрабатывается в других областях мозга, расположенных ниже ее. Тот факт, что ощущения воспринимаются на различных горизонтальных уровнях нервной системы, показывает, что и здесь существует продольная унификация функций. Этим занимается ретикулярная активирующая система, которая координирует деятельность всех уровней центральной нервной системы, отвечающих за восприятие сенсорной информации.
Так же как в случае с двигательной областью, область сенсорной коры разделена на участки, которые в перевернутом виде воспринимают информацию об ощущениях с различных частей тела. На самом верху сенсорной области расположено представительство ног, следующие органы представлены в следующей последовательности сверху вниз - бедро, туловище, шея, плечо, предплечье, кисть, пальцы. Ниже области, воспринимающей ощущения с пальцев, находится область представительства головы. В самом низу находится представительство языка, которое, среди прочих, обрабатывает ощущение вкуса. (Другие ощущения химических веществ, например обоняние, локализовано в основании лобной доли. У человека эта доля очень мала, в отличие от других позвоночных, у которых она сильно развита.)
Участки сенсорной области, представляющие губы, язык и кисть руки (как и следует ожидать), развиты больше относительно размеров этих органов, чем участки, представляющие органы более крупные. Действительно, иногда на рисунки, изображающие кору головного мозга, наносят изображение перевернутого человечка, отдельные части тела которого наложены на области их представительства в коре. На рисунках как сенсорного, так и двигательного человечка его торс непропорционально мал, малы также ноги, хотя очень велики стопы, направленные к верхушке мозга. К нижней части коры направлены очень большая кисть руки, огромная голова и гигантские губы и язык.
Все это достаточно разумно. Что касается движений, то манипуляции ртом и языком в процессе порождения речи и движения кистью руки в процессе трудовой деятельности очень тонки, и именно они отличают людей от животных. Что же касается ощущений, то движения рукой не могут быть точными, если мы в каждый данный момент времени не будем ощущать ее точное положение в пространстве и взаимное расположение пальцев. Ощущения, информация о которых передается с губ и языка, не специфичны только для человека, поскольку прием пищи очень важен и для него, несмотря на весь его интеллект, поэтому сигналы с языка и губ требуют большого внимания головного мозга.
Два очень важных ощущения, - важных и специализированных, - зрение и слух, имеют в своем распоряжении специально зарезервированные для этого доли. Это участок височной доли, расположенный непосредственно книзу от сенсорной области, служащий для восприятия и обработки слуховых ощущений и называемый поэтому слуховой областью, и затылочная доля, в которой расположена зрительная область коры головного мозга. Зрительная кора расположена на самых задних участках коры обоих полушарий головного мозга.
Не нашли то, что искали? Воспользуйтесь поиском:
Человеческий организм всяким образом стремится к энергоемкости и пластичности. Небольшой орган, выполняющий определенную функцию лучше, чем орган большой, исполняющий ту же функцию. На дороге эволюции мозг (как многофункциональная система) прогрессировал этим путем: он формировался и увеличивался благодаря сложной системе извилин и борозд. Таким образом, находясь внутри ограниченной в объеме черепной коробке, конечный мозг увеличивал свою площадь, сохраняя при этом весь набор функций.
Что это такое
Извилины головного мозга это небольшие возвышения над его поверхностью, ограниченные бороздами. Эти складки располагаются на территории всего конечного мозга, и площадь их составляет в среднем 1200 см3. О том факте, что функциональная поверхность увеличивается благодаря специфическим складкам, говорят цифры: большая часть (2/3) коры располагается между складками в глубинах впадин. Такому явлению, как образование извилин, существует объяснение: в процессе внутриутробного развития мозг младенца развивается неравномерно в разных местах, и, вследствие этого, напряжение поверхностей в разных отделах отличается.
Борозды головного мозга это своеобразные канавки, разделяющие извилины друг от друга. Эти образования классифицируют: первичные, вторичные и третичные. Первый тип углублений образуются самыми первыми в процессе формирования плода. Вторичные борозды появляются позже и являются постоянными. Третичные канавки изменчивы: борозды могут менять свою форму, направление и даже размер. Данные углубления разделяют поверхность больших полушарий на основные доли: теменную, височную, лобную, островковую и затылочную.
Строение
Схема извилин и борозд головного мозга лучше всего видна на схематических изображениях. Углубления, разделяющие кору на две части (полушария) называются первичными. Кроме этого, существуют и другие фундаментальные ограничители отделов коры, а именно:
- Сильвиева борозда (латеральная, боковая): разделяет височную и лобную кору.
- Роландова впадина (центральная): отделяет теменную от лобной.
- Теменно-затылочная впадина: разграничивает затылочную и теменную долю мозга.
- Поясная впадина, переходящая в гиппокампальную: отделяет поверхность обонятельного мозга от других отделов.
Эти структуры также носят и другое название: борозды первого порядка головного мозга.
Всякий отдел конечного мозга вмещает в себя несколько извилин, разделяющихся вторичными впадинами. Третичные же углубления развиваются сугубо индивидуально: их наличие зависит от личностных особенностей человека и его умственных способностей. Третий тип выемок придает индивидуальный рельеф складкам.

Эта область конечного мозга ограничена тремя бороздами: латеральной, части затылочной и центральной. Боковая впадина берет свое начало от латеральной ямки. Развиваясь немного вверх и назад, образование заканчивается на верхнелатеральной поверхности.
На верхнем краю одного из полушария начинается центральная борозда. От его середины она идет кзади и частично вперед. Спереди от этой выемки располагается лобная доля мозга, а сзади – теменная кора.
Конец затылочной области служит краем теменной области. Эта канавка не имеет четкой границы, поэтому разделение осуществляется искусственно.
Данная часть полушарий обладает постоянными глубокими бороздами. Говоря об образованиях медиальной поверхности, в первую очередь, как правило, вспоминают о борозде мозолистого тела (1). Выше этой канавки располагается поясная впадина (2), образующая колено и в последующем ветвь. Также в этой области находится гиппокампальная борозда (3) или борозда морского коня. Ближе к затылочной доле располагается коллатеральная борозда (4). На территории задней части срединной поверхности лежит шпорная борозда (5).
Между первыми двумя образованиями располагается опоясывающая извилина. А гиппокампальная и коллатеральная канавка ограничивает извилину, принадлежащую височной коре полушария.

Эта часть мозга распространяется на разных отделах коры – височной, затылочной и лобной. Нижняя поверхность включает в себя следующие борозды:
- Обонятельная (1)
- Орбитальная (2)
- Прямая (3)
- Нижняя височная (4)
Эта область полушария не обладает выдающимися извилинами, однако, все же одну отметить следует – это язычковая извилина (5).
Читайте также: