
Меккелев хрящ нижней челюсти
На территории Китая палеонтологи нашли древнее млекопитающее, чьи слуховые кости в составе стремечка, молоточка и наковальни еще не утратили прямую связь с нижней челюстью. Эта находка наконец-то окончательно подтвердила справедливость предположения о том, что элементы среднего уха наземных позвоночных произошли из костей челюстного аппарата.
Недавно китайские палеонтологи сделали открытие, которое смогло подтвердить справедливость одной старой эволюционной реконструкции, которая, кстати, до сих пор присутствует в большинстве отечественных учебников по зоологии. Им удалось фактически доказать, что кости среднего уха современных наземных позвоночных произошли от челюстных элементов предковых форм. Найденное учеными Поднебесной примитивное млекопитающее, обитавшее примерно 120 миллионов лет тому назад явилось тем самым недостающим переходным звеном, которое так долго искали эволюционисты.
Собственно говоря, кому-то, может быть, подобное открытие покажется смешным — экая невидаль, итак любой школьник знает, откуда у млекопитающих взялось среднее ухо. На самом деле здесь не все так просто. Дело в том, что данная реконструкция, которую многие поколения отечественных учащихся принимало как нечто, уже давно доказанное, на самом деле таковой, строго говоря, не являлось.
Как известно, в науке ни одну теорию или даже гипотезу нельзя принимать на веру, то есть без соответствующих доказательств (именно этим наука отличается от других мировоззренческих систем, например, религии). Однако доказательства часто невозможно предъявить сразу — иногда на то, что бы добыть их, уходят годы или даже десятилетия. Кстати, если речь идет об эволюционных реконструкциях, то в этой области подобный временной разрыв между предположением и окончательным выводом — обычный случай.
До тех пор, пока нет четких доказательств, любая гипотеза принимается учеными в лучшем случае как презумпция (то есть с условием: "данное утверждение считается верным до тех пор, пока не доказано обратное"). В худшем же случае — просто как одна из рабочих версий. Многие известные нынче научные теории и законы существовали в виде презумпций достаточно длительное время (например, между появлением знаменитой теории относительности Эйнштейна и ее первым фактическим доказательством прошло больше десяти лет), а некоторые и поныне являются таковыми (всем известный закон сохранения массы и энергии).

Так вот, реконструкция, выводящая среднее ухо наземных позвоночных из челюстных элементов предковых форм появилась еще в концеXIX — начале XX веков. Ряд биологов, в числе которых были и знаменитые отечественные эволюционисты А. Н. Северцов и И. И. Шмальгаузен сделали подобное предположение, исходя, однако, не из палеонтологических, а из эмбриологических данных. Суть этой гипотезы заключалось в следующем.
Косточек среднего уха (у млекопитающих это стремечко, молоточек и наковальня, а у птиц и рептилий — только стремечко), как известно, нет ни у амфибий, ни у рыб, являющихся предками всех наземных позвоночных. Зато у последних имеется так называемый гиомандибулярный аппарат (комплекс из нескольких костей и хрящей), который осуществлял связь челюстей друг с другом, а также с черепом и жаберными дугами. Однако когда на базе древних рыб возникли первые земноводные, их череп несколько изменился, а жаберный аппарат вообще исчез.
В новом черепе этот гиомандибулярный аппарат утратил прежнюю роль подвеска челюстей. Кроме того, замена жаберного дыхания легочным (и кожным) сопровождалась прекращением деятельности вентиляционного механизма жаберной крышки, которая также редуцировалась за ненадобностью. В итоге, потеряв обе свои прежние функции, рассматриваемый нами элемент черепа также подвергся некоторой редукции.
Однако - поскольку гиомандибулярный аппарат располагался в черепе сбоку от слуховой капсулы и его отростки упирались в стенку последней и в накладные кости височной области черепа, под которыми здесь располагается полость - ему нашлось весьма интересное применение. Дело в том, что при нахождении организма предка всех позвоночных на суше эта полость оказалась заполненной воздухом и ее наружная стенка после редукции жаберной крышки стала относительно тонкой. Предполагается, что она могла вибрировать в ответ на колебания окружающего воздуха, как барабанная перепонка. А гиомандибулярный аппарат, упиравшийся в данную перепонку и в стенку слуховой капсулы, скорее всего, передавал трансформированные колебания воздуха к внутреннему уху. Так этот исходный элемент челюсти функционально превратился в составляющую примитивного среднего уха.
Далее его судьба у разных потомков древних амфибий была различной. У так называемых завроморфных рептилий, которые стали предками современных пресмыкающихся, динозавров и птиц от гиомандибулярного аппарата произошла только одна слуховая кость — стремечко, или слуховой столбик (это было доказано отечественными палеонтологами на примере черепа древней ящерицеподобной рептилии Bashkyroleter mesensis). В принципе этим животным, которые исходно представляли собой засадных хищников и очень медлительных бронированных фитофагов, для которых основным способом получения информации является зрение, дальнейшее усовершенствование слухового аппарата было просто не нужно — оно не несло никаких эволюционных выгод.

А вот более активные тероморфные рептилии, от которых впоследствии произошли млекопитающие, добавили к стремечку еще две кости нижней челюсти, функционально связанные с гиомандибулярным аппаратом (проще говоря, он за них цеплялся) — сочленовую и квадратную. В результате тероморфы остались с укороченной нижней челюстью (видимо, именно это потом подвигло их на дифференцировку зубов на резцы, клыки и коренные) и совершенным слуховым аппаратом среднего уха, состоящего из трех "усилителей" — стремечка, молоточка и наковальни. Им такой "радар" был жизненно необходим, поскольку в основном это были сумеречные хищники, привыкшие больше доверять своим ушам, нежели глазам.
По мнению ученых, эти процессы у древних наземных позвоночных происходили в интервале от 270 до 115 миллионов лет назад. Однако долгое время прямых подтверждений из палеонтологической летописи эта схема не имела — исследователи просто не находили останков промежуточных форм. Так что до начал нынешнего века данная гипотеза держалась лишь на данных по развитию зародышей наземных позвоночных — еще в первой половине прошлого века было установлено происхождение из одних и тех же структур зародыша квадратной и суставной костей в челюстях у рептилий и молоточка и наковальни в среднем ухе у млекопитающих.
Однако, как вы сами понимаете, подобное доказательство, строго говоря, не является исчерпывающим — оно позволяет лишь говорить о том, что так могло быть в процессе эволюции, но не о том, что так было на самом деле. Однако недавно китайским палеонтологам удалось добыть более авторитетные доказательства и перевести данную реконструкцию из разряда вероятных в разряд достоверных.
Еще в 2009 году ими был найден зверёк размером с бурундука, получивший название Maotherium asiaticus из группы древних млекопитающих морганукодонов (Morganucodon), которые вымерли, не оставив потомков. Их меккелев хрящ (производная гиомандибулярного аппарата) уже находился в состоянии окостенения, что может свидетельствовать о его грядущей утрате у более поздних видов и развитии на этом месте слуховых косточек. Произошло это 200-125 миллионов лет назад. Такой окостеневший гиомандибулярный аппарат уже был связан со слуховой капсулой и, скорее всего, играл роль "усилителя". Однако сочленовая и квадратная кость у данного животного еще находятся в составе нижней челюсти, хотя остаются с ним связанными.
А вот совсем недавно было найдено еще одно "недостающее звено". В Поднебесной было найдено древнее млекопитающее из рода Liaoconodon. Этот похожий на белку зверек обитал на нашей планете около 120 миллионов лет назад. Так вот, у него меккелев хрящ уже представляет собой настоящую кость (подобную слуховому столбику рептилий) и на найденном образце достаточно хорошо видно, что он служит связующим звеном между слуховой капсулой и квадратной и сочленовой костью, которые уже отделились от нижней челюсти, хотя находятся непосредственно рядом с ней. Как видите, это и есть тот самый промежуточный вариант, существование которого почти сто лет назад предсказал А. Н. Северцов.
"Это первое недвусмысленное доказательство, являющееся переходной формой", — говорит автор находки палеонтолог Цзинь Мэн. По словам ученого, с таким слуховым аппаратом этот зверек обладал уже очень хорошим слухом (по сравнению с рептилиями), чувствительным к высоким частотам, что возможно помогало ему в поиске насекомых в темноте ночного леса (он был активен по ночам, об этом говорят его весьма крупные глаза). Что, кстати, тоже подтверждает одно из вышеупомянутых предположений о причине дальнейшего совершенствования среднего уха у тероморф.
Итак, еще одна палеонтологическая реконструкция пополнилась рядом из промежуточных форм, который доказывает ее корректность. Так что теперь школьники и студенты, читающие об эволюции среднего уха древних позвоночных, могут быть уверены в том, что давным-давно, в юрском, а затем и в раннем меловом периоде все именно так и было…
Читайте самое интересное в рубрике "Наука и техника"
Встройте "Правду.Ру" в свой информационный поток, если хотите получать оперативные комментарии и новости:
Подпишитесь на наш канал в Яндекс.Дзен или в Яндекс.Чат
Добавьте "Правду.Ру" в свои источники в Яндекс.Новости или News.Google
Также будем рады вам в наших сообществах во ВКонтакте, Фейсбуке, Твиттере, Одноклассниках.
Мысли и впечатления // Pensoj kaj impresoj
Под катом - довольно специфический текст о моей работе. Содержание может показаться скучным, а в отдельных моментах - возможно даже немного шокирующим (ну, вы знаете, чем там анатомы-морфологи могут заниматься. ). Но есть несколько интересных картинок ручной работы (вы видите одну из них).
Последние полторы-две недели на работе у меня проходят в состоянии лёгкого опупения. Пришли первые стёкла с гистологии, которые нужно будет изучить и каким-то образом включить в мою научную работу, которая рано или поздно будет всё же дописана. Гистологию я никогда особо не любил да и изрядно подзабыл уже, так что зрелище, открывшееся мне в микроскопе было просто удивительным (полагаю, не менее удивительным, чем для тех моих френдов, которые к биологии и медицине не имеют совершенно никакого отношения). Плюс всё осложняется ещё и тем, что на этих стёклах - эмбриологический материал (то есть, попросту говоря, человеческий зародыш), где всё не так, как должно быть у взрослых людей, анатомию которых я более-менее знаю. В общем, разбираюсь и делаю открытия.
На всех картинках представлены гистологические срезы лицевого отдела головы человеческого зародыша возрастом около 10-12 недель (то есть, это уже не эмбрион, а молодой плодик). На самой первой картинке (вы её уже видели) - фронтальный (вертикальный) срез, в центре которого видна закладка будущей поднижнечелюстной слюнной железы (увеличение микроскопа - 40 раз). Тёмно-фиолетовое пятно в правой верхней части картинки - это нижняя челюсть (предположительно - задняя часть её основания или даже начальный отдел ветви).
На следующей картинке - ещё один вертикальный срез, но поле зрения располагается несколько выше (кадр не очень удачный, ну да ладно, переделаю ещё). Хорошо видна ветвь нижней челюсти (она уже оссифицируется, то есть, частично представлена костной тканью), но самое главное - это половинка тёмного пятна округлой формы в нижней части снимка левее челюсти. Увеличение такое же (40 раз).
По всем признакам - это хрящевая ткань. Но что именно это за структура - с ходу "никто догадаться не мог". У взрослых людей никаких подобных структур медиальнее (кнутри) от нижней челюсти нет, а это непонятное "хрящевое тело Можаева" ещё и было достаточно длинным, хорошо визуализировалось как на фронтальных, так и на горизонтальных срезах на довольно большом протяжении. Сегодня я "засел за инструкцию", то есть, стал освежать свои знания в области эмбриологии. И, кажется, сделал для себя удивительное открытие. Вот снимок этого тела крупным планом (увеличение 100 раз):
Как, вы ещё не догадались, что это? Это - удивительная структура, доставшаяся нам в наследство от наших далёких предков по филогенезу, а именно - так называемый Меккелев хрящ, являющийся производным первой жаберной дуги. Если максимально упростить, то у рыб скелетные элементы глотки образованы такими своеобразными полукольцами (5 пар у большинства современных рыб) - они поддерживают систему жабр и жаберные щели между ними. А у филогенетических потомков рыб - наземных позвоночных вроде нас с вами - с утратой жаберного дыхания эти дуги утрачиваются, а их производные образуют совершенно другие вещи, типа подъязычной кости, хрящей гортани и др.
Но у эмбрионов всё не так просто: в соответствии с биогенетическим законом Геккеля-Мюллера на ранних стадиях внутриутробного развития у зародышей наблюдаются признаки, схожие с анатомическими признаками филогенетических предков. У зародыша человека тоже имеются жаберные дуги, однако, ещё до рождения они продолжают развиваться. В частности, на определённом этапе внутриутробной перестройки первая жаберная дуга (обозначена на рисунке римской цифрой I) у человека образует длинный хрящ, косо направленный сзади наперёд - его-то и описал товарищ Меккель. У низших рыб он сохраняется всю жизнь и функционирует как нижняя челюсть. А вот у человека с Меккелевым хрящом происходят удивительные превращения.
Задние части этих хрящей у человека модифицируются в молоточек и наковальню - две маленькие слуховые косточки, расположенные в полости среднего уха. Передние части хрящей закостеневают и образуют самые передние части двух половинок нижней челюсти (у новорождённых нижняя челюсть состоит из двух половинок, соединённых спереди посередине тонким хрящом, который в течение 1-1,5 годов тоже полностью окостеневает). А вот средняя часть Меккелева хряща служит как бы "моделью", вокруг которой "нарастает" нижняя челюсть, при этом сам хрящ в своих средних отделах уменьшается и исчезает, образуя в итоге канал нижней челюсти.
Таким образом, на этих срезах (ещё раз напомню - после оплодотворения прошло около 10-12 недель!) удалось застать ту стадию, когда Меккелев хрящ ещё присутствует, но уже видно, как он вот-вот начнёт окостеневать (по его окружности видна "скорлупа", более плотная, чем сам хрящ). Этакий привет из нашего далёкого эволюционного прошлого. Я восхищён! :) Просто удивительно, как эту структуру можно было открыть аж в 1820-ом году!
А на этой картинке - просто слизистая полости рта (нёба). Обычный многослойный эпителий (увеличение - 400 раз). После Меккелева хряща даже как-то неприлично вести разговор о таких обыденных вещах.
Ну, и напоследок красивое фото ветви нижней челюсти (она окостеневает независимо от тела, которое формируется вокруг Меккелева хряща, и немножко раньше) под крупным увеличением (400 раз). За счёт относительно большой толщины среза и крупного увеличения создаётся эффект объёмности изображения. Просто удивительно, как всё хитро устроено. Микроскоп - замечательная игрушка! :)
Развитие нижней челюсти начинается с образования костной ткани из нескольких точек окостенения, расположенных в клетчатке, прилежащей к меккелеву хрящу. Постепенно формируется покровная кость, окружающая этот хрящ, сам же хрящ редуцируется, уступая место развивающемуся телу нижней челюсти.
Задние отделы челюсти, ее ветви, образуются независимо от меккелева хряща из соответствующих точек окостенения. Окостенение двух половин нижней челюсти заканчивается их сращением, т. е. превращением в непарную кость, уже после рождения, к концу 1-го года жизни.
Эмбрион человека (8-я неделя).
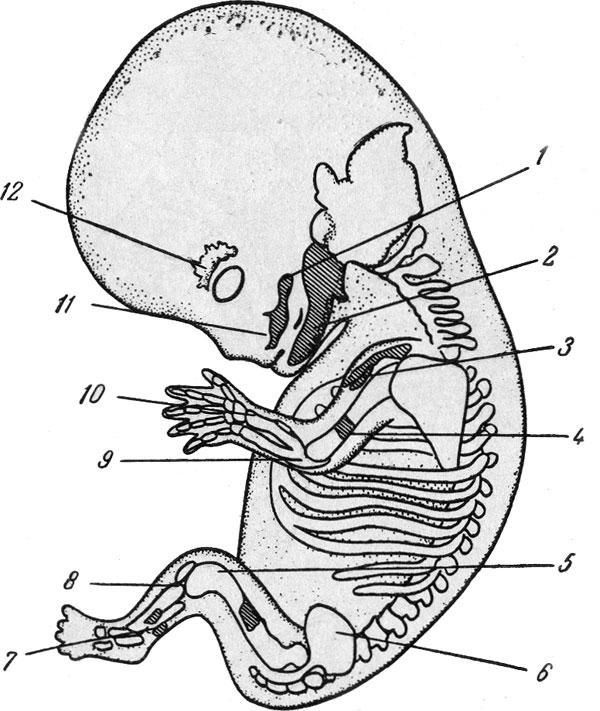
Эмбрион человека длиной 23,5 мм (8-я неделя), центры окостенения (по Дж. Шеферу): 1 — скуловая кость, 2 — нижняя челюсть, 3 — ключица, 4 — плечевая кость, 5 —бедренная кость, 6— подвздошная кость, 7 — малоберцовая кость, 8 — большеберцовая кость, 9 —локтевая кость, 10 — лучевая кость, 11 — верхняя челюсть, 12 — лобная кость.
Эмбрион человека (9-я неделя).
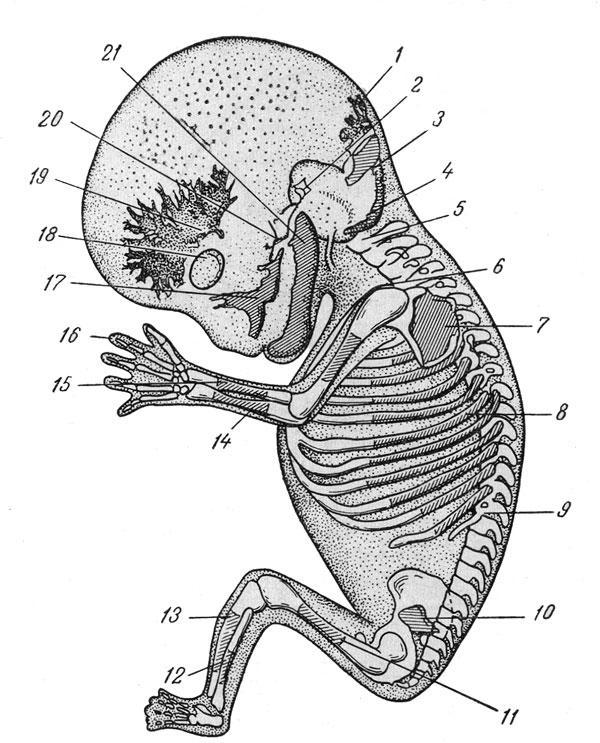
Эмбрион человека длиной 39 мм (9-я неделя), ход окостенения (по Дж. Шеферу): 1 — интерпарнетальный центр, 2 — чашуйчатая часть височной кости, 3 — супраокципитальный центр, 4 — затылочная дуга, 5 —нервная дужка, 6 — ключица, 7 —лопатка, 8 — VI ребро, 9 — XII ребро, 10— подвздошная кость, 11 — бедренная кость, 12 — малоберцовая кость, 13 — большеберцовая кость, 14 — локтевая кость, 15 — лучевая кость, 16— дистальная фаланга, 17 — верхняя челюсть, 18 — глаз, 19 — лобная кость, 20 — скуловая кость, 21 — нижняя челюсть.
Альвеолярный отросток челюсти развивается из мезенхимы, окружающей зачаток зуба. Закладка альвеолярного отростка нижней челюсти происходит на 3-м месяце утробного развития, отростка верхней челюсти — на 4-м месяце. Срастание альвеолярного отростка с телом нижней челюсти происходит к 5 месяцам, с телом верхней челюсти — к 6 месяцам.
С окончанием прорезывания зубов заканчивается формирование альвеолярного края, а с окончанием формирования корня — формирование его основания. В толще формирующихся челюстей образуются и развиваются зачатки зубов. С развитием и прорезыванием зубов тесно связаны рост и формирование челюстей.
Кожа детей наиболее раннего возраста по своим морфологическим и физиологическим особенностям отличается значительным своеобразием. Роговой слой тонок и состоит из 2—3 рядов слабо связанных между собой и постепенно слущивающихся клеток. Основной слой развит сильно, можно всегда увидеть интенсивное деление в нем эпителиальных клеток. Более толстый ростковый слой кожи и большая наклонность детского организма к пролиферативным…
Строение кости альвеолярного отростка в период прорезывания зубов отличается от его строения после прорезывания. В период прорезывания вершины межальвеолярных перегородок как бы срезаны в сторону прорезывающегося зуба, располагаются вблизи или на уровне его эмалево-цементной границы. При этом создается впечатление, что у коронки прорезавшегося зуба имеется костный карман. Компактная пластинка в верхнем отделе межальвеолярной перегородки на…
У многих новорожденных в области затылка, на границе волосистой части на лбу и реже в области бровей имеются красные пятна различной, большей частью неправильной формы, отчетливо отграниченные от окружающей нормальной кожи. Под давлением пальца краснота исчезает, но затем сразу появляется. Это вызвано расширением кожных сосудов. Такие пятна в дальнейшем большей частью совершенно исчезают. Они имеют…
По сравнению с другими органами и тканями мышечная ткань с момента рождения до зрелости в количественном (весовом) отношении подвергается наибольшим изменениям. У новорожденного на ее долю приходится 23,3% общего веса тела, в возрасте 7—8 лет—27,2%, в 14—15 лет —32,3%, а в возрасте 17—18 лет — 44,2%. Механическая мышечная возбудимость в первые дни жизни значительно повышена….
Точных данных о возрастных особенностях локализации вкусовых сосочков у детей в современной литературе не имеется. Есть основание предполагать, что поверхность, воспринимающая вкусовые раздражения, у детей значительно больше, чем у взрослых (А. Ф. Тур). Чувство вкуса у новорожденного развито хорошо. На различные вкусовые раздражения он реагирует различно: на сладкое — сосательными и глотательными движениями и общей…
Задание 3. Билет № 8
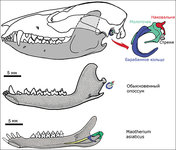
В эволюции висцерального черепа позвоночных прослеживается несколько типов сочленения нижней челюсти с черепом. На представленных схемах показаны два основных типа соединения челюстной дуги с черепом (схема прилагается).
Здесь приложена схема шести черепов. Какие будут на экзамене, неизвестно. Возможно, что этот ответ из двух билетов или более. Надо запомнить все черепа.
Эволюциядвух первых жаберных (висцеральных) дуг в ряду позвоночных:

Точками обозначены хрящи подъязычной дуги и производное гиомандибулярного хряща - слуховая косточка (стремечко); заштрихованы нёбноквадратный хрящ и меккелев хрящ и их производные - слуховые косточки (наковальня и молоточек); не заштрихованы покровные кости.
А - акула; Б - костистая рыба; В - амфибия; Г - рептилия; Д- териодонт; Е – млекопитающее.
1 - нёбноквадратный хрящ; 2 - меккелев хрящ; з - квадратная кость - наковальня млекопитающих; 4 - нёбная кость; 5 - крыловидные кости; 6 - сочленовная кость - молоточек млекопитающих; 7 - зубная кость;8 - угловая кость; 9 - гиомандибулярный хрящ - слуховая косточка (стремечко) наземных позвоночных; 10 - чешуйчатая кость; 11 - гиоидный (подъязычный) хрящ (или кость).
Перечислите основные элементы челюстной и подъязычных дуг.
Основные элементы челюстной дуги (два хряща):
- верхний: небно-квадратный – первичная в/ч.
- нижний: меккелев хрящ – первичная н/ч.
Основные элементы подъязычной дуги:
- гиомандибулярный хрящ (в гиостильном сращен с основанием черепа)
- гиоид (до рептилий соединен с меккеевым хрящем).
2.Чем характеризуется каждый тип сочленения?
Гиостильный: прикрепление через гиомандибулярный элемент - через подвесок (рыбы).
Аутостильный: небно-квадратный элемент (в/ч-ная дуга) срастается с основанием черепа. Начало: с амфибий, далее рептилии, млекопитающие, человек.
3.Для каких классов позвоночных характерны данные типы сочленения. С какими эволюционными событиями связан переход от одного типа сочленения к другому?
Филогенетически мозговой череп прошел три стадии развития: перепончатую, хрящевую и костную. У круглоротых он практически весь перепончатый и не имеет передней, несегментированной, части (протостильный тип). Череп хрящевых рыб почти полностью хрящевой, причем включает в себя как заднюю, первично сегментированную, часть, так и переднюю. У костных рыб и остальных позвоночных осевой череп становится костным за счет процессов окостенения хряща в области его основания.
Висцеральный череп впервые появляется также у низших позвоночных. Он формируется из мезенхимы эктодермального происхождения, которая группируется в виде сгущений, имеющих форму дужек, в промежутках между жаберными щелями глотки. Первые две дужки получают особенно сильное развитие и дают начало челюстной и подъязычной дугам взрослых животных. Следующие дуги в числе 4—5 пар выполняют опорную функцию для жабр и называются жаберными.
У хрящевых рыб впереди челюстной дуги располагаются обычно еще 1—2 пары предчелюстных дуг, имеющих рудиментарный характер. Челюстная дуга состоит из двух хрящей. Верхний называют нёбно-квадратным, он выполняет функцию первичной верхней челюсти. Нижний, или меккелев, хрящ — первичная нижняя челюсть. На вентральной стороне глотки меккелевы хрящи соединены друг с другом таким образом, что челюстная дуга кольцом охватывает ротовую полость. Вторая висцеральная дуга с каждой стороны состоит из гиомандибулярного хряща, сращенного с основанием мозгового черепа, и гиоида, соединенного с меккелевым хрящом. Таким образом, у хрящевых рыб обе первичные челюсти соединены с осевым черепом через вторую висцеральную дугу, в которой гиомандибулярный хрящ выполняет роль подвеска к мозговому черепу.
У костных рыб начинается замещение первичных челюстей вторичными, состоящими из накладных костей — челюстной и предчелюстной сверху и зубной внизу. Нёбно-квадратный и меккелев хрящи при этом уменьшаются в размерах и смещаются кзади. Гиомандибулярный хрящ продолжает выполнять функции подвеска, поэтому череп - гиостильный.
У амфибий(фиг. В) с выходом их на сушу (при превращении головастика в лягушку) существенному преобразованию подвергается подъязычная дуга. Ее верхний отдел (гиомандибулярная кость) теряет связь с нижним (гиоидным, или собственно подъязычным, хрящом) и приобретает новую функцию - становится маленькой слуховой косточкой (столбиком), передающей звуковые колебания внутреннему уху (рис. 2). Челюстная дуга своим верхним элементом — нёбно-квадратным хрящом — срастается полностью с основанием мозгового черепа, и череп становится, таким образом, аутостильным.
Висцеральный череп пресмыкающихся также аутостилен. Для челюстного аппарата характерна более высокая степень окостенения, чем у земноводных. Урептилий и птиц (фиг. Г) квадратная кость сохраняет значение подвески и служит для связи черепа с нижней челюстью через сочленовную косточку (преобразованный меккелев хрящ). Элемент подъязычной дуги - столбик получает здесь более сложное строение: на нем выступает стремечко, закрывающее овальное отверстие слуховой капсулы.
Нижняя челюсть млекопитающих сочленяется с височной костью сложным суставом, позволяющим не только захватывать пищу, но и совершать сложные жевательные движения. У млекопитающим (фиг. Е), мы видим, что у них сочленение нижней челюсти с черепом осуществляется совершенно иным образом, чем у других позвоночных, - уже без всякого участия остатков первой висцеральной дуги: нижние челюсти, образуемые зубными костями, сочленяются без всякого подвеска с чешуйчатыми костями черепа.Куда же девались квадратная и сочленовная кости? Исследования эмбрионального развития черепа обнаружили, что у млекопитающих они совершенно выпадают из челюстного аппарата и входят в состав слуховых косточек. Здесь квадратная кость становится наковальней, а бывшая сочленовная - молоточком (Рис. 3).Так, в среднем ухе у млекопитающих из миниатюрных остатков челюстной дуги и верхней части подъязычной образовался передаточный аппарат из трех звеньев: молоточка, прирастающего к барабанной перепонке, наковальни, занимающей промежуточное положение, и стремечка, которое закрывает собой овальное отверстие внутреннего уха. Что же касается нижней части подъязычной дуги, то она преобразуется в подковообразную подъязычную кость, подвешенную к черепу посредством пары связок.
4.В состав каких структур включаются элементы челюстной и подъязычной висцеральных дуг?
Одна слуховая косточка — столбик,— характерная для земноводных и пресмыкающихся, уменьшаясь в размерах, превращается в стремечко, а рудименты нёбно-квадратного и меккелева хрящей, полностью выходящие из состава челюстного аппарата, преобразуются соответственно в наковаленку и молоточек. Таким образом, создается единая функциональная цепь из трех слуховых косточек в среднем ухе, характерная только для млекопитающих.
- нёбно-квадратный хрящ (первичная в/ч) - превращается в наковальню.
-меккелев хрящ (первичная н/ч)- превращается в молоточек.
-гиомандибулярный хрящ, после преобразований – слуховая косточка-столбик- стремечко.
Рыбы хрящевые.
Челюстная дуга образована двумя парными хрящами. Левый и правый элементы каждой пары спереди соединяются. Функция верхней челюсти – небно-квадратный хрящ, образующий челюстной сустав с нижней челюстью – меккелевым хрящем. Подъязычная дуга лежит за челюстной. Состоит из следующих частей. Верхний парный хрящ – подвесок или гиомандибулярий верхним концом при помощи сустава и связок подвижно прикрепляется к слуховому отделу мозгового черепа, а его нижний конец подвижно сочленяется как с обоими элементами челюстной дуги. Так и с нижним парным элементом подъязычной дуги – гиоидом. Гиоиды соединяются между собой через копулу. Это было описание гиостилии. У древних акул – амфистилия – челюстная дуга прикрепляется к черепу через подвесок и, кроме того отросток небноквадратного хряща сочленяется с основанием мозгового черепа. Для скатов характерна гиостилия типичная, но гиоид редуцируется.
Рыбы костные.
Характерна гиостилия. Покровные кости укрепляют челюстную дугу, образуя вторичные челюсти. В результате окостенения первичной верхней челюсти – небноквадратного хряща – с каждой стороны образуется несущая зубы небная кость, а в задней его части – задняя крыловидная и квадратная кости. Между ними покровные наружные и внутренние крыловидные кости. Первичная нижняя челюсть – меккелев хрящ, окостеневает, и превращается в сочленовную кость, образующую с квадратной челюстной сустав. В верхней челюсти из покровных костей – предчелюстная и верхнечелюстная кости. Покровная кость нижней челюсти – зубная. Подъязычная дуга из основных костей. Подвесок соединяется со слуховым отделом осевого черепа. От подвеска отщепляется симплектикум. Через него соединение с квадратной костью. Передние концы гиоидов правой и левой сторон соединены друг с другом копулой. К гиоиду прикрепляются косточки – лучи жаберной перепонки.
Небноквадратный хрящ сохраняется в течение всей жизни. Передним и задним концами прирастает к черепной коробке – аутостилия. К небноквадратному прилегают парные покровные кости – предчелюстная и верхнечелюстная. Сзади небноквадратный хрящ покрыт квадратно-скуловой костью и чешуйчатой костью (это сверху), а снизу – крыловидной костью. На меккелевом хряще, на переднем конце подбородочно-челюстные кости. За ними, прикрывая меккелев хрящ, лежат покровные зубные кости. Задняя часть меккелева хряща обрастает покровная угловая кость. Суставным отростком меккелев хрящ соединяется с задним концом небно-квадратного хряща, образуя челюстной сустав. Подвесок превратился в слуховую косточку.
Небноквадратный хрящ дает начало квадратным костям, которые верхом связаны с мозговой коробкой, а низом с нижней челюстью. Передняя часть небноквадратного хряща заменяется покровными костями дна черепа: парными небными и крыловидными. Поперечные кости соединяют крыловидные с верхнечелюстными, а у ящериц и гаттерий еще и верхнекрыловидные связывают крыловидные с теменными. Образуется вторичное костное небо. Нижняя челюсть. Вместо меккелева хряща – сочленовная, соединенная с квадратной костью, зубная, угловая, надугловая, венечная. Гиомандибуляре превращен в стремечко.
Верхнечелюстные кости сливаются с небными костями своими отростками. К задненаружному краю верхнечелюстной кости прирастает тонкая костная перекладина, состоящая из скуловой и квадратной скуловой кости. Квадратная скуловая кость сочленяется с квадратной костью, нижний конец которой образует суставную поверхность для сочления с нижней челюстью, а длинный верхний конец суставом прикрепляется к чешуйчатой и переднеушной костям. Небные кости концами налегают на клювовидный отросток парасфеноида и суставом соединяются с парными крыловидными костями, которые связаны с квадратными костями. Такое строение костного неба характерно для кинетизма. Нижняя часть клюва образуется слиянием костей – зубной, сочленовной и угловой. Челюстной сустав формируют сочленовная и квадратная кости.
Млекопитающие.
Череп синапсидного типа. Он обладает скуловой дугой, образованной костями: верхнечелюстная – скуловая – чешуйчатая. Нижняя челюсть образована всего из одной кости – зубной, которая причленяется к скуловому отростку чешуйчатой кости. Суставная кость нижней челюсти рептилий, уменьшаясь в величине, превращается в одну из косточек среднего уха – молоточек. Другая часть среднего уха образована квадратной костью, превращающейся в наковальню.
Висцеральная часть черепа составлена несколькими костями. В составе вторичной верхней челюсти спереди расположены небольшие предчелюстные кости, позади которых находятся верхнечелюстные. Небные отростки этих костей образуют для всех млекопитающих твердое костное небо. Участвуют в его формировании также покровные небные кости. К их задним выступам прирастают крыловидные кости. К первичной верхней челюсти относятся замещающие крыло - клиновидная и наковальня в среднем ухе которая.
Читайте также: