
Характеристика отношений типа хищник жертва паразит хозяин
Читайте также:
|








| Поделиться: |
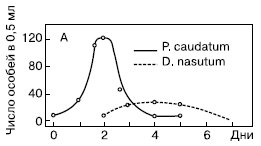
Хищник—жертва (взаимоотношения)Взаимоотношения хищников и их жертв. Прямые трофические связи лежат в основе и этой формы биоценогаческих отношений. И в этом случае совместная эволюция трофически связанных популяций приводит к выработке противоположно направленных коадаптаций: у жертв они способствуют снижению пресса хищников, а у последних — повышению результативности охоты даже с учетом противодействующих приспособлений жертв.[ . ]
Важнейший вид взаимоотношений между организмами в биоценозе, фактически формирующими его структуру, — это пищевые связи хищника и жертвы: одни — поедающие, другие — поедаемые. При этом все организмы, живые и мертвые, являются пищей для других организмов: заяц ест траву, лиса и волк охотятся на зайцев, хищные птицы (ястребы, орлы и т. п.) способны утащить и съесть как лисенка, так и волчонка. Погибшие растения, зайцы, лисы, волки, птицы становятся пищей для детритофагов (редуцентов или иначе деструкторов).[ . ] Хищничество — взаимоотношения, при которых один из участников (хищник) умерщвляет другого (жертва) и использует его в качестве пищи. Например, волки и зайцы.[ . ] Хищничество — взаимоотношения, при которых один из участников (хищник) умерщвляет другого (жертва) и использует его в качестве пищи. Например, волки и зайцы. Состояние популяции хищника тесно связано с состоянием популяции жертв. Однако при сокращении численности популяции одного вида жертв хищник переключается на другой вид. Например, волки могут использовать в качестве пищи зайцев, мышей, кабанов, косуль, лягушек, насекомых и т.д.[ . ] Отсюда вопрос о взаимоотношениях хищников и жертв представляется не той схематически простой системой, которая легла в основу математического анализа Лотка и особенно Вольтерра, поскольку даже при наличии максимально однородного материала индивидуальные особенности отдельных пищевых объектов небезразличны для животного-потребителя.[ . ] Полной защиты от хищников всех особей в популяциях жи-вотных-жертв не происходит, так как это привело бы не только к гибели голодающих хищников, но в конечном итоге — к катастрофе популяций жертв. Ухудшается их генофонд (сохраняются больные и старые животные) и ввиду резкого увеличения численности подрывается кормовая база. Равновесию В.х.-ж. способствует наличие в пищевой цепи третьего звена (крупный хищник, паразит). ВЗАИМОПОМОЩЬ РАСТЕНИЙ (В.р.) — взаимоотношения растений типа комменсализма и протокооперации. В результате В.р. растения, высеянные группой, развиваются лучше, так как у них легче формируется мутуализм с грибами и бактериями. Достаточно обычны случаи срастания корней у деревьев, при этом часть пластических веществ переходит от более сильного растения к более слабому. Однако при достижении растениями определенного возраста положительный эффект группы сменяется конкуренцией, и растения в центре группы развиваются хуже, чем расположенные по краю.[ . ] Простейшая модель взаимоотношений популяций хищника и жертвы, основанная на логистическом уравнении роста, названа (как и модель межвидовой конкуренции) по имени ее создателей - Лотки и Вольтерры. Эта модель крайне упрощает исследуемую ситуацию, но все же полезна в качестве отправной точки в анализе системы хищник-жертва.[ . ] Пример отрицательной (стабилизирующей) обратной связи — взаимоотношение между хищником и жертвой или функционирование карбонатной системы океана (раствор СОг в воде: СО2 + Н2О —> Н2СОз). Обычно количество углекислоты, растворенное в воде океана, находится в парциальном равновесии с концентрацией углекислого газа в атмосфере. Локальные увеличения углекислоты в атмосфере после извержения вулканов приводят к интенсификации фотосинтеза и поглощению ее карбонатной системой океана. При снижении уровня углекислого газа в атмосфере карбонатная система океана высвобождает СОг в атмосферу. Поэтому концентрация углекислого газа в атмосфере достаточно стабильна.[ . ] Как отмечает Р.Риклефс (1979), существуют факторы, способствующие стабилизации взаимоотношений в системе “хищник-жертва”: неэффективность хищника, наличие у хищника альтернативных пищевых ресурсов, уменьшение запаздывания в реакции хищника, а также экологические ограничения, налагаемые внешней средой на ту или иную популяцию. Взаимодействия между популяциями хищника и жертвы весьма разнообразны и сложны. Так, если хищники достаточно эффективны, они могут регулировать плотность популяции жертвы, удерживая ее на уровне ниже емкости среды. Через влияние, оказываемое ими на популяции жертвы, хищники воздействуют на эволюцию различных признаков жертвы, что приводит в конечном итоге к экологическому равновесию между популяциями хищника и жертвы.[ . ] Применение показателя х позволяет оценить природу одного из существенных элементов взаимоотношения между хищником и жертвами.[ . ] Итак, наши исследования показали, что индивидуальное пространственное поведение и территориальные взаимоотношения с конспецифи-ческими особ яки у личинок С. эр1епс1епв сложнее, чем можно было ожидать от сидячего подстерегающего хищника, использующего лишь тактильную сенсорную систему в процессе охоты. Поведение названных личинок наряду с другими экологическими факторами имеет значение в формировании структуры популяции этого хищника и в его взаимоот>-ношениях с жертвами. В частности, поведение в какой-то степени влияет на распределение в пространстве мест долговременного пребывания особей и в связи с этим определяет плотность и границы обитания популяции. Передвижения особей, коррелирующие по частоте и дальности с плотностью популяции, обеспечивают контакт с жертвами во всем местообитании хищников, даже при невысокой их плотности. Данные о способности личинок С. Бр1еп6епэ к ориентированию заставляют думать о значительно больших, чем предполагалось, возможностях их сенсорных систем, по-видимому, используемых в передвижениях и охоте.[ . ] Поскольку направление сдвига и степень асимметрии кривых распределения показателей элективности характеризуют взаимоотношения между хищниками и жертвами, представляет значительный интерес найти количественный способ выражения этой асимметрии.[ . ] Число трофических связей и пищевая конкуренция были минимальными и эффект стабильности в этой системе поддерживался взаимоотношениями единственного хищника — балхашского окуня, имевшего высокую численность, с его жертвами, которыми были практически все виды (в основном их молодь), обитающие до среднего течения рек.[ . ] Нередко польза и вред бывают односторонними. Для льва безразлично, поедают ли грифы и шакалы остатки его пищи (,комменсализм); для жуков-навозников несущественно, что в полете они переносят нематод-копрофагов к новым навозным кучам — их субстрату. При паразитизме и хищничестве один из партнеров извлекает для себя пользу во вред другому. Эти два типа взаимоотношений, как уже было отмечено ранее, различаются тем, что в первом случае нападающий организм меньше своей жертвы, а во втором — крупнее. Репродуктивный потенциал, как правило, у паразита больше, чем у хозяина, а у хищника — меньше, чем у жертвы.[ . ] Скорость биологических круговоротов и общее количество вовлекаемого в эти циклы вещества определяются масштабами и экологическими условиями в экосистемах. Для экосистем характерны различные экологические условия, под которыми подразумеваются экологические факторы внешней среды, прямо или косвенно воздействующие на живые организмы. Эти факторы могут быть абиотическими и биотическими.. Абиотическими (ракшрами являются климат, рельеф местности, почва, свет, тепло, вода, воздух, снеговая нагрузка и др. Биотические факторы порождаются взаимоотношениями организмов: конкуренцией, взаимоотношениями типа "хищник-жертва", "хозяин-паразит", симбиоз и др.[ . ] Конкуренцию принято считать главным организующим фактором сообщества как по причине ее воздействия на размеры ниши, так и на основе определения ниши, данного Хатчинсоном [195]. Это в известной мере вводит в заблуждение, поскольку альтернативным механизмом потенциально может быть и хищничество. Однако хищничеству нельзя отводить главную роль ни в непосредственном определении видового состава (в пользу этого данных очень мало), ни в предотвращении конкурентного исключения (соответствующие данные относятся главным образом к низшим трофическим уровням и к регуляции разнообразия среди сидячих организмов, конкурирующих за пространство). Возможно, что в некоторых случаях хищничество служит фактором, непосредственно регулирующим видовое богатство, но конечным механизмом, создающим структуру сообщества, все же остается конкуренция Даже в самых эффектных примерах регуляции видового богатства хищниками, а именно в сообществах приливно-отливной зоны, влияние хищничества просто накладывается на основу, созданную конкурентными взаимоотношениями среди жертв.[ . ] Лишь несколько работ отличаются системным подходом к изучению трофических связей имаго стрекоз, хотя они и не лишены некоторых методических недостатков. Так, в статье по стрекозам агробиоценозов сои во Флориде / Neal, Whitcomb, 1972/ проанализирован видовой состав стрекоз с одновременной регистрацией времени лета, характерных особенностей питания и рациона. Всего в агробиоценозах сои отмечено 35 видов стрекоз, 9 из которых были массовыми и потенциально могли участвовать в регуляции численности от дельных групп вредителей. Однако полевые наблюдения показали, что только три из них (Erythemis simplicicollis, Pachydiplax longipennis и Perithemis tenera seminole ) оказывали влияние на численность некоторых экономически важных вредителей. Но авторы отмечают, что все эти виды активно питались и в других биоценозах. Лишь одна равнокрылая стрекоза Anomalagri— on hastatum была постоянно связана с культурой сои. Какие-либо количественные данные по оценке экономической значимости стрекоз в биоценозах сои в этой работе отсутствуют, осталось неясным их место в системе "хищник — жертва", взаимоотношения и сосуществование с другими хищниками.[ . ] Читайте также:
|

