
Какая плодовитость у паразитов
Высокая плодовитость паразитических животных обеспечивается разными способами.
Мы уже упоминали о роли гермафродитизма, при котором все особи популяции дают потомство. Как приспособление к перекрестному оплодотворению для паразитов характерен протеандрический гермафродитизм, когда вначале формируются мужские половые органы, а затем женские. Протерогенический гермафродитизм, т. е. развитие женской половой системы раньше мужской, крайне редок.
При функциональном гермафродитизме животные продуцируют преимущественно один род половых клеток и лишь время от времени другой, либо последовательно выступают в роли одного или другого пола, выполняют функции самцов и самок одновременно. Пример функционального гермафродитизма дает Danalia curvata.
Для одноклеточных животных многочисленное потомство обеспечивается бесполым размножением при непродолжительном периоде индивидуальной жизни клетки, а также способностью к множественному бесполому размножению – шизогонии, наблюдаемому у споровиков. Метагенез у простейших способствует не только увеличению их количества, но и благодаря половому способу размножения обеспечивает повышение уровня их жизнеспособности (шизогония, гаметогония и спорогония у Plasmodium и Eimeria).
Важное значение имеет гипертрофия половой системы у паразитов (Trematoda ), а также умножение числа половых комплексов (Cestoda). Причем увеличение числа половых аппаратов происходит не только в повторении их в каждом членике, но и в развитии двух половых систем во всех члениках (Dipylidium caninum) стробилы.
Гипертрофия половой системы встречается и у паразитических нематод. Так, у Sphaerularia bombi, паразитирующей в полости тела шмелей, развившаяся матка превосходит по объему все остальное тело нематоды в 15-20 тыс раз.
Повышению плодовитости, значительному увеличению числа потомков способствует включение в жизненные циклы паразитов различных дополнительных способов размножения.
У эхинококка из попавшего в различные органы (чаще в печень и легкие) шестикрючного зародыша развивается пузырчатая финозная стадия, образующая на внутренней стенке многочисленные капсулы с большим количеством головок и дочерние пузыри, соответствующие ценурам. В средней величины эхинококке образуются в результате бесполого размножения, или почкования, многие тысячи головок. Причем в случае разрыва эхинококка часть сколексов может давать вторичные пузыри, увеличивая степень поражения хозяина. Подобное почкование финозных стадий наблюдается в цикле развития мозговика (Multiceps) и некоторых других цестод.
У дигенетических сосальщиков помимо полового размножения гермафродитной мариты в цикле развития происходит партеногенез. Из мирацидия, проникшего в моллюска, развивается партеногенетическая спороциста, имеющая вид простого мешка. Спороциста – половозрелая особь (партенита). В задней части ее тела находятся зародышевые клетки, из которых у разных трематод развиваются или спороцисты второго, затем третьего порядка, или редии. Редии отличаются от спороцисты подвижностью, наличием мешковидного кишечника и отверстия на переднем конце тела, через которое из редии выходит новое поколение зародышей, развившихся из партеногенетических яиц внутри редии. Таким образом, у трематод в цикле развития закономерно чередуются половое размножение гермафродитной мариты и партеногенез спороцист и редии, т. е. имеет место гетерогония.
Гетерогония присутствует и в жизненном цикле моногеней, в частности, у гиродактилид. В матке гигодактилюса из оплодотворенного яйца развивается зародыш, внутри которого на ранних стадиях его развития закладывается эмбрион следующего поколения. В нем в свою очередь формируется зародыш третьего поколения, из обособленной клетки которого в дальнейшем развивается эмбрион четвертого поколения. В результате образуется система из четырех как бы вложенных друг в друга зародышей, находящихся на разных стадиях развития.
Знакомство с циклами развития паразитов показывает, что чем сложнее жизненные циклы, чем больше препятствий возникает в развитии и расселении паразитов, тем большим должно быть их потомство. Это свойство паразитов получило название Закона большого числа яиц, или Закона множественного размножения. Следует однако заметить, что высокая плодовитость присуща отнюдь не только паразитическим животным, но и некоторым свободноживущим. Так, рыбы, обитающие в морях в связи с наружным оплодотворением отличаются большой плодовитостью: сельдь ежегодно выметывает до 30 тыс. икринок, треска – 9 млн., в яичнике луны-рыбы насчитывается до 300 млн. яиц. Самка термитов за 10 лет жизни продуцирует 100 млн. яиц, или откладывает 1 яйцо каждые 2-3 сек.
Данные о подолжительности жизни некоторых распространенных паразитов и их плодовитости сведены в таблице 3.
Продолжительность жизни и плодовитость паразитических животных
Защитные приспособления паразитов
Полупаразиты
Широко известны луговые травянистые полупаразиты. Среди них погремки большой и малый, мытник, очанки и др. К корням травянистых растений они прикрепляются с помощью гаусториев. Полупаразитный образ жизни ведут омела белая и ремнецветник европейский, поселяющиеся на ветвях древесных пород, таких, как липа, тополь и др. Всего насчитывают 1964 вида полупаразитов.
Для паразитов выход во внешнюю среду, как правило, чреват многими опасностями, отсюда на той стадии жизненного цикла, которую паразиты проводят вне хозяина, у них развиваются различные защитные приспособления, позволяющие пережить этот критический период: толстые и многослойные оболочки яиц гельминтов,
цисты кишечных амеб,
ооцисты со спорами кокцидий,
способность к анабиозу у ряда личинок нематод и т. д.).
Основные экологические трудности, с которыми сталкиваются внутренние сожители живых организмов, — это ограниченность жизненного пространства для внутриклеточных и тканевых обитателей, сложности снабжения кислородом, трудность распространения от одной особи хозяев к другим, а также защитные реакции организма хозяина против паразитов.
Ограниченность жизненного пространства особенно сказывается на размерах и форме внутриклеточных паразитов. Так, грегарины, живущие в полости кишечника, — крупные споровики со сложно расчлененной клеткой, тогда как малярийные плазмоиды, являющиеся внутриклеточными паразитами, отличаются мелкими размерами и упрощенным внешним строением.
Недостаток кислорода в тканях и, особенно в желудочно-кишечном тракте организмов-хозяев приводит к тому, что у многоклеточных обитателей внутриорганизменной среды вырабатывается преимущественно анаэробный тип обмена. Необходимая для работы клеток энергия высвобождается за счет разных видов брожения, а не за счет дыхания. Так, у человеческой аскариды утрачены все ферменты дыхательного цикла.
Однако целый ряд паразитов не утрачивает полностью способности к дыханию и может переключаться с анаэробного типа обмена на аэробный, таковы жгутиковые эхинококк и др.
Важная адаптация паразитов — синхронизация их жизненных циклов с сезонным развитием растений-хозяев, позволяющая осуществить заражение хозяина в нужный момент. Это явление широко известно для многих паразитических грибов. Например, слишком рано появившиеся проростки повилики прекращают рост, иногда на несколько недель, до тех пор, пока рядом не разовьются проростки травянистых растений — возможных хозяев.
Существенными адаптациями паразитов являются повышенная способность к размножению, выработка сложных жизненных циклов, использование переносчиков и промежуточных хозяев. Например, полупаразит колдунья трава широко распространен в Африке, Южной Азии, Австралии на зерновых культурах, сорго, сахарном тростнике, образует на одном растении до 0,5 млн. очень мелких семян.
У ряда паразитов приспособления к умножению потомства проявляются и в виде партеногенеза, полиэмбрионии (клетки одного делящегося яйца дают начало множеству зародышей), бесполого размножения(почкование у пузырчатых стадий ленточных червей). Это приводит к чередованию поколений — полового и партеногенетического или полового и бесполых.
У многих паразитов чередование поколений сочетается со сменой двух или более хозяев, следовательно, одно поколение существует в одном хозяине, а другое — в другом. К примеру, ленточный червь, обитающий в кишечнике лисицы, откладывает там крошечные яички. Эти яички вместе с экскрементами лисицы попадают на землю. Как им теперь вернуться в кишечник какой-нибудь другой лисицы? Процесс возвращения включает в себя стадию промежуточного хозяина или несколько хозяев. Траву, на которую попали яйца ленточного червя, съедает заяц. В его организме яйца превращаются в личинки, они вбуравливаются в ткани зайца и там переходят в покоящуюся стадию — цисты. В дальнейшем, если лисица поймает этого зайца, то цисты попадут в ее кишечник, там превратятся в молодого ленточного червя, и жизненный цикл начнется сначала. Многообразие и сложность жизненных циклов паразитов выработались как приспособление для передачи от одной особи хозяина к другой, для распространения.
Адаптация паразита к существованию на определенном хозяине включает также весьма тонкую биохимическую специализацию — выработку определенного набора ферментов, облегчающих проникновение в тело хозяина и использование поступающих от него веществ.
В ряде случаев паразиты сами становятся средой обитания других видов — возникает явление сверхпаразитизма, или гиперпаразитизма. Например, для паразита капустной белянки наездника известно более 20 видов вторичных паразитов из перепончатокрылых. Явления сверхпаразитизма в растительном мире сравнительно редки. К ним относятся случаи поселения одного вида омелы на другом в тропических лесах Индии.
В разных частях тела многоклеточного организма условия неоднородны. Его паразиты специализируются к жизни в определенных органах и тканях. Они приурочены к определенному возрастному и физиологическому состоянию хозяина. Так, в волосах человека обитают одни разновидности вшей, тогда как в волосяном покрове других частей тела — совсем иные. В пищеварительном тракте кролика одновременно могут паразитировать несколько видов кокцидий, каждый из которых локализуется в определенных частях кишечника: в начальном отделе тонкой кишки, в средней, в последней ее петле, преимущественно в слепой кишке и т. д.
В Сибири, в лиственницах, тонкоусый еловый усач заселяет преимущественно прикомлевую часть до высоты примерно 1 м, лиственничная златка осваивает ствол выше, до 4—5 м, продолговатый короед распространяется по всей его средней части, а вершину и ветви заселяют короед-гравер и заболотник Моравица.
Большое число паразитов обитает не внутри, а на поверхности тела хозяина, выступающего в этом случае как часть внешней среды паразита, снабжая его пищей, предоставляя убежище, трансформируя микроклимат. Связь с хозяином эктопаразита может быть постоянной или временной. Одна из основных жизненно важных экологических задач эктопаразитов— удержаться на теле хозяина с помощью мощных органов прикрепления — присосок, крючьев, коготков и т. д., которые развиваются у самых разных видов.
Нам важно ваше мнение! Был ли полезен опубликованный материал? Да | Нет
Site biology teachers lyceum № 2 Voronezh city, Russian Federation
Взаимоотношения между организмами. Паразитизм
Паразитизм – это способ питания за счет питательных веществ другого организма (хозяина), причем последний от этого не погибает, но чувствует себя угнетенно. В мертвом теле хозяина паразиты не живут. Имеются виды организмов, которые паразитируют на других организмах, но способны одновременно и сами добывать пищу. Такие организмы называют полупаразитами. Например, растения омела, марьянник, мытник, паразитируя на других растениях, одновременно сами осуществляют фотосинтез.
Изучением паразитов и явления паразитизма, во всех его проявлениях, а также разработкой методов борьбы с паразитами и болезнями, вызываемыми паразитами, занимается наука паразитология .
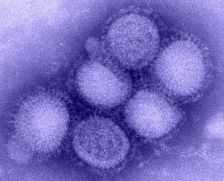
Вирусы — простейшая форма жизни, микроскопические частицы, представляющие собой молекулы нуклеиновых кислот (обычно только один тип: ДНК или РНК), заключённые в белковую оболочку и способные инфицировать живые организмы.
Вирусы являются обязательными паразитами, так как не способны размножаться вне клетки хозяина. В настоящее время известны вирусы, размножающиеся в клетках растений, животных, грибов и бактерий (последних обычно называют бактериофагами).
Впервые существование вируса, как нового типа возбудителя болезней, доказал в 1892 году русский учёный Д. И. Ивановский.
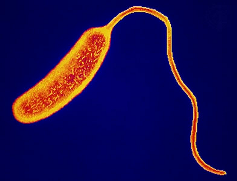
Паразитические бактерии. Среди бактерий, как и среди других таксономических групп живых организмов, встречается довольно большое количество видов, ведущих паразитический образ жизни. В процессе своей жизнедеятельности такие бактерии вырабатывают токсины — ядовитые вещества, воздействующие на определенные системы органов, которые вызывают характерный комплекс симптомов, по которым можно диагностировать болезнь и установить её возбудителя.
У человека паразитические бактерии вызывают туберкулёз, дифтерию, ангину, дизентерию, чуму, холеру и другие заболевания.
Грибы-паразиты используют в качестве источника питания живые ткани различных организмов. Они являются возбудителями заболеваний растений, животных и человека.
Различают грибы, паразитирующие на животных, — зоопатогенные ; на растениях — фитопатогенные , которые являются наиболее распространёнными среди паразитических грибов. Они воздействуют токсическими выделениями или ферментами на ткани растений, а затем используют их для питания. Многие из фитопатогенных грибов поражают хозяйственно ценные растения. Известна большая группа древоразрушающих грибов, развивающихся как на растущих деревьях (разные виды трутовиков), так и на деревянных частях построек.
Растения-паразиты — растения, полностью или частично живущие за счёт питательных веществ хозяина. Они встречаются как среди низших, так и среди высших растений, в том числе цветковых. Цветковые растения паразитируют главным образом на высших растениях.
Связь с растением-хозяином паразит осуществляет через гаустории — многоклеточные образования, возникающие в результате преобразования зародышевого корня или в редких случаях тканей стебля. Гаустории эктопаразитов внедряются в ткани организма-хозяина и извлекают из них питательные вещества. Эндопаразиты полностью или главным образом развиваются в тканях растения-хозяина и выходят на поверхность его тела только для размножения.
Растения-полупаразиты, в отличие от паразитов, полностью сохранили фотосинтетическую функцию листьев. В связи с этим, так же как насекомоядные растения, они по типу питания автотрофны. Вместе с тем полупаразиты характеризуются слабым развитием корневой системы и почти полным отсутствием корневых волосков. С помощью корневых присосок они получают воду и питательные вещества из корней растения-хозяина. В некоторых случаях полупаразиты полностью лишены корней и присасываются к побегам.
Гнездовой паразитизм — тип паразитизма, при котором паразит использует хозяина, которым может являться особь как своего (внутривидовой паразитизм), так и другого вида (межвидовой паразизм), с целью выращивания своего потомства. В результате такого взаимодействия паразит избавлен от необходимости строить гнездо и выкармливать потомство, а может тратить больше времени на добычу пищи и размножение.

Гнездовой паразитизм встречается у птиц, рыб, насекомых, но в целом является малораспространённым явлением.

Особые приспособления паразитов. Особые условия среды обитания паразитов: защищённость от внешних врагов, обилие легкоусвояемой пищи, относительная стабильность условий — делают ненужной сложную дифференцировку тела, и поэтому многие внутренние паразиты характеризуются в эволюции вторичным упрощением строения, вплоть до потери целых систем органов. Так, ленточные черви , всасывающие переваренную хозяином пищу через покровы, отличаются отсутствием пищеварительной системы и редукцией нервной.

Подобно животным паразитам упрощается и строение растений-паразитов: утрачиваются вегетативные органы и способность к фотосинтезу. У раффлезии на поверхности земли образуются лишь отдельные цветки, а вегетативное тело представлено клеточными нитями, подобными гифам гриба, и полностью погружено в ткани питающего растения.
В случае, если часть жизненного цикла паразита проходит вне организма хозяина, а во внешней среде, у него развиваются различные защитные приспособления, позволяющие пережить этот период. Если в жизненном цикле паразитов нет стадии выхода во внешнюю среду, как, например, у малярийного плазмодия, то таких защитных приспособлений не обнаруживается.
Недостаток кислорода в тканях и особенно в желудочно-кишечном тракте организмов-хозяев приводит к тому, что у многоклеточных эндопаразитов вырабатывается преимущественно анаэробный тип обмена.
К важным приспособлениям паразитов относятся также повышенная способность к размножению, выработка сложных жизненных циклов, использование переносчиков и промежуточных хозяев. Эти приспособления связаны с тем, что среда обитания паразитов ограничена как во времени, так и в пространстве.

Большой круг паразитов обитает не внутри, а на поверхности тела хозяина. Для постоянных или длительно связанных с хозяином эктопаразитов одна из основных жизненно важных задач — удержаться на теле хозяина. В связи с этим типичные эктопаразиты обычно характеризуются наличием мощных органов прикрепления — присосок, крючьев, коготков, которые независимыми путями развиваются у самых разных по происхождению видов.
Коготки на лапах блох обеспечивают прикрепление и передвижение насекомых в шерсти животных

Высокая плодовитость паразитов является их важной приспособительной особенностью. Одним из главных преимуществ паразитов является их обильное снабжение пищей за счёт содержимого клеток, соков и тканей тела хозяина или содержимого его кишечника. В результате этого они способны к быстрому росту, а кром е того, легкодоступная пища служит условием высокого потенциала их размножения, которое обеспечивает им вероятность заражения других хозяев.
Естественный отбор привёл к развитию у паразитов гигантской яйцепродукции по сравнению со свободноживущими формами. У ряда паразитов число яиц достигает нескольких сотен тысяч и даже миллионов. Причина столь высокой плодовитости кроется в том, что на ранних стадиях развития под действием биотических и абиотических факторов большое количество яиц паразитов гибнет. Только чрезвычайная плодовитость увеличивает шансы на выживание и завершение жизненного цикла хотя бы немногих потомков, поддерживая существование вида.

Временные паразиты всё своё развитие совершают вне тела хозяина и нападают на него в основном для питания.
Постоянные паразиты проходят все стадии жизни или большую их часть, от момента рождения до смерти, на поверхности или внутри тела хозяина.
Наружные паразиты, или эктопаразиты, — паразитические организмы, временно или постоянно обитающие на поверхности тела хозяина. Некоторые из них неподвижно прикреплены к хозяину, большинство же передвигается по телу хозяина. Основным приспособлением к существованию на хозяине являются разнообразные и часто сложно устроенные органы прикрепления — присоски, присасывательные диски, крючья и т. п.
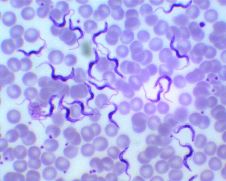
Внутренние паразиты, или эндопаразиты, — паразитические организмы, обитающие во внутренних органах животных и растительных организмов, в том числе и внутри их отдельных клеток. В последнем случае имеет место внутриклеточный паразитизм. Для многих эндопаразитов характерны сложные жизненные циклы, сопровождающиеся сменой хозяев.
Трипаносомы — паразитические одноклеточные простейшие, вызывающие многие заболевания, среди которых сонная болезнь.
Биологический метод борьбы с вредителями основан на использовании их естественных врагов — различных насекомоядных птиц, некоторых млекопитающих, хищных и паразитических насекомых, паразитических нематод и болезнетворных микроорганизмов, что позволяет поддерживать численность вредителей на низком уровне, а не приводит к их полному уничтожению. Таким образом, не происходит нарушений в функционировании экологических систем.

Основное направление в биологической защите от вредителей — использование хищных и паразитических насекомых-энтомофагов против вредных насекомых; акарифагов — против клещей. Среди хищных насекомых широко известны различные виды жужелиц, которые уничтожают яйца гусениц, взрослых особей многих садовых вредителей. Личинки златоглазки поедают яйца и гусениц чешуекрылых, тлей и медяниц. Очень полезны личинки и взрослые особи божьих коровок, которые питаются тлями, медяницами, щитовками, яйцами и гусеницами бабочек.
Другое ведущее направление в биологической борьбе с вредителями — микробиологический метод. Как и все другие живые организмы, насекомые-вредители могут поражаться различными болезнями, вызываемыми бактериями, грибами, вирусами. На основе этих микроорганизмов изготавливают препараты, уничтожающие многих насекомых-вредителей лесов и плодово-ягодных культур.
Помимо этого существуют ещё несколько способов:
Природные средства химической борьбы — это выделенные, идентифицированные, синтезированные гормоны и феромоны насекомых, используемые для прерывания их жизненного цикла. Преимущества природных химикатов заключаются в их высокой специфичности для вредителей, а также в том, что они не токсичны.

Наездники — одна из самых разнообразных групп насекомых, включающая десятки тысяч видов. Они играют огромную роль в природных экосистемах, а также в сельском хозяйстве, контролируя численность растительноядных насекомых, в том числе многих вредителей сельскохозяйственных культур.
Личинки наездника окуклились на выеденной ими изнутри гусенице бабочки.

Наездники развиваются, как паразиты, на различных хозяевах, которыми могут быть многие насекомые. Внешним видом они напоминают ос и в большинстве случаев имеют тёмную или пёструю окраску. Яйца наездник откладывает на насекомое, его личинку, гусеницу или в их тела с помощью длинного тонкого яйцеклада, которым молниеносно протыкает тело жертвы, сидя на ней верхом. В случае если яйца были отложены в гусеницу, после вылупления личинок оболочка её тела трескается, выпуская личинок наездника, которые питаются за счёт тканей хозяина, тем самым высасывая его изнутри. Личинки наездника затем превращаются в куколки, в то время как хозяин погибает.
Размножение гельминтов
Образ жизни паразитических червей обусловил определенную атрофию органов чувств и нервной системы, явился причиной развития своеобразного пищеварения. Однако, по мнению ряда исследователей, такая атрофия некоторых органов не может считаться признаком деградации животного: это только приспособление к паразитическому образу жизни.
Как ни велико упрощение, которое претерпел паразит в результате регрессивных изменений своего организма, его индивидуальное существование, даже в крайних случаях, является вполне гарантированным, если только у него достаточно развиты органы прикрепления, и он не может быть оторван от своего хозяина. Разумеется, что в тех случаях, когда паразит утрачивает органы передвижения, существование его становится в полную зависимость от жизни хозяина. Но это обыкновенно не является для него невыгодным обстоятельством, поскольку продолжительность жизни хозяина в среднем всегда больше, чем паразита.
Одно из наиболее специфических приспособлений гельминтов — хорошо развитая половая система. Если вспомнить, о тысячах смертельных опасностей, грозящих яйцам гельминтов и их личинкам, то невольно встает вопрос: почему большинство паразитических червей с беззаботным упрямством выбрасывают яйца во внешнюю среду?
Количество выделенных во внешнюю среду яиц определяется не случайностью и не физическими возможностями паразитов. Это сложилось под воздействием многих факторов, которые стоит перечислить и рассмотреть.
Возьмем для примера ленточных червей. Каждый их членик производит огромное количество яиц. В каждом яйце находится зародыш. Таких зародышей в яйцевых оболочках в каждом зрелом членике в зависимости от вида гельминтов — от нескольких десятков до нескольких сотен тысяч. Подсчитано, что каждый членик бычьего цепня содержит до 175 тыс. яиц. Человек, зараженный этим паразитом, в течение года выделяет 440 млн. яиц.
Функция размножения у большинства гельминтов является доминирующей. Кроме обычных способов размножения, у них возникают дополнительные (явление, которое часто встречается у свободноживущих организмов).
Большинство гельминтов переходят от промежуточного хозяина к окончательному. Последний поедает первого вместе' с заключающимися в нем зародышами паразитов.
Во время странствования гельминта от промежуточного хозяина к окончательному огромное количество молодых зародышей гибнет от различных случайностей: одни — от врагов, другие — от того, что не находят себе благоприятного пристанища для дальнейшего развития, и только немногие попадают в надежные условия. Так, широкий лентец за всю жизнь выделяет 11 млрд. яиц, а в организм человека попадают и развиваются до взрослой формы, как правило, один или два его потомка. Не будь у широкого лентеца столько препятствий для продолжения своего рода, количество выделенных им яиц, несомненно, было бы го- раздо меньшим.
От чего зависит большая плодовитость паразитических червей? От способа попадания в организм хозяина. В течение суток анкилостома, или кривоголовка, выделяет около 9 тыс. яиц, а человеческая аскарида — до 250 тыс. Поскольку анкилостома живет 5-6 лет, одна самка в течение жизни может отложить 25-30 млн. яиц. Вместе с тем у личинки анкилостомы, способной двигаться во внешней среде, нападать на хозяина и активно в него вбуравливаться, возможностей проникать в организм хозяина больше, чем у аскариды. Кроме того, надо учесть, что личинки анкилостомы живут около двух месяцев, а яйца аскарид — до 10 и более лет. Человеческая аскарида за 5-6 месяцев своей половозрелой жизни продуцирует около 50-60 млн. яиц.
Еще Р. Лейкарт, приводя эти цифры, отмечал, что если бы женщина сравнилась с аскаридой по плодовитости, т. е. если бы она затрачивала на воспроизведение своего потомства такое же количество материала, сколько его затрачивает самка аскариды, то она должна была бы ежедневно рожать по 70 детей. По приблизительному подсчету вес яиц, отложенных самкой человеческой аскариды за год, в 1700 раз превышает ее собственный вес. Для сравнения отметим, что пчелиная матка (одна из самых плодовитых форм) производит в год такое количество яиц, которое лишь в 100 раз превышает ее собственный вес.
Высокую плодовитость принято считать одной из самых типичных черт паразитом, и ее обычно снизывают с высокой смертностью во время расселении. Однако точные оценки практически получить очень трудно. По расчетам, значение числового фактора заражения находится в прямой зависимости от плодовитости гельминта, интенсивности выделения инвазионных яиц и их жизнеспособности. Так, если цепь поколений самок аскарид (по одной в каждом поколении) заражает внешнюю среду 125 млн. яиц, то аналогичная цепь поколений самок детских остриц заражает внешнюю среду 12 тыс. яиц.
Отмеченное несоответствие объясняется экологическими отношениями гельминтов как с открытой внешней средой, так и с организмом хозяина. Яйца аскарид выбрасываются во внешнюю среду вместе с фекалиями хозяина, и их контакт с другими индивидами хозяина маловероятен.
Самки остриц, содержащие зрелые яйца, активно выползают из анального отверстия человека и откладывают яйца на поверхности его тела. Яйца достигают способности заражать через несколько часов. В отличие от яиц аскарид яйца остриц сохраняют контакт с телом человека, с его одеждой, предметами обихода и жилищем, вероятность попадания в пищеварительный тракт другого или этого индивида хозяина соответственно выше.
Вследствие огромной продуктивной способности у гельминтов резко увеличены размеры половых органов. Так, у трематод они занимают большую часть тела. У ленточных червей они не только заполняют почти весь членик, но и с разделением тела на членики повторяются в каждом из них. Процесс увеличения размеров половых органов у некоторых видов гельминтов может заходить так далеко, что весь организм превращается в едва заметный придаток своей собственной половой системы. Например, у нематоды сферулярии, паразитирующей в полости тела шмелей, разрастающаяся матка выпадает через половое отверстие наружу, превосходя по объему все остальное тело в 15-20 раз.
Такое соотношение мы не встречаем у свободноживущих организмов, оно возможно лишь у паразитов. Здесь один-единственный орган вырастает значительно больше всего тела, отделяется от него и находит в организме другого животного те условия для питания и роста, которые должен был бы ему создать его собственный организм.
Паразитический образ жизни, прикованность гельминтов к своим хозяевам затрудняют встречу между особями разного пола. Это привело к тому, что у гельминтов очень часто наблюдается гермафродитизм. Прекрасным примером может служить бычий цепень, состоящий из множества члеников. В каждом из них есть собственные органы размножения — яичники и семенные железы. Когда половые продукты в них созревают, семенные тельца каждого отдельного членика могут оплодотворить яйца того же членика. Происходит то, что принято называть самооплодотворением. Однако известно, что у цепня яйцевые клетки одного членика могут оплодотворяться также семенными тельцами другого членика.
Некоторые ленточные черви, живя в кишечнике, время от времени отторгают от себя зрелые членики, которые способны самостоятельно двигаться. В лабораторных условиях можно наблюдать, как членик, ползая по чашке Петри или предметному стеклу, оставляет за собой полоску жидкости белого цвета, которая содержит тысячи яиц.
Таким образом, только очень высокой половой производительностью многих гельминтов, подкрепленной к тому же гермафродитизмом, уравновешивается непроизводительная потеря огромного количества яиц, которая могла бы быть роковой при отсутствии подобной приспособленности.
У свободноживущих гермафродитных животных отмечаются самые разнообразные заботы о том, чтобы, несмотря на нахождение женских и мужских половых органов в одной и той же особи, не произошло бы самооплодотворения, чтобы по возможности обе половые клетки, при слиянии которых образуется способное к развитию яйцо, принадлежали разным особям. Наиболее распространенное приспособление для предотвращения самооплодотворения заключается в том, что мужские и женские половые органы развиваются не одновременно, а один за другим, так что более молодые особи, у которых сперва созревают лишь мужские половые органы, функционируют и качестве самцов, и, лишь позднее, когда видоизменяются эти мужские железы, начинают созревать женские. Такое приспособление называется последовательным гермафродитизмом. Потом оно превращается и раздельное благодаря тому, что у одних особей остаются недоразвитыми мужские, у других - женские половые органы.
Чтобы не утратить способности зародышевого вещества к созданию новых отклонений, даже у самых типичных гермафродитных паразитов выработались замечательные приспособления. Так, при внимательном исследовании сосальщика спайника, живущего на жабрах рыб, обнаружили, что он состоит из двух цельных червей нормальной формы, которые срастаются между собой в виде буквы X. Из яиц червя выходят отдельные молодые особи, которые садятся на жабры хозяина и отличаются от родителей наличием особой брюшной присоски и шипика. Эти животные давно, известны и получили особое название— дипорп. Вскоре после рождения дипорпы отыскивают друг друга. Один из них схватывает своей брюшной присоской другого за спинной. шипик и оба срастаются в таком положении. У обоих, животных имеются совершенно развитые мужские и женские, половые органы. При срастании они располагаются таким образом, что семенники одной соединяются с яичниками другой и наоборот. Так происходит непрерывное взаимное оплодотворение.
Представляет интерес еще одна форма приспособления паразитов — явление живорождения, свойственное некоторым видам нематод и трематод.
На жабрах пресноводных рыб обитает гиродактил — живородящий сосальщик. У него нередко бывает виден внутри тела взрослого животного молодой зародыш, в котором размножается уже третье поколение, а в третьем имеется зачаток четвертого.
Среди нематод также встречаются живородящие виды: трихинелла, ришта и др. Благодаря живорождению жизненный цикл трихинелл сокращен. Одна особь млекопитающего играет для них роль окончательного, а затем и промежуточного хозяина. По расчетам ученых, в каждой самке трихинеллы имеется 20-25 личинок, обладающих способностью к внедрению в мышечные волокна.
В случае раздельности полов наиболее выраженному регрессивному превращению подвергаются самки. Самцы обыкновенно сохраняют большую подвижность и по своей организации меньше отличаются от ближайших свободноживущих родичей. Вероятно, именно самцу принадлежит активная роль в отыскании особи противоположного пола.
В отличие от цестод и трематод нематоды, как правило, раздельнополые. Акта совокупления у этих паразитов, не обладающих способностью более или менее долгое время жить в искусственных средах, где можно было бы изучать их, никто не наблюдал. Поэтому о нем можно судить лишь по аналогии со свободноживущими формами.
С увеличением количества откладываемых яиц повышается и их способность переносить неблагоприятные условия внешней среды. Они приобретают новые качества: покрываются плотными защитными оболочками, скорлупой и т. п. Так, яйца аскарид могут выдержать продолжительное пребывание в 30-40%-ном алкоголе, а яйца острицы даже развиваются в спирте.
В ряде случаев у паразитических червей наблюдается понижение плодовитости, которая может зависеть от восприимчивости хозяина к данному виду гельминта. Одна самка собачьей анкилостомы, паразитируя в организме собаки, в среднем откладывает 16 тыс. яиц в сутки, а в организме кошки — лишь около 2300. Аналогичные факты были установлены и относительно других гельминтов.
Английский ученый Мак Кой наблюдал уменьшение яйцеклеток у анкилостом, развившихся в собаках, подвергнутых вторичному заражению. У контрольных собак на одну самку анкилостом приходилось в среднем по 17190 яиц, в то время как у вторично зараженных — лишь по 6364 яйца.
По вопросу о влиянии беременности животных на плодовитость гельминтов имеются противоречивые данные. Временное прекращение продукции яиц, а иногда и полное освобождение животных от паразитов, как считают некоторые исследователи, зависит от химико-физического состояния кишечника хозяина в ближайшие дни после родов. Другие авторы, наоборот, наблюдали, что гельминты у беременных выделяли вдвое больше яиц по сравнению с теми, которые живут у небеременных животных.
На плодовитость гельминтов отрицательно влияет также голодание хозяина. Ученые установили, что у очень истощенных больных людей особи широкого лентеца были мелкими, тонкими и весили 4-6 г, что приводило их к низкой плодовитости. Также установили, что в нерестящихся голодающих лососях ленточные черви эуботриум теряют большую часть стробилы. У лососей до голодания живая масса цестод в среднем составляла 24,3 г, а у голодающих она уменьшилась до 2,8 г.
Интересные эксперименты осуществляли и на курах. Кур опытной группы кормили путем внутримышечных инъекции. Никакой другой пищи, кроме питательных раствором, содержащих витамины и животные белки, они не получали. При экспериментальном заражении аскаридами у кур данной группы развилось меньше паразитов и они были мельче, чем у кур, питавшихся нормально.
Американский исследователь С.Рейд на протяжении нескольких лет наблюдал зa состоянием цестод райллетина, паразитирующих в цыплятах. Он выявил, что при условии голодания цыплят псе запасы гликогена гельминты потребляют в течение 24-28 часов. Ленточные гельминты настолько при этом истощаются, что их мускулатура, лишенная источника энергии, не способна противостоять перистальтике кишечника, и паразиты, обрываясь у головного конца, выделяются наружу. При этом членики отделяются уже через 24 часа. При восстановлении питания членики снова отрастают от головного конца паразита и происходит нормальное продуцирование жизнеспособных яиц.
Чрезвычайное увеличение плодовитости паразитом, помимо факторов питания, обусловлено другим важным моментом — непрерывно действующим естественным отбором наиболее плодовитых особей. Однако плодовитость паразитом должна быть строго соразмерной с шансами встретиться с хозяином и заразить его, в противном случае при избыточной плодовитости ставится под угрозу существование хозяина, а также самого паразита, при недостаточной плодовитости не гарантируется встреча с хозяином и паразиту грозит вымирание. Эту соразмерность, выработавшуюся в процессе эволюции, надо признать серьезным прогрессивным признаком гельминтов.
Таковы основные условия размножения паразитических червей. Отдельные особи гельминтов, как правило, недолговечны и могут погибнуть вследствие воздействия неблагоприятных факторов. Однако, производя в течение жизни тысячи и миллионы новых особей, они тем самым сохраняют жизнь вида.
Животные, у которых значительное количество потомства уничтожается, откладывают и огромные массы яиц. Стоит вспомнить рыб, откладывающих миллионы икринок. Животные, яйца которых не развиваются внутри материнского организма и не достигают там зрелости (не получают достаточного количества питательных веществ, в частности желтка), обладают способностью рано вылупляться из яиц и самостоятельно добывать пищу. С этим связано образование временных личиночных органов и приобретение, формы, резко отличающейся от половозрелых животных. Из жизни непаразитических животных можно привести достаточно примеров того, что и на их развитие, как и на развитие паразитов, климатические условия иногда действуют сдерживающим образом и что форма яйца приспосабливается к предстоящему существованию. Упомянем хотя бы о бедных желтком и снабженных тонкой оболочкой летних яйцах и о толстостенных, богатых желтком зимних яйцах некоторых ресничных червей и коловраток.
Плодовитость нередко повышается за счет бесполого размножения, происходящего в определенный период жизненного цикла. Например, у трематод существует партогенетическое размножение (развитие яиц без предшествующего оплодотворения). Оно обеспечивает возможность произвести многочисленное потомство даже из одной единственной особи.
Читайте также: