
Особенности метаболизма у прокариот внутриклеточных паразитов
Хламидии –сферические грамотрицательные бактерии, облигатно паразитирующие в клетках теплокровных организмов. Они неспособны самостоятельно синтезировать высокоэнергетические соединения и обеспечивать собственные потребности в энергии. Нормальное развитие хламидий возможно только в условиях внутриклеточного паразитирования. Размножаются бинарным делением.
Риккетсии – мелкие грамотрицательные палочковидные бактерии, облигатно паразитирующие в различных клетках. Не растут на питательных средах, для культивирования используют куриные эмбрионы.
Микоплазмы – мелкие бактерии без истинной клеточной стенки, которую заменяет трехслойная клеточная мембрана, обеспечивающая осмотическую резистентность бактерий. Основной компонент клеточной мембраны – холестерин. Микоплазмы не способны к образованию холестерина и утилизируют его из тканей или питательных сред.
Дисбактериоз. Причины развития. Лабораторная диагностика. Эубиотики в лечении дисбактериоза.
Дисбиоз (дисбактериоз) — качественное и количественное изменение состава нормальной микрофлоры макроорганизма. Вследствие общего характера нарушений обменных процессов при дисбактероизе он играет определенную роль в развитии:
• нарушений свертываемости крови.
В то же время дисбактериоз может быть ярко выражен клинически в виде нарушений деятельности дыхательной системы (бронхиты и бронхиолиты, хронические заболевания легких) и желудочно-кишечного тракта (диарея, неспецифический колит, синдром малой сорбции), хотя может протекать и без выраженных клинических проявлений.
Диагноз дисбактериоза устанавливается повторным (с интервалом в 5—7 дней) бактериологическим исследованием материала, взятого из того или иного биотопа.
При этом количественная оценка результатов определения видов и вариантов обнаруживаемых микроорганизмов, входящих в состав обследуемого биоценоза, является обязательной.
Наличие дисбиоза определяется изменениями состава нормальной микрофлоры, а его выраженность — степенью этих изменении.
Показатели дисбактериоза:
• снижение общего количества бактерий, представителей нормальной микрофлоры или их отдельных представителей;
• увеличение числа редко встречающихся в норме микроорганизмов или появление не свойственных данному биотопу видов;
• появление измененных вариантов микроорганизмов — представителей нормальной микрофлоры (изменение биохимических свойств штаммов этих микроорганизмов и/или приобретение ими некоторых факторов вирулентности);
• ослабление антагонистической активности микроорганизмов входящих в состав нормальной микрофлоры.
Стадии дисбактериоза
I стадия дисбактериоза характеризуется умеренным уменьшением численности облигатных бактерий в полости кишечника. Патогенная микрофлора, как правило, развита незначительно, а признаки расстройства функции кишечника (симптомы болезни) отсутствуют.
II стадия дисбактериоза характеризуется критическим снижением численности бифидобактерий и лактобацилл кишечника (облигатная флора). При этом отмечается стремительное развитие популяции патогенных бактерий. На этой стадии дисбактериоза появляются первые признаки нарушения работы кишечника: понос (зеленоватого цвета), боли в животе, метеоризм.
III стадия дисбактериоза характеризуется воспалительным поражением стенок кишечника под влиянием патогенов. На этом этапе дисбактериоза понос приобретает стойкий хронический характер, а в каловых массах определяются частички непереваренной пищи. У детей может появиться отставание в развитии.
IV стадия дисбактериоза представляет собой стадию, предшествующую острой кишечной инфекции. На этом этапе дисбактериоза облигатная флора кишечника присутствует в очень малых количествах. Основное количество микробов приходится на условно патогенных и патогенных бактерий и грибов. На этой стадии дисбактериоза возникает общее истощение организма, анемия, авитаминоз.
Наиболее логичной коррекиией состава микрофлоры при дисбак-териозе выглядит заместительная терапия живыми бактериями, населяющими толстый кишечник. Она проводится с помощью эубиотиков — препаратов, содержащих леофилизированные живые штаммы микроорганизмов — представителей нормальной микрофлоры.
К наиболее известным в настоящее время такого рода препаратам относятся:
• бификол (комбинированный препарат из 2 предыдущих);
• бифи-форм (комбинированный препарат из бифидобактерий и энтерококков — Enterococcus faecalis);
Однако применение эубиотиков для лечения больных с дисбактериозом не всегда достигает клинического успеха. Установлено, что входящие в эти препараты микроорганизмы в организме человека стойко, как правило, не приживаются. После прекращения поддерживающей терапии искусственно введенные штаммы быстро элиминируются из кишечника и замещаются случайной микрофлорой.
Выбор эубиотика по результатам бактериологического анализа (например, назначение бифидумбактерина при дефиците бифидобактерий) — не более чем иллюзия: клинический опыт показывает отсутствие коррелятивной зависимости клинической эффективности назначенного препарата и показателей дисбактериоза.
В связи с этим в последние годы в значительной степени утвердилось мнение, что применение антибиотиков (что раньше категорически опровергалось), с учетом чувствительности к ним условно-патогенной микрофлоры, одновременно с применением антибиотикорезистентных вариантов бифидобактерий и лактобактерий может привести к нормализации микробиоценоза.
3. Многолетние клинические исследования по лечебному и профилактическому применению препаратов на основе представителей нормальной микрофлоры показали, что наименьшим побочным эффектом при длительном применении обладают эубиотики. в состав которых входят бифидобактерий.Это послужило основанием создавать препараты и продукты питания с включением в них бифидобактерий.
Установлено, что положительный эффект на организм человека оказывают продукты питания, содержащие живые бактерии в количестве не менее 10 8 КОЕ/мл.
Кисломолочный бифидумбактерин (бифидобактерии, добавленные в кисломолочные продукты) и йогурты, содержащие эти микроорганизмы, прошли широкую клиническую апробацию.
Спектр показаний к их применению достаточно широк:
• терапия запоров и их профилактика.
Для обозначения препаратов живых культур бактерий — представителей нормальной микрофлоры человека (чаще бифидобактерии, лактобактерий), добавляемых в продукты питания (чаще кисломолочные) в целях профилактики дисбактериозов и обеспечения функционального питания, в последнее время в литературе вместо термина "эубиотики" все чаще используют термин "пробиотики", распространяя его на все остальные аналогичные препараты.
Спирохет, актиномицетов, риккетсий, хламидий, микоплазм
Сравнительная характеристика
Методы изучения морфологии микоплазм.
1. В световом микроскопе в мазках, окрашенных по Романовскому-Гимза, обнаруживаются лишь самые крупные формы микоплазм.
2. ФКМ и ТПМ нативных препаратов для выявления нитевидных форм.
3. Электронная микроскопия для выявления ультраструктурных компонентов микоплазм.
В табл. 18 приведена сравнительная характеристика описанных выше микроорганизмов.
| Признак | Спирохеты | Актиномицеты | Риккетсии | Хламидии | Микоплазмы | |
| Тип строения КС | Г рам- | Грам+ | Грам- | Грам- | Не имеют КС, подобны Грам- | |
| Методы изучения морфологии | Окраска по Романовскому-Гимзе, серебрение по Морозову, ТПМ или ФКМ | Простые методы, окраска по Граму | По методу Здродовского, по Граму, ЭМ | Окраска мазков-соскобов по Романовскому-Гимза | ФКМ, ЭМ, окраска по Романовскому-Гимза для выявления нитевидных форм | |
| Морфология | Тонкие спирально извитые нити, изогнутые вокруг центральной оси, до 50 мкм. Трепонемы (8-20 завитков), боррелии (3-10завитков), лептоспиры (12-30 завитков) | Нитевидные ветвистые клетки, имеющие вид палочки | Мелкие полиморфные бактерии кокковидной, палочковидной, реже — нитевидной формы | Элементарные тельца сферической формы (вне организма человека) и ретикулярные тельца (внутри-клеточные) | Мелкие или крупные сферические, овоидные или нитевидные клетки | |
| Особенности ультрастуктуры | экзо-споры | - | + как способ размножения | - | - | - |
| ж | Осевые фибриллы (периплазмати-ческие жгутики) | - | - | - | +/- | |
| к | - | - | Микрокапсулу образуют в клетках хозяина | - | Капсуло-подобный слой | |
| кс | Типичной нет (слабо выражен пептидогликановый слой) | В составе есть сахара, отсутствующие у истинных бактерий, миколовые и жирные кислоты | + | + | Отсутствует | |
| Представители | Сапрофитные и патогенные: трепонемы, боррелии, лептоспиры | Большинство —сапрофиты, патогенные: роды актиномицеты и нокардии | Облигатные внутриклеточ- ные паразиты | Облигатные внутриклеточные паразиты | Патогенные и сапрофитные | |
| Роль в патологии | Сифилис и бытовые трепонематозы, возвратные тифы (эпидемический и эндемический), болезнь Лайма, лептоспироз | Актиномикоз (шейно-челюстно-лицевой, органов дыхания, ЖКТ) | Сыпной тиф и болезнь Бриля-Цинссера, эндемический клещевой риккетсиоз | Трахома, паховый лимфо-гранулематоз, урогенитальные хламидиозы, синдром Рейтера, пневмонии, орнитоз, атеросклероз | Респира-торные и урогениталь-ные инфекции |
Для роста и размножения микроорганизмы нуждаются в веществах, используемых для построения структурных компонентов клетки и получения энергии. В основе жизнедеятельности микроорганизмов лежит непрерывный обмен веществ и энергии с окружающей средой (метаболизм).
Метаболизм(от лат. metabole — изменение, превращение) — совокупность всех химических превращений в клетке, имеет две составляющих (рис. 56):
| – анаболизм (от греч. anabole — подъем) (конструктивный (пластический) метаболизм, ассимиляция) — биосинтез макромолекул из простых низкомолекулярных соединений и их ассимиляция (усвоение, накопление); – катаболизм (энергетический метаболизм, диссимиляция) — химический ферментативный процесс разрушения высокомолекулярных веществ до мономеров, направленный на получение энергии и ее запасы в форме АТФ. |
|

Метаболизм у прокариотов принципиально протекает так же как у высших животных и растений.
Особенности метаболизма у прокариотов:
– бактерии разнообразны по своим пищевым потребностям. Одно и то же соединение для одних бактерий может быть продуктом питания, а для другого — ядовитым веществом. Известны бактерии, способные усваивать фенол, парафин, уксусную кислоту, антибиотики;
– способность включать в обмен веществ любые органические и неорганические соединения;
– высокая интенсивность метаболизма;
– высокая адаптационная способность к меняющимся условиям окружающей среды;
– многообразие путей метаболизма, промежуточных и конечных продуктов;
– возможность искусственного культивирования многих микроорганизмов в бесклеточных средах;
– несовершенство регуляции метаболических процессов.
Риккетсии — облигатные внутриклеточные паразиты (не способны синтезировать некоторые макромолекулы).
Хламидии— облигатные внутриклеточные паразиты (не способны синтезировать некоторые макромолекулы) и энергетические паразиты (не способны синтезировать АТФ).
Микоплазмы—мембранные паразиты (не способны синтезировать стерины для ЦПМ).
Изучение процессов метаболизма у микроорганизмов необходимо для понимания механизмов патогенеза заболеваний, для идентификации микроорганизмов и этиологической диагностики заболеваний, для проведения химиотерапии, а также для получения необходимых для человека материалов с использованием биотехнологии.
Нам важно ваше мнение! Был ли полезен опубликованный материал? Да | Нет
Риккетсии – прокариотические микроорганизмы, неподвижные, клеточная стенка построена аналогично Гр- бактериям. Морфологически риккетсии это полиморфные организмы – чаще наблюдаются небольшие палочковидные формы, но могут быть длинные нитевидные образования и очень мелкие, проходящие через бактериальные фильтры. Риккетсии – облигатные паразиты. Существование вне клеток организма хозяина у этих бактерий невозможно, тк не могут синтезировать HAД,а следовательно и получать энергию. R.prowazekii-возб-ль эпидемич сыпного тифа(через вшей) и болезни Брилла и R.typhi-возб-ль эндемического сыпн тифа. Другие виды выз-т пятнистую лихорадку.
Хламидии – облигатные паразитические неподвижные бескапсульные Гр- бактерии. Полная зависимость от клетки хозяина обусловлена неспособностью хламидий синтезировать АТФ. Вне клеток хозяина хламидии существуют в виде элементарных телец сферической формы. Эндоцитозом проникают внутрь клетки. В клетке хозяина они подавляют слияние с лизосомами ,превращаются в ретикулярные тельца, которые начинают делиться, в результате чего образуются включения внутри цитоплазмы – микроколонии, содержащие промежуточные формы развития. Потом экзоцитозом выходят из клетки,которая под-ся лизису.Покидая клетку, они превращаются в элементарные тельца. Цикл развития продолжается 40-72 ч.
Они не имеют ПГ в клет стенке,выраженный плеоморфизм(кокки,различ потинкториал свойствами и размерам)
Микоплазмы – Гр-, небольшого размера, неподвижные, не образующие спор. обладающие выраженным полиморфизмом: могут иметь вид овоидных, круглых или удлиненных, а также нитевидных образований. Самые мелкие проходят через бактериальные фильтры. Такое морфологическое разнообразие объясняется отсутствием клеточной стенки, вместо которой микоплазмы окружены трехслойной мембраной. Микоплазмы – хемоорганотрофы (в качестве доноров электронов используют только органические соединения) и не способны к синтезу стеринов, необходимых для их роста. Ауксотрофны по ХС. По типу дыхания – факультативны анаэробы. Попадая во внешнюю среду, быстро погибают. Обладают уникальным свойством мембранных паразитов. Механизм включает тесный контакт мембран клеток бактерии и клеток хозяина. Липидные компоненты микоплазм диффундируют в мембрану клеток, а холестерин клетки макроорганизма поступает в мембрану микоплазм. Заканчивается этот процесс откреплением микоплазм от клеток, либо поглощением их вследствие фагоцитоза (внутриклеточное размнжение).
2. Механизмы, определяющие антивирусную активность Т-эффекторов (CD4+, CD8+):
1. Цитолиз вирусинфицированных клеток. 2. Апоптоз зараженных клеток. 3. Продукция гамма-интерферона. 4. Активация макрофагов.
Вопрос 2
ИММУНОЛОГИЯ
Механизм противовирусного иммунитета
Все строится на принципе, какая мишень, это либо зараженная клетка, т е инфицированная, либо это свободный вирион, который только попал в организм, но еще не инфицировал клетку, либо синтезировался и вышел из клетки.
Свободные вирионы атакуются антителами (это гуморальный специфический иммунитет), протективность которых определяется блокадой вирусных рецепторов. Это действие хорошо выражено при инфекциях с длительным инкубационным периодом, когда вирус перед тем, как достичь мишеней, должен пройти через этап вирусемии (этап циркуляции), где он нейтрализуется даже небольшим количеством антител.
А против внутриклеточных форм, уже включается специфический клеточный, клеточный и гуморальный неспецифический иммунитет. Специфический клеточный – цитотоксические т-лимфоциты. Что они делают: уничтожение зараженной клетки, элиминация внутриклет вируса и использование цитотоксического и антивирусного потенциала других клеток эффекторов. Для уничтожения клетки они могут использовать цитолиз, разрушая мембрану, но чаще – апоптоз вызывают. Элиминация вируса без повреждения клетки – т-лимф могут секретировать гамма интерферон, он тормозит вирусную репликацию. Но это включается очень поздно, обычно тогда, когда клетки уже несут на себе вирусные антигены, которые воспринимают т клетки, помимо этого гамма интерферон активирует макрофаги и НК.
Неспецифический гуморальный - Механизм действия интерферонов (альфа и бета – они секретируются всеми клетками и быстро, а гамма только активированными т-лимф) – (сами они цитокины с противовирусной активностью) при заражении вирусом клетка начинает синтезировать и выделять интерфероны, которые связываются с рецепторами других клеток соседних. В итоге интерфероны побуждают клетки синтезировать ферменты, которые блокируют синтез вирусных белков и разрушают вирусные мРНК и подавляют ее трансляцию на рибосомы клетки. В результате другие клетки остаются неинфицированные, данная реакция развивается в теч нескольких часов и держится 1-2 дня. Создается барьер из клеток, которые устойчивы к вирусы и сдерживающие его распространение. +доп. Отличие альфа бета гамма – а и б подавляют вирус внутри клетки и стимулируют экспрессию HLA-1 и повышают активность цитотокс т лимф; а гамма повышает
HLA-2 зависимую экспрессию АГ на поверхности зараженной клетки и усиливает цитотоксическую функцию макрофагов и натуральных киллеров.
Неспецифический клеточный – естественные киллеры (НК)они распознают неэпитопные структуры гликопротеинов, которые экспрессируются на мембране вирус инфицированных клеток, т е у них нет антигензависимой избирательности АТ и Т-лимф. При контакте с мишенью они выделяют содержимое своих гранул. Они вызывают апоптоз клетки или лизируют ее. Интерфероны усиливают цитотоксичность НК, а они в свою очередь продуцируют гамма интерферон как цитотокс т-лимф, способствуя интеграции механизмов противовирусной защиты
различают специфический и неспецифический, которые в свою очередь классифицируются:
Неспециф (врожденный) – клеточный (естественные киллеры NK) и гуморальный (интерфероны)
Специф (приобретенный) – клеточный (цитотоксические Т-лимфоциты) и гуморальный (антитела)
Неспециф клеточный и неспециф гуморальный + специф клеточный – эффективны против внутриклеточных форм (мишень – вирус инфицирующий клетку), а специф гуморальный – эффективны против внеклеточных форм (вирионы). Поэтому есть 2 мишени – вирион и вирус инфицир. клетку.
Естественные киллеры – они экспрессируют на своей поверхности вирусн пептиды в составе hla-1 hla – 2. Прямой эффект – цитолиз и апоптоз, антителзависимая клеточно опосредованная цитотоксичность.
Интерфероны – контролируют вирусную репликацию и ингибируют синтез вирусных белков внутри клетки, +стимулирует соседние клетки к синтезу антивирусных элементов)
Цитотокс. Т лимф – ццитолизм, апоптоз вирус инфицированных клеток. Cd-4 т-лимф через продукцию цитокинов (гамма интерферон) для активации cd-8, натуральных киллеров, макрофагов. (поцелуй смерти).
(+интерфероны – это цитокины с противовирусной активностью:
-гуморальные факторы неспецифического иммунитета
-выделяют 3 типа – альфа бета гамма; различают по происхождению,физико-химическим свойствам и биоактивности)
2. Механизмы, определяющие антивирусную активность Т-эффекторов (CD4+, CD8+):
+1. Цитолиз вирусинфицированных клеток.
+2. Апоптоз зараженных клеток.
+3. Продукция гамма-интерферона.
+4. Активация макрофагов.
5.Антителозависимая клеточная цитотоксичность.
-1 просто чаще всего в качестве мишени для т-киллеров выступают вирус-зараженные клетки (но это может быть любая клетка, представляющая АГ в комплексе с HLA-1)
-2 создается контакт с клеткой мишенью, и т-киллер (сд8) выделяет перфорины и создает каналы, а гранзимы которые идут по каналам запускают апоптоз
-3 сд-4 клетки продуцируют цитокины, когда распознают клетку чужеродную с помощью TCR. А как раз этими цитокинами являются интрферон гамма и IL-2, которые в свою очередь стимулируют другие эффекторы, обладающих собственной биоцидностью (макрофига, натур киллеры).
-4 сд-4 клетки продуцируют цитокины, когда распознают клетку чужеродную с помощью TCR. А как раз этими цитокинами являются интрферон гамма и IL-2, которые в свою очередь стимулируют другие эффекторы, обладающих собственной биоцидностью (макрофига, натур киллеры).
-5 они обладают опосредованной (непрямой) цитотоксичностью, распознают клетку чужеродную с помощью TCR.
БАКТЕРИОЛОГИЯ
Дата добавления: 2018-02-28 ; просмотров: 398 ;
7.1. Особенности метаболизма микроорганизмов
Микроорганизмы обладают рядом принципиальных отличий в протекании метаболических процессов.
А. Бактерии, вследствие принципиально иной клеточной организации, по сравнению с эукариотическими клетками, обладают и принципиально иными чертами осуществления метаболических процессов, которые, в свою очередь, позволяют им занимать их уникальное положение в мире живых существ.
2. Прокариотическая клетка, по сравнению с эукариотической обладает несравненно более высокой скоростью метаболических процессов.
3. Бактерии в целом обладают высокой адаптационной способностью к меняющимся условиям окружающей среды. Эта способность реализуется у них как на клеточном, так и на популяционном уровнях.
а. На клеточном уровне эта способность детерминируется у прокариот высокой скоростью изменчивости, как фенотипической, так и генотипической.
Б. Вирусы, в отличие от клеточных форм жизни не обладают, строго говоря, собственным метаболизмом, используя метаболические потенции клетки-хозяина.
7.2. Классификация бактерий по источнику углерода
Так как жизнь на Земле основана на соединениях углерода, этот элемент является основным для построения и успешного функционирования бактериальной клетки.
А. Бактерии, использующие в качестве источника углерода неорганические соединения (углекислый газ, карбонаты), называются аутотрофами. Аутотрофы не вызывают патологических процессов в организме человека и не являются предметом изучения медицинской микробиологии.
Б. Медицинская микробиология имеет дело с бактериями, которые в качестве источника углерода используют органические соединения. Такие бактерии называются гетеротрофами.
2. Те гетеротрофы, которые используют в качестве источника углерода органические соединения (метаболиты) живой клетки, называются паразитами.
7.3. Пути проникновения питательных веществ в бактериальную клетку
А. Процесс проникновения питательных веществ через оболочку бактериальной клетки без затраты энергии называется диффузией. Энергия для осуществления диффузии не нужна, так концентрация диффундирующих веществ вне клетки выше, чем внутри ее, т.е. их перемещение происходит по градиенту концентрации.
1. Если в этом процессе не участвуют пермеазы, то это – простая диффузия. Таки путем внутрь бактериальной клетки проникают вещества, образующие истинные растворы.
2. Ряд субстратов (например, глицерин) диффундируют через оболочку бактериальной клетки с помощью пермеаз. Такой процесс называется облегченной диффузией.
Б. Концентрация подавляющего большинства необходимых питательных веществ внутри бактериальной клетки превышает их концентрацию в околоклеточном пространстве. Поэтому для их перемещения через клеточную оболочку необходима затрата энергии. И, конечно, обязательно в этом процессе участвуют пермеазы.
1. Если при этом переносимые молекулы не претерпевают химических изменений, то такой процесс называется активным транспортом.
2. Если транспорт молекулы сквозь клеточную оболочку сопровождается ее химической модификацией (например, фосфорилированием), то такой процесс называется транслокацией химических групп.
7.4. Классификация бактерий по потребностям в факторах роста
Под факторами роста (или ростовыми факторами) понимают макромолекулы (азотистые основания, аминокислоты, витамины, липиды и пр.), необходимые для построения и функционирования бактериальной клетки.
А. Бактерии, способные самостоятельно синтезировать все необходимые им факторы роста из более простых веществ (в типичном случае – солей аммония и глюкозы), называются прототрофами.
7.5. Классификация бактерий по особенностям энергетического метаболизма
А. Бактерии, которые получают энергию непосредственно солнечного света, называются фототрофами. Такие бактерии не вызывают патологические процессы в организме человека и медицинской микробиологией не изучаются.
Б. Бактерии, которые получают энергию, разлагая более сложные химические вещества до более простых в процессе окислительно-восстановительных реакций с запасанием полученной энергии в виде АТФ и других макроэргических соединений, называются хемотрофами.
1. Донорами электронов при этом могут выступать как неорганические, так и органические соединения.
а. Хемотрофы, использующие в качестве доноров электронов в химических реакциях, сопровождающихся синтезом АТФ, неорганические соединения, называются литотрофами.
б. Хемотрофы, использующие в качестве доноров электронов в химических реакциях, сопровождающихся синтезом АТФ, органические соединения, называются органотрофами.
2. Акцептором электронов в таких реакциях может служить молекула, которая находится или вне бактериальной клетки (так называемый внешний акцептор электронов) или внутри бактериальной клетки (в этом случае говорят о внутреннем акцепторе электронов).
1. Если в качестве такого внешнего акцептора электронов выступает молекулярный кислород, то такой процесс называется аэробным дыханием.
2. В качестве внешнего акцептора электронов некоторые бактерии (например, клостридии) используют не молекулярный кислород, а нитрат или фумарат. В этом случае используют термин анаэробное дыхание (нитратное или фумаратное дыхание).
б. Цепь окислительно-восстановительных реакций для получения макроэргических соединений с переносом электронов на внутренний акцептор называется ферментацией или брожением.
7.6. Классификация бактерий по отношению к кислороду воздуха
Вовлечение кислорода воздуха в окислительно-восстановительные реакции сопровождается образованием перекисного радикала (О • ) – атома кислорода с лишним электроном на внешней оболочке. Это очень сильный окислитель. В зависимости от того, как бактерии решают проблему его нейтрализации, и зависит их способность существовать в кислородной среде. На практике простейшим методом определить отношение изучаемого вида бактерий к кислороду воздуха, является регистрация характера роста этих бактерий при засеве уколом в столбик полужидкой среды (Рис. 7-1).
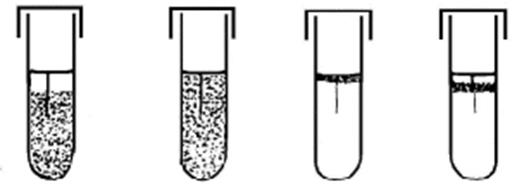 Рис. 7-1. Рост (слева направо) облигатных анаэробов, факультативных анаэробов, облигатных аэробов и микроаэрофилов при засеве уколом в столбик полужидкого агара |
А. Бактерии, которым кислород абсолютно необходим как внешний акцептор электронов, образовавшийся перекисный радикал с помощью фермента супероксиддисмутазы переводят в перекись кислорода. Это соединение также является окислителем, хотя и менее сильным, чем перекисный радикал. Перекись водорода нейтрализуется с образованием воды и молекулярного кислорода с помощью, например, каталазы. В отсутствии кислорода такие бактерии не растут, так как не могут использовать другие молекулы в качестве акцептора электронов, и поэтому называются облигатными аэробами.
Б. Размножение некоторых бактерий возможно только в атмосфере особого газового состава. Такие бактерии обладают супероксиддисмутазой, но степень активности каталазы или других ферментов, нейтрализующих перекись водорода, может быть недостаточной.
1. Микроаэрофилы растут при более низком, чем в атмосферном воздухе, содержании кислорода. К ним относятся, например, молочнокислые бактерии, в большом количестве населяющие кишечник человека.
2. Капнофилы нуждаются в более высоком, чем в атмосферном воздухе, содержании углекислого газа. К ним, например, относятся бруцеллы, вызывающие бруцеллез.
3. Существуют также бактерии, которые требуют для своего успешного культивирования создания особой газовой смеси, включающей как пониженное количество кислорода, так и повышенное – углекислого газа. Т.е. такие бактерии можно определить одновременно как микроаэрофилы и капнофилы. Такие особые требования к аэрации при культивировании предъявляют, например, кампилобактеры и геликобактер.
В. Подавляющие большинство бактерий могут расти в отсутствии кислорода, вследствие чего их называют анаэробами.
1. Некоторые бактерии лишены не только ферментов, нейтрализующих перекись водорода, но и супероксиддисмутазы. В результате чего они погибают при малейшем контакте с атмосферным воздухом и называются облигатными анаэробами. При микробиологической диагностике болезней, вызываемых этой группой бактерий, и взятие патологического материала и посев его на питательные среды, не говоря уже о самом культивировании, необходимо проводить с соблюдением специальных приемов, исключающих контакт возбудителя с атмосферным воздухом.
2. Часть анаэробов, не имея ферментов, нейтрализующих перекись водорода, содержат супероксиддисмутазу. Такие бактерии – аэротолерантные анаэробы – выдерживают кратковременный контакт с атмосферным воздухом, например во время взятия патологического материала или засева его на питательную среду, но культивироваться могут лишь в анаэробных условиях.
3. Однако наиболее часто медицинская микробиология имеет дело с бактериями, которые имеют полный набор ферментов, позволяющих им расти и размножаться и в присутствии и в отсутствии кислорода воздуха – т.е. в любых условиях аэрации. Такие бактерии называются факультативными анаэробами.
7.7. Особенности метаболизма риккетсий, хламидий, микоплазм
Риккетсии, хламидии, а также микоплазмы имеют, по сравнению с остальными прокариотами, принципиальные отличия метаболических процессов.
А. Риккетсии не способны синтезировать некоторые макромолекулы, вследствие чего вынуждены получать их в готовом виде от клетки-хозяина. Поэтому они могут функционировать только внутри живой эукариотической клетки (облигатный внутриклеточный паразитизм).
7.8. Классификация бактериальных ферментов
А. По своей локализации бактериальные ферменты подразделяются на две группы.
2. Ферменты, которые локализуются внутри бактериальной клетки, называются эндоферментами. Эндоферменты содержатся в периплазматическом пространстве грамотрицательных бактерий (здесь находится часть тех ферментов, которые у грамположительных бактерий являются экзоферментами, например, протеазы), а также в цитоплазматической мембране и цитоплазме.
Б. По зависимости своего синтеза от наличия субстрата, на который действует фермент, бактериальные ферменты подразделяются также на две группы.
1. Конститутивные ферменты синтезируются постоянно, вне зависимости от наличия в данный момент субстрата для их действия.
В. К ферментам вирулентности относятся такие бактериальные ферменты, чьим субстратом действия являются клетки и ткани макроорганизма, в котором паразитирует микроб. Более подробно эта группа бактериальных ферментов будет охарактеризована в 19 главе.
Механическое удерживание земляных масс: Механическое удерживание земляных масс на склоне обеспечивают контрфорсными сооружениями различных конструкций.


Общие условия выбора системы дренажа: Система дренажа выбирается в зависимости от характера защищаемого.

Папиллярные узоры пальцев рук - маркер спортивных способностей: дерматоглифические признаки формируются на 3-5 месяце беременности, не изменяются в течение жизни.
Читайте также: