
Паразиты одноклеточные или многоклеточные
Паразиты – организмы, живущие за счет своих носителей (растения, животные, люди). Данные паразитирующие организмы ни коим образом не связаны со своими хозяевами на генетическом уровне, тем не менее довольно серьезно осложняют им жизнь. То есть, биология паразитов подразумевает то, что их деятельность осуществляется за счет других организмов.

Зачастую, попав в тело человека такие организмы не дают о себе знать довольно длительное время, таким образом они себя защищают от воздействия медикаментозных препаратов, используемых при проведении антипаразитарной терапии. Это позволяет им на протяжении длительного времени развиваться и размножаться.

Группы паразитов
Специалистами выделяется несколько групп паразитов которые разделяются между собой по месту локализации.
- Эктопаразиты – организмы, ведущие паразитарный образ жизни на теле человек (в основном кровососущие насекомые). Эктопаразиты отличаются достаточно сложным жизненным циклом, подразумевающим, что для прохождения всех его этапов, паразиту требуется смена носителей.
- Эктопаразиты – опасные переносчики разнообразных болезней и инфекции, по этой причине борьба с ними играет важную роль в профилактике и устранении вызываемых недугов.
- Эндопаразиты – организмы развивающиеся и размножающиеся внутри тела носителя (черви, чесоточные клещи и прочие). Основной этап жизненного цикла эндопаразитов проходит в теле окончательного носителя.
Рассматривая паразитов в биологическом смысле, хотелось бы обратить внимание на то, что серьезный вред жизни человека могут нанести всего несколько десятков паразитирующих организмов. Поэтому медиками ставится важная задача: своевременная диагностика паразитарной инфекции и последующая эффективная терапия.

Простейшие паразиты
К простейшим паразитам относят одноклеточные организмы:
споровики;
инфузории;
жгутиковые;
амебы.
Чаще всего в человеческом организме встречаются представители жгутиковых – лямблии, основной причиной заражения паразитами является пренебрежение элементарными правилами гигиены.

Многоклеточные
К многоклеточным паразитирующим организмам относят червей, насекомых и паукообразных. Опасность заражения организма гельминтам в том, что черви механическим путем повреждают внутренние органы и ткани, а также отравляют организм человека изнутри продуктами своей жизнедеятельности – токсинами. Медицина разделяет гельминтов на несколько основных групп.
- Трематоды. Черви, отличающиеся сложным жизненным циклом (несколько этапов, подразумевающих переход от промежуточного носителя к основному).
- Ленточные черви – данные гельминты входят в группу облигатных паразитов, основное место локализации глистов: тонкий кишечник человека. Тело паразитов состоит из большого количества члеников, которые отделяются и вместе с яйцами выходят во внешнюю среду. К особенностям строения данных паразитов можно отнести то, что ленточные черви не имеют пищеварительной системы, то есть поглощение питательных веществ из тела носителя осуществляется всем телом гельминта. Наиболее известные представители этого вида глистов: широкий лентец, свиной /бычий цепень.
- Круглые черви – наиболее распространенный вид паразитов поражающий человеческое тело (острицы, аскариды, угрицы, трихинеллы, токсокары и т.д.). Попадают в организм носителя в зависимости от среды обитания: грязные руки, несоблюдение технологий при приготовлении мясных / рыбных продуктов, контакт с зараженными паразитарной инфекцией животными и людьми. Большинство представителей данного класса гельминтов имеют промежуточного носителя.
А.В.Чесунов, Д.М.Милютин, А.В.Евсеев
Нематоды, или собственно круглые черви, (Nematoda) удивительно разнообразны. Одни виды паразитируют только на взрослой стадии, другие — исключительно на личиночной, третьи — на всех стадиях; у одних нематод жизненный цикл протекает со сменой хозяев, у других — без; одни стремятся убить хозяина, другие, наоборот, стараются сохранить его живым как можно дольше; одни живут на поверхности тела, вторые — в кишечнике, третьи поражают внутренние органы.
Удивительна и непритязательность нематод в выборе хозяина. Конечно, большинство видов предпочитает жить в организмах позвоночных животных, но немало нематод облюбовало наземных членистоногих (которые могут стать как промежуточными, так и окончательными хозяевами), наземных малощетинковых червей (олигохет) и легочных улиток. Морские паразитические нематоды поражают и ракообразных, и иглокожих, и полихет, а также приапулид, сипункулид и даже своих собственных сородичей. Но все же полной неожиданностью оказалось открытие своеобразных видов нематод в одноклеточных простейших животных — фораминиферах.
Фораминиферы (Foraminiferida) относятся вместе с голыми, лишенными внутреннего скелета и наружной раковины амебами (Lobosea), радиоляриями (Radiolaria) и солнечниками (Heliozoa) к большой группе простейших — подтипу саркодовых (Sarcodia). Их общая особенность — передвижение с помощью временных выростов цитоплазмы — псевдоподий, или ложноножек. Фораминиферы довольно крупные существа для простейших: нередко их размер достигает одного миллиметра или даже более.
Почти все фораминиферы облачены в раковинку, которая может быть чисто органической, кожистой, а чаще либо построенной из подручного материала — песчинок и других мелких частиц, склеенных известковым цементом (агглютинированная), либо чисто известковой (секреционной). В домике есть отверстие, откуда вытягиваются длинные нитевидные, ветвящиеся псевдоподии. У многих видов раковинка пронизана тонкими порами, откуда также растут ложноножки. Из-за тяжелой раковинки фораминиферы если и ползают, то очень медленно, а чаще — неподвижны и даже могут быть прикреплены к субстрату. Раскинутые во все стороны псевдоподии образуют ловчую сеть, в которую попадают бактерии, одноклеточные водоросли и другие живые организмы, даже мелкие многоклеточные животные.
Изображения агглютинированных раковинок разных видов фораминифер (слева) и живой особи Allogromia ovoidea.
Для геологов не меньше, чем для биологов, фораминиферы — популярный объект исследования: их домики прекрасно сохраняются в ископаемом состоянии и с их помощью удобно датировать слои осадочных пород, в том числе нефте- и газоносных. Однако и в современных морях на дне существует плотное и разнообразное население фораминифер. Вот в таких простейших и обнаружены нематоды.
Это произошло совершенно неожиданно летом 1995 г. в Смитсоновском музее естественной истории в Вашингтоне (США). Просматривая подобранную серию морских паразитических нематод, один из авторов этих строк наткнулся на готовый глицериновый препарат со странной, похожей на плоское веретено с двумя устьями на вытянутых противоположных концах, фораминиферой, внутри которой находились две хорошо прокрашенные нематоды. Препарат аккуратно сделан безвестным лаборантом, разбиравшим пробы мейобентоса [1] из глубоководья у побережья Анголы.
Фораминифера, которую впоследствии определила сотрудница кафедры зоологии беспозвоночных МГУ им. М.В.Ломоносова Е.М.Майер, оказалась довольно редкой формой — Vanhoeffenella aff. gaussi. Боковые стенки ее агглютинированной раковинки затянуты кожистой прозрачной мембраной (характерный признак рода) — через эти своеобразные окна и были видны две нематоды (самец и самка). Они находились не просто в цитоплазме, а были заключены в большую шаровидную вакуоль, или камеру, с довольно плотными, хотя и мягкими, совершенно прозрачными кожистыми стенками.
Нематоды оказались не известны науке. Вместе с американским нематодологом Д.Хоупом мы исследовали их строение, описали новый вид и отнесли его к новому роду — Smithsoninema inaequale [2]. Родовое название нематода получила в честь Смитсоновского института, видовое — отражает большое различие между самцом и самкой.
Фораминифера Vanhoeffenella aff. gaussi. В ее внутренней кожистой камере поселилась семейная пара Smithsoninema inaequale — тонкий самец и толстая самка. Внизу они показаны “крупным планом”
Нитевидно тонкий (в диаметре до 20 мкм) самец свернут в пузыре несколько раз, так как его длина (почти 3 мм) многократно превышает размеры раковинки фораминиферы. У самца присутствуют все признаки типичных свободноживущих нематод: головные щетинки, большие амфиды (органы химического чувства), нормальный кишечный тракт, хвостовые железы. По форме амфида (в виде запятой), наличию твердого палочковидного образования (стилета) в ротовой полости, железистой глотке и особой форме копулятивных органов (спикул) самец этого вида несомненно принадлежит к семейству Camacolaimidae (группа морских свободноживущих нематод). Самка совсем другая — короткая (всего 0.8 мм длиной) и толстая (почти 100 мкм в середине тела). Амфидов у нее не видно, хотя головные щетинки есть. Стилета в ротовой полости нет, пищевод железистый, средняя кишка без видимого внутреннего просвета, хвостовые железы отсутствуют. Различия между самцом и самкой настолько велики, что, будь они встречены по отдельности, их вряд ли отнесли бы к одному роду, а то и семейству.
Естественно, открытие S.inaequale инициировало поиски подобных нематодно-фораминиферных ассоциаций в других морях. В Белом море самый массовый вид агглютинированных фораминифер — Reophax curtus. Домик реофакса состоит из линейно расположенных камер. Начальная камера самая маленькая. По мере роста часть объема цитоплазмы выдавливается из устья и возникает новая камера, больше предыдущей. Живая цитоплазма занимает в основном последнюю, самую крупную камеру. Именно здесь следовало искать необычных паразитов.
В реофаксах мы обнаружили девять видов нематод. Шесть из них известны как свободноживущие обитатели донного осадка, и нам попались, видимо, случайно. Три других обнаруженных в фораминиферах вида — похоже, действительно настоящие внутренние эндобионты простейших. Все три вида оказались новыми для науки, причем два из них — представители неизвестных родов из семейства Camacolaimidae. Пока формальные диагнозы этих нематод не будут опубликованы в академическом “Зоологическом журнале”, мы, согласно Международному кодексу зоологической номенклатуры, не имеем права упоминать их научные названия. Поэтому обозначим их условно: один как Camacolaimus sp. (что означает “вид из рода Camacolaimus”) и второй как Camacolaimidae gen. sp. (“вид и род из семейства Camacolaimidae”).
Самка Camacolaimidae gen. sp. 2 в раковинке реофакса и отдельно при большем увеличении.
Camacolaimus sp. отличается от своих многочисленных свободноживущих родственников в основном размерами: это самый маленький вид рода. И это не удивительно, ведь новый камаколаймус живет в очень тесном пространстве — внутри последней камеры реофакса. Кроме того, бросается в глаза размах изменчивости этого вида как по размерам, так и по морфологическим признакам.
Camacolaimidae gen. sp. 1 еще сильнее отличается от свободноживущих нематод и даже от других представителей камаколаймид: у него нет столь обычного для семейства дорсального зуба, или стилета, и развит половой диморфизм. Самцы — тонкие, стройные и в общем похожи на свободноживущих нематод, а взрослые самки — толстые, веретеновидные и не имеют ничего общего с сородичами.
Самым загадочным оказался третий вид: организация этой нематоды столь своеобразна, что ее трудно отнести к какому-нибудь из существующих семейств. Самцы пока неизвестны, мы нашли только самок и неполовозрелых особей (личинок). На голове не видно никаких органов чувств, ни щетинок, ни амфидов. Ротовая полость невооружена и упрощена. Глотка железистая в задней части. Кишка без клеточных границ и внутреннего просвета, очевидно, она представляет собой сплошной синцитиальный цилиндр. Задняя кишка — рудимент. Половая система самки представлена двумя гонадами, заполненными многочисленными небольшими ооцитами. Судя по строению кишечника, эта нематода не заглатывает пищу ртом, а если и питается во взрослом состоянии, то осмотически, поглощая растворенные органические вещества всей поверхностью тела. Другая особенность, также связанная с обитанием внутри другого живого существа, — чрезмерная плодовитость. Однозначно определить, к какому семейству принадлежит эта форма, нет возможности, так как самцы не найдены (может быть, у этого вида самцы либо очень редки, либо вообще их нет), а у самок отсутствуют сенсиллы — важный диагностический признак. Однако наличия железистой глотки и многоклеточного яйцевода, а также общее сходство с самкой Smithsoninema позволяют предварительно отнести третий вид также к камаколаймидам и обозначить его как Camacolaimidae gen. sp. 2.
В Белом море примерно в 10% раковинок реофакса содержатся нематоды. Черви встречаются как в живых фораминиферах, так и в пустых раковинках. Живые легко распознаются по зернистой липкой цитоплазме, которая становится видна при разламывании домика. Внутри домика нематоды не просто плавают в цитоплазме, а локализованы либо внутри больших пузырей, или вакуолей, либо между внутренней поверхностью стенки раковинки и клеточной мембраной цитоплазмы. Видимо, фораминифера старается изолировать себя от чужого организма, окружая его тонкой кожистой оболочкой, подобной той, которая образуется в начале построения раковинки. Чаще всего в одной особи реофакса живут самец и самка нематоды, как бы семейная пара (если это виды Camacolaimus sp. или Camacolaimidae gen. sp. 1). Редко нематод бывает больше, а соотношение полов отличается от 1:1. И еще реже в одной особи простейшего встречаются больше одного вида нематод.
Код ЕГЭ: 3.1. Разнообразие организмов: одноклеточные и многоклеточные;
автотрофы, гетеротрофы, аэробы, анаэробы
К одноклеточным эукариотам относится множество очень отличающихся друг от друга организмов, которых объединяет один признак — их единственная клетка является в то же время и целым организмом. Хотя в целом они устроены как типичная эукариотическая клетка, однако зачастую могут иметь дополнительные органеллы.
СТРОЕНИЕ. Поверхностный аппарат клетки, отделяющий организм одноклеточного от окружающей среды, зачастую устроен очень сложно. Как и у других клеток, его главная часть — плазмалемма. Надмембранный аппарат может быть представлен гликокаликсом, клеточными стенками различного химического состава, различными чешуйками и домиками (например, как у диатомовых водорослей). Подмембранный комплекс включает различные элементы цитоскелета, именно с ним связано передвижение одноклеточных эукариот. В состав подмембранного комплекса входят основания ресничек и жгутиков, с помощью трансформации элементов цитоскелета происходит движение псевдоподий (ложноножек). С цитоскелетом подмембранного комплекса связаны особые органеллы, которые характерны только для одноклеточных, — экструсомы. Это окружённые мембраной органеллы, которые служат для нападения и защиты.
Ядро у одноклеточных эукариот имеет типичное строение, но у некоторых организмов на протяжении всей жизни или на определённых этапах жизненного цикла в клетке содержится несколько (иногда до сотни) ядер. У инфузорий имеются ядра двух типов: небольшой микронуклеус (генеративное ядро), хранящий генетическую информацию и участвующий в половом процессе, и макронуклеус (вегетативное ядро) — крупное ядро, отвечающее за все процессы жизнедеятельности.
В цитоплазме некоторых одноклеточных эукариот (преимущественно пресноводных) имеются сократительные вакуоли, служащие для осморегуляции. Это одномембранные органеллы, снабжённые выводным каналом, выходящим на поверхность клетки. У инфузорий в состав сократительной вакуоли входит центральный резервуар и радиально расходящиеся канальцы. В сократительную вакуоль поступает жидкость, которая при периодическом сокращении вакуоли выводится наружу.
ПИТАНИЕ. По типу питания среди одноклеточных эукариот имеются как автотрофы, так и гетеротрофы. У автотрофов имеются хлоропласты различной формы (например, чашевидные, лентообразные). Кроме хлорофилла, хлоропласты могут содержать другие пигменты, служащие для лучшего улавливания солнечного света. Гетеротрофные организмы питаются различными органическими частицами или небольшими организмами (бактериями, другими одноклеточными и т. д.). Частицы захватываются при помощи ложноножек в ходе заглатывания частиц (фагоцитоза) или капель (пиноцитоза). У некоторых одноклеточных эукариот имеется особый участок клетки — клеточный рот (цитостом), в котором происходит захват пищевых частиц. Переваривание осуществляется в содержащих пищеварительные ферменты пищеварительных вакуолях (лизосомах).
Тип питания некоторых организмов зависит от образа жизни и среды обитания. Так, эвглена на свету питается автотрофно, производя органические вещества в ходе фотосинтеза, а в темноте переходит к гетеротрофному питанию, поглощая растворённые в воде питательные вещества.
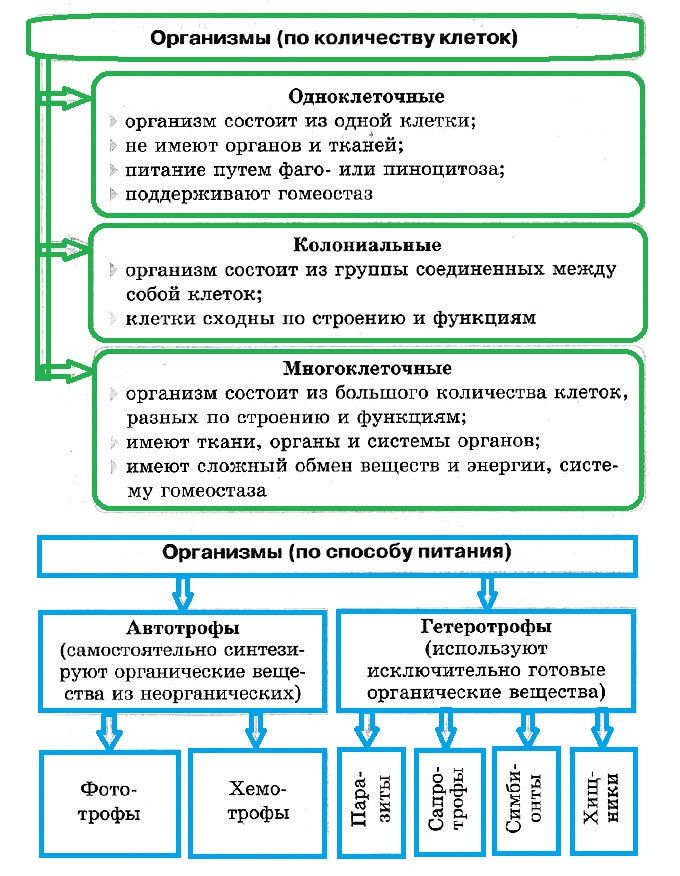
СРЕДА ОБИТАНИЯ. Одноклеточные эукариоты обитают практически повсеместно, уступая в этом отношении только бактериям. Они распространены в пресных и солёных водоёмах, в почве, иногда живут на суше, хотя обычно для них необходима капельная влага. Также часто протисты (другое название одноклеточных эукариот) населяют другие организмы.
Жизнь почвенных одноклеточных обычно имеет две стадии: активную (во время которой происходит питание, рост и размножение) и период покоя. Период покоя наступает вследствие различных причин: недостатка питательных веществ или кислорода, слишком высокой плотности популяции, сухости, накопления различных химических веществ, низкой температуры и др. Хотя существует мнение, что для некоторых видов стадия покоя в жизненном цикле является обязательной. Почвенные одноклеточные принимают участие в почвообразовании и повышают плодородие почв.
В теле многих губок, коралловых полипов, некоторых плоских червей и моллюсков могут обитать водоросли, дающие своим хозяевам кислород и питательные вещества и получающие от них убежище. Такая группа организмов, как лишайники, представляет собой сожительство гриба и водоросли. Обитая в кишечнике различных организмов (термитов и жвачных парнокопытных), они помогают хозяину переваривать пищу.
При паразитизме хозяину наносится вред. Паразитизм среди одноклеточных эукариот распространён довольно широко: они могут вызывать множество заболеваний животных и растений.
Одноклеточные организмы могут объединяться в некое подобие многоклеточного организма, т. е. образовывать колонии. Отдельные особи в колонии могут быть неотличимы друг от друга (некоторые виды зелёных водорослей или инфузорий) или иметь достаточно сильные отличия и даже выполнять различные функции. Колонии образуются в результате бесполого размножения: при делении дочерняя клетка не отделяется от материнской, а остаётся связанной с ней.
Наиболее сложно устроены колонии вольвокса — представителя зелёных водорослей. Это полые шары величиной до 2 мм, они могут включать до 60 тыс. отдельных клеток. По краям колонии находятся двужгутиковые клетки, обеспечивающие передвижение. Кроме них имеются более крупные неподвижные репродуктивные клетки, которые, размножаясь, дают новые колонии. Дочерние колонии развиваются внутри материнской, а затем выходят из неё.
Полагают, что колониальные организмы являются связующим звеном между одноклеточными и многоклеточными организмами, и возникновение многоклеточности происходило через колониальность, причём в разных группах организмов неоднократно.
Тело многоклеточных организмов во взрослом состоянии состоит из множества клеток и их производных (межклеточное вещество). Их клетки различаются по строению и выполняемым функциям, т. е. проявляется дифференциация клеток. Клетки, сходные по строению и происхождению, объединяются в ткани.
Грибы, однако, не имеют настоящих тканей, поэтому некоторыми учёными они не включаются в состав многоклеточных организмов. Из различных тканей образуются органы, которые у многоклеточных животных объединяются в системы органов, выполняющие определённую функцию (дыхание, выделение, пищеварение и т. д.).
Для многоклеточных организмов характерен сложный процесс индивидуального развития (онтогенез). Он начинается в большинстве случаев (за исключением вегетативного размножения) с деления одной клетки — зиготы (оплодотворённой яйцеклетки) — или споры.
Многоклеточность возникала в ходе эволюции неоднократно, она развивалась параллельно у разных групп организмов. Существует несколько гипотез возникновения многоклеточного организма, но все они сходятся в том, что многоклеточность возникла из колониальности.
Многоклеточные организмы могут образовывать колонии, которые образуются в результате вегетативного (бесполого) размножения, когда дочерняя особь остаётся связанной с материнской. Особи в колонии могут быть связаны в разной степени, зачастую их объединяет общее пищеварение. Между отдельными организмами колонии может происходить разделение функций.
По типам питания все живые организмы подразделяются на две группы:
- Автотрофные. К ним относятся фототрофы – зеленые растения, и хемотрофы – некоторые протисты, грибы и бактерии. Это организмы, являющиеся продуцентами, производящие органические вещества из неорганических. Они располагаются схематично на первой ступени экологической пирамиды.
- Гетеротрофные. Это – организмы, питающиеся органическими веществами, произведенными другими их видами. В экологической пирамиде занимаются все уровни, кроме нижнего, на котором расположены автотрофы. В свою очередь гетеротрофные организмы разделяются на консументов – потребителей и редуцентов, разлагающих органику до простых органических и неорганических веществ. При этом, растительноядные животные являются гетеротрофами первого уровня, хищники, поедающие растительноядных – гетеротрофами второго уровня, хищники питающиеся хищниками – третьего и так далее.
По отношению к кислороду живые организмы делятся на четыре большие группы:
- Облигатные аэробы – тех, кто не может жить без кислорода, так как невозможными становятся процессы клеточного дыхания. К ним относятся большинство животных и зеленые растения.
- Микроаэрофилы – это некоторые виды бактерий, которым для жизнедеятельности необходимо небольшое количество кислорода – около 2 %.
- Факультативные анаэробы – живые организмы, которые могут обходиться без кислорода, но способны переключиться на кислородное дыхание. Это маслянокислые и молочнокислые бактерии, дрожжи.
- Облигатные анаэробы – эти организмы гибнут в кислородной среде. К ним относятся хемосинтезирующие бактерии и археи.
Анаэробные бактерии играют важную роль в круговороте вещества, делая его доступным для других участников экологических систем. Биологически же, анаэробный способ получения энергии намного менее эффективен, чем кислородное дыхание. Так, например, при дыхании образуется из одной молекулы глюкозы 38 молекул АТФ, а при бескислородном ее сбраживании – 2 молекулы.
Многоклеточные организмы - это организмы , состоящие [внезапно] из более чем одной клетки , в отличие от одноклеточных организмов.
Все виды животных, наземные растения и большинство грибов являются многоклеточными, как и многие водоросли , тогда как некоторые организмы являются частично одно- и частично многоклеточными, как слизистые плесени и социальные амебы, такие как род Dictyostelium.
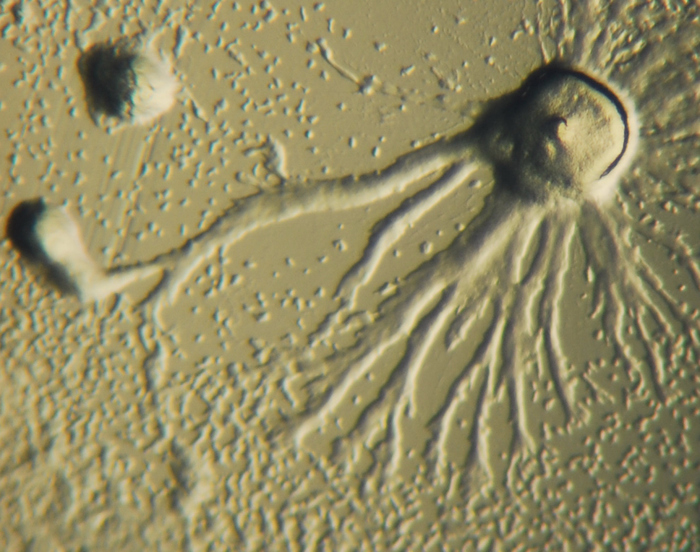
Рис. 1. Колония Dictyostelium в процессе агрегации (Автор Bruno in Columbus)
Многоклеточные организмы возникают различными путями, например, путем деления клеток или путем агрегации (соединения) множества отдельных клеток. Колониальные организмы являются результатом того, что отдельные индивидуальные клетки объединяются в колонию. Однако часто бывает трудно отделить колониальных простейших от настоящих многоклеточных организмов, потому что эти две концепции не различаются.
Многоклеточность развивалась независимо по меньшей мере в 46 раз у эукариот [статья на эту тему на английском языке тут], а также у некоторых прокариот, таких как цианобактерии, миксобактерии, актиномицеты, у экстремофильной бактерии Magnetoglobus multicellularis и у группы метановых архебактерий Methanosarcina.
Однако сложные многоклеточные организмы развивались только в шести эукариотических группах: животные, грибы, бурые водоросли, красные водоросли, зеленые водоросли и наземные растения. Мноклеточность неоднократно развивалась для Chloroplastida (зеленые водоросли и наземные растения), один или два раза для животных, один раз для бурых водорослей, три раза в грибах (хитриды, аскомицеты и базидиомицеты) и, возможно, несколько раз для слизистых плесени и красные водоросли.
Первым доказательством многоклеточности являются цианобактериоподобные организмы, которые жили 3–3,5 миллиарда лет назад.
Некоторые группы организмов в своем эволюционном развитии утратили многоклеточность.
Многие грибы (например, Saccharomycotina, Cryptococcus и другие дрожжи), некоторых красные водоросли (например, Porphyridium sp.), но возможно, что они примитивно одноклеточные.
Потеря многоклеточности также считается вероятной у некоторых зеленых водорослей (например, Chlorella vulgaris и некоторые из группы Ulvophyceae). В других группах, как правило, паразитах, происходило уменьшение количества клеток или уменьшение типов клеток (например, многие представители Myxozoa, многоклеточные организмы, ранее считавшиеся одноклеточными, являются паразитами морских животных).
Одна из гипотез о происхождении многоклеточности заключается в том, что группа функционально-специфических клеток объединяется в слизистую массу, которая перемещается как многоклеточная единица. Это в основном то, что делают слизевики.
Другая гипотеза состоит в том, что примитивная клетка подвергалась делению ядра, превращаясь в коеноцит (клетку с множеством ядер). Мембрана, в таком случае, сформировалась бы вокруг каждого ядра (и клеточного пространства и органелл, занятых в пространстве), таким образом приводя к группе связанных клеток в одном организме. Этот механизм наблюдается у дрозофилы.
Третья гипотеза состоит в том, что при разделении одноклеточного организма дочерние клетки не могли отделиться, что привело к конгломерации идентичных клеток в одном организме, что впоследствии могло развить специализированные ткани. Это то, что делают эмбрионы растений и животных, а также колониальные хоанофлагелляты.
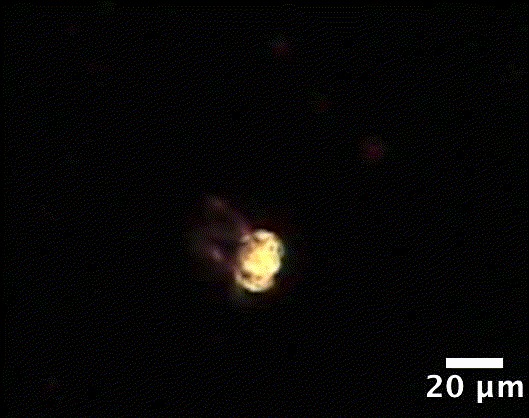
Гиф. 1. Tetrabaena socialis - зеленая водоросль, самый примитивный многоклеточный организм, состоящий из четырех клеток, несущих по два жгутика
Поскольку первые многоклеточные организмы были простыми, мягкими организмами, у которых не было костей, раковин или других твердых частей тела, они плохо сохранились в ископаемом материале. Одним из исключений может быть Обыкновенная губка (Demospongiae), которая, возможно, оставила химический след в древних породах.
Самые ранние окаменелости многоклеточных организмов включают Grypania spiralis (см. рисунок 1) и окаменелости черных сланцев палеопротерозойской фервильской группы в Габоне (2,5 млрд лет назад).

Рис. 2. Grypania spiralis в отложениях
Далее я более подробно расскажу о современных теориях возникновения многоклеточности:
Эта теория предполагает, что первые многоклеточные организмы произошли от симбиоза (кооперации) различных видов одноклеточных организмов, каждый из которых играет разные роли. Со временем эти организмы станут настолько зависимыми друг от друга, что не смогут выжить независимо, что в конечном итоге приведет к объединению их геномов в один многоклеточный организм. Каждый соответствующий организм стал бы отдельной линией дифференцированных клеток во вновь создаваемых видах.
Однако проблема этой теории заключается в том, что до сих пор неизвестно, как ДНК каждого организма может быть включена в один геном, чтобы составить их как единый вид. Хотя считается, что такой симбиоз имел место (например, митохондрии и хлоропласты в клетках животных и растений - эндосимбиоз), он случался очень редко, и даже тогда геномы эндосимбионтов сохраняли элемент различия, раздельно реплицируя свою ДНК во время митоза видов-хозяев.
Например, два или три симбиотических организма, образующих составной лишайник , хотя и зависят друг от друга в отношении выживания, должны воспроизводиться по отдельности, а затем переформироваться, чтобы создать еще один отдельный организм.
Эта теория утверждает, что один одноклеточный организм с несколькими ядрами мог бы создать внутренние мембранные перегородки вокруг каждого из своих ядер.
Многие протисты, такие как инфузории или слизевики, могут иметь несколько ядер, что подтверждает эту гипотезу. Однако простого наличия нескольких ядер недостаточно для поддержки теории. Множественные ядра инфузорий различны и имеют четко дифференцированные функции. Макроядро служит потребностям организма, а микроядро используется для полового размножения с обменом генетического материала.
Синцитиальные слизевики образуются из отдельных амебоидных клеток, подобно синцитиальным тканям некоторых многоклеточных организмов, а не наоборот.
Чтобы считаться действительной, эта теория нуждается в наглядном примере и механизме образования многоклеточного организма из ранее существовавшего синцития.

Рис. 3. Слизевик Fuligo septica (Автор фото Kreis Tuttlingen)
Теория предполагает, что симбиоз многих организмов одного и того же вида (в отличие от симбиотической теории , которая предполагает симбиоз разных видов) привел к многоклеточному организму.
Преимущество колониальной теории состоит в том, что она была обнаружена независимо в 16 различных типах простейших. Например, во время нехватки пищи амеба Dictyostelium (Рис. 1) группируется в колонии, которая перемещается как единое целое в новое место. Некоторые из этих амеб затем немного отличаются друг от друга.
Другими примерами колониальной организации у простейших является, например, Volvox sp. (Рис. 4), который состоит из 500-50000 клеток (в зависимости от вида), только часть из которых размножается.
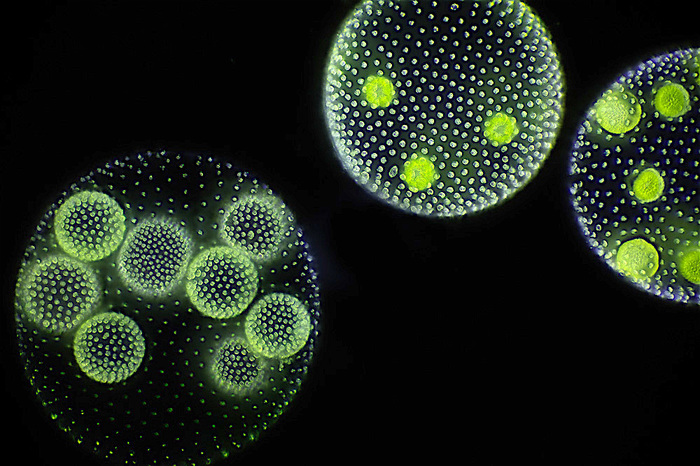
Рис. 4. Колонии вольвокса (Автор фото Frank Fox)
В отечественной литературе колониальную теорию обычно делят на теорию гастреи (Геккель, 1872), теорию фагоцителлы (Мечников, 1878) и теорию синзооспоры (Захваткин, 1949). Обе теории довольно схожи, разница в том, как, согласно эти теориям, происходила интеграция клеток в колонии.
Согласно этой теории предком многоклеточных была гастрея - многоклеточный двуслойный организм. Она произошла от колониальных протистов с шарообразными колониями. Процесс интеграции клеток в колонии сделал возможным разделение функций между клетками: передние клетки утрачивают жгутики и превращаются в фагоциты, сидящие во впячивании на переднем конце - образуется кишечник.Остальные клетки утрачивают пищеварительную функцию и становятся чисто двигательными. Рот гастреи находился на переднем конце, и пища "сама заплывала" в кишечник. Симметрия у гастреи была радиальной. При переходе к сидячему образу жизни ее потомки эволюционировали в губок и кишечнополостных, а при переходе к ползанию по дну - в плоских червей и всех остальных многоклеточных (Рис. 5).
Так развиваются ланцетники и коралловые полипы.
Фагоцителла не имела рта и кишечника, пищеварение было внутриклеточное. Рот сформировался,как просвет между клетками наружного слоя, ведущий во внутреннюю паренхиму. Располагался он, в отличие от гастреи на заднем конце тела. Кишечника еще не было. Но теперь возникла возможность питаться более крупной добычей: внутренние клетки могли окружать ее, образуя гигантскую пищеварительную вакуоль. Однако для хищничества нужна еще способность ловить добычу. Поэтому хищничать научились только настоящие многоклеточные - после того, как у них возникли мышцы и управляющая ими нервная система. Постепенно у потомков фагоцителлы сформировался постоянный кишечник. По мере увеличения размеров он мог усложняться: возникли боковые карманы, чтобы доставлять пищу к наружным слоям клеток. В дальнейшем у некоторых животных эти карманы могли отделиться, дав начало полости тела - целому. Фагоцителла обитала в толще воды. Нетрудно представить себе, как от нее могли произойти современные группы животных при переходе к жизни на дне. Когда рта еще не было, осевшая на дно фагоцителла "превратилась" в трихоплакса. После появления рта, но до появления кишечника при переходе к ползанию возникли бескишечные турбеллярии. Рот у них сместился на брюхо, и они стали двустороннесимметричными. После появления кишечника часть потомков фагоцителлы перешли к сидячему образу жизни на дне - они превратились в кишечнополостных (Рис. 5).
Так развиваются обыкновенные губки, гидроидные и сцифоидные стрекающие.
Основным кандидатом на роль предка всех многоклеточных является воротничковый жгутиконосец (Choanoflagellata), их клетки снабжены жгутиком, окруженный воротничком. Жгутик создает токи жидкости, позволяющие хоанофлагеллятам плавать в толще воды. Эти же токи пригоняют в воротничковую зону бактерий, которыми хоанофлагелляты питаются. Многие их этих простейших образуют колонии, причем легко переходят из одноклеточного состояния в колониальное и обратно.
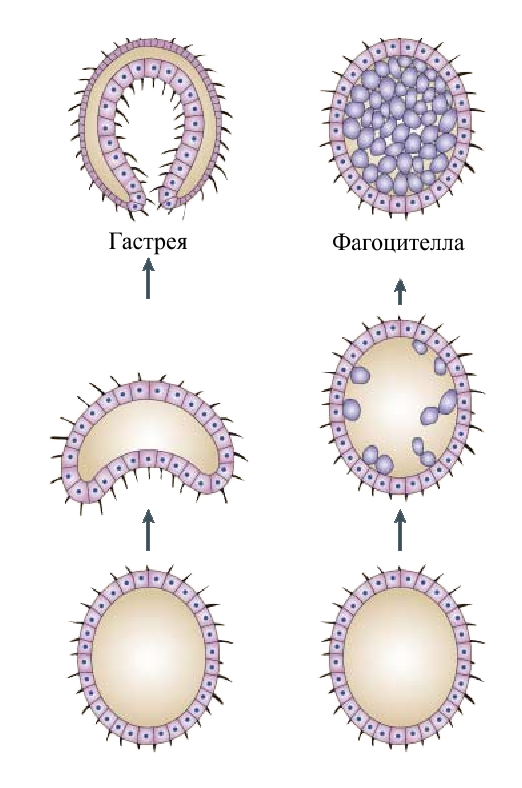
Рис. 5. Гастрея и фагоцителла
Согласно данной теории многоклеточные также произошли от колониальных протистов. У протистов встречаются клетки, сильно увеличенные за счет запасания питательных веществ - как яйцеклетка у животных. Часто такие клетки делятся несколько раз подряд - это похоже на дробление. Таким способом образуются одноклеточные мелкие расселительные стадии - зооспоры.
У колониальных протистов зооспоры могут оставаться все вместе, образуя колонию - синзооспору. В процессе эволюции могла произойти неотения и утратиться взрослая сидячая стадия.
Отличия от теорий фагоцителлы и гастреи:
- Считается, что никогда не существовало однослойного шарообразного предка. Об этом свидетельствует то, что у всех многоклеточных бластулы не питаются. Не питаются и образующиеся из них паренхимулы. Поэтому и у древних многоклеточных такие стадии не были взрослыми организмами - это были всегда только личинки.
- Сидячий образ жизни примитивных взрослых многоклеточных.
- Наиболее примитивной из расселительных личинок считается бластула. В паренхимулу она превращается, готовясь к превращению во взрослый организм. Этот метаморфоз происходит после перехода к сидячему образу жизни. У всех остальных многоклеточных взрослая сидячая стадия утратилась. У этих животных личинки стали взрослыми - произошла неотения.
Многоклеточность позволяет организму превышать пределы размера, обычно налагаемые диффузией: отдельные клетки с увеличенным размером имеют уменьшенное отношение поверхности к объему и испытывают трудности с поглощением достаточного количества питательных веществ и их транспортировкой по всей клетке.
Таким образом, многоклеточные организмы имеют конкурентные преимущества увеличения размера без его ограничений. Многоклеточность также позволяет увеличивать сложность, позволяя дифференцировать типы клеток в пределах одного организма.
Я попытался привести современные данные по возникновению многоклеточности. Тема спорная, вариантов куча и все они интересны. Есть, что добавить - пиши в комменты.

Нашел у себя косяк в абзац после рис. 4. Читайте не "обе теории", а "все три". Не вычитал при редактуре.
- Сидячий образ жизни примитивных взрослых многоклеточных.
Очень интересно) ещё бы было побольше примеров с картинками) так, таким малообразованным, как я, понятнее))
Блин, игра такая была, где нужно было другие клетки и ядра поглощать. Не Spore, попроще. Кто помнит?
всё чаще посещают мысли, что скоро в метод.указаниях по зооллгии буду на пикабу студентов посылать: и написано яснее, и актуальнее, и ссылки с доказательствами на публикации есть. Только вот почему-то это ни разу не смешно, а даже грустно.
Эти зеленые водоросли всю жизнь в аквариуме портят, то вылазят то пропадают))

Чувствуете, пахнУло курсом беспов и Малаховым?
Я поддерживаю теорию гастреи.
А что не теорию фагоцителлы? Только фагоцителла, только хардкор
А может кто помнит мини игру где просто выставляешь параметры для своих клеток (скорость размножения, выживаемость, сила) и можно было соревноваться с другими клетками.
Мне напомнило Spore 2008г. там развитие с клетки до покорны космоса
Все виды животных . являются многоклеточными
Однако стоит загуглить
как возникает мысль, что кто-то пиздит: либо гугл, либо ТС.
Инфузория - протист или простейшее, современная систематика не относит протист к животным.
ну, это кто как: современная систематика - она пока ещё сильно резиновая
Фотосинтезирующий моллюск!
Модель межорганизменных взаимоотношений моллюска элизия Elysia viridis и морских водорослей Codium fragile.
* Elysia viridis - морской моллюск размером 3 см, обитает на атлантическом побережье Северной Америки.

* Связь трофической цепи (цепь питания) и биохимического симбиоза: морской моллюск Elysia viridis питается водорослью Codium fragile.

Моллюск ухитряется переселить хлоропласты съеденных водорослей в свои собственные клетки, располагающиеся вдоль кишечника, и долгое время сохранять их там живыми, приобретая, таким образом, способность к фотосинтезу.
Данный вид живет благодаря субклеточному взаимодействию с хлоропластами, полученными от водоросли. Последние обеспечивают моллюска продуктами фотосинтеза. Моллюск питается водорослью и абсорбирует из нее хлоропласты.
Симбиоз морских и ряда пресноводных животных и водорослей
Множество морских и некоторые пресноводные животные (радиолярии, кораллы, актинии, медузы, гигантский двустворчатый моллюск тридакна и др.) содержат в своем теле одноклеточные водоросли: золотистые зооксантеллы. В то же время зеленая гидра, плоский червь конволюта, некоторые губки и др. содержат одноклеточные водоросли зеленые зоохлореллы.
Симбиоз водоросли и животного
Организм-хозяин выставляет на яркий свет своих симбионтов, которые фотосинтезируя и подпитываясь поставляемыми хозяином соединениями азота и фосфора, производят органические вещества. Этим питаются животные и в случае излишек симбионтов в организме - их переваривают
Механизм взаимодействия моллюска и водоросли
Существует строгая специфичность взаимодействия:
зеленая элизия (E.viridis) поедает только водоросль Codium tomentosum, черно-зеленая (E.atroviridis) – лишь C.fragile, а зеленоухая (E.chlorotica) – вошерию (Vaucheria litorea).
Прокалывая стенку водоросли моллюск глоткой выкачивает ее содержимое. Жидкость переваривается, а хлоропласты проникают в густо расположенные под кожей печеночные выросты и хранятся там в одной-двух (на вырост) специальных крупных клетках эпителия.
Геном моллюска контролирует работу хлоропластов водоросли, синтезируя недостающие им белки.


Почему у человека некоторые органы — парные (например, легкие, почки), а другие — в одном экземпляре?

Иллюстрация, показывающая различные типы симметрий, существующие на Земле. Демонстрируется в Музее естественной истории имени Филда в Чикаго, США. Здесь показаны и те виды симметрии, которые не были описаны в этой статье, поскольку не встречаются у человека (например, спиральная симметрия раковин моллюсков). Изображение с сайта en.wikipedia.org
Вначале попробуем ответить на вспомогательный вопрос: почему у человека некоторые части тела симметричны, а другие — нет?
Симметрия — базовое свойство большинства живых существ. Быть симметричным очень удобно. Подумайте сами: если у вас со всех сторон есть глаза, уши, носы, рты и конечности, то вы успеете вовремя почувствовать что-то подозрительное, с какой бы стороны оно ни подкрадывалось, и, в зависимости от того, какое оно, это подозрительное, — съесть его или, наоборот, от него удрать.

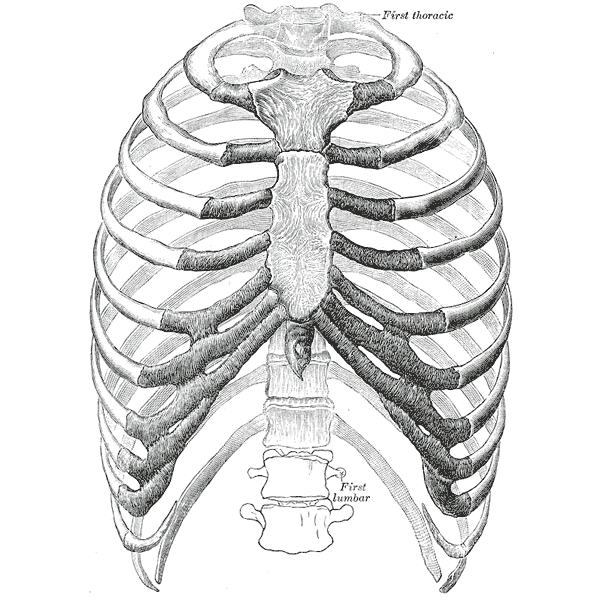
Человеческие ребра имеют некоторые черты метамерной симметрии. First thoracic — первый грудной позвонок, first lumbar — первый поясничный. Изображение с сайта ru.wikipedia.org
Итак, почему у человека есть парные органы, мы разобрались. Теперь обсудим, откуда взялись непарные.
Теперь посмотрим на кровеносную систему.
Читайте также: