
Один шейный позвонок характерный признак класса рыб
Земноводные, или амфибии (от греч. amphi - с обеих сторон и греч. bios - жизнь) - класс типа хордовые, объединяющий животных, которые могут обитать как в воде, так и на суше. Это немногочисленный наиболее примитивный класс наземных позвоночных животных. Произошли земноводные от древних кистеперых рыб.
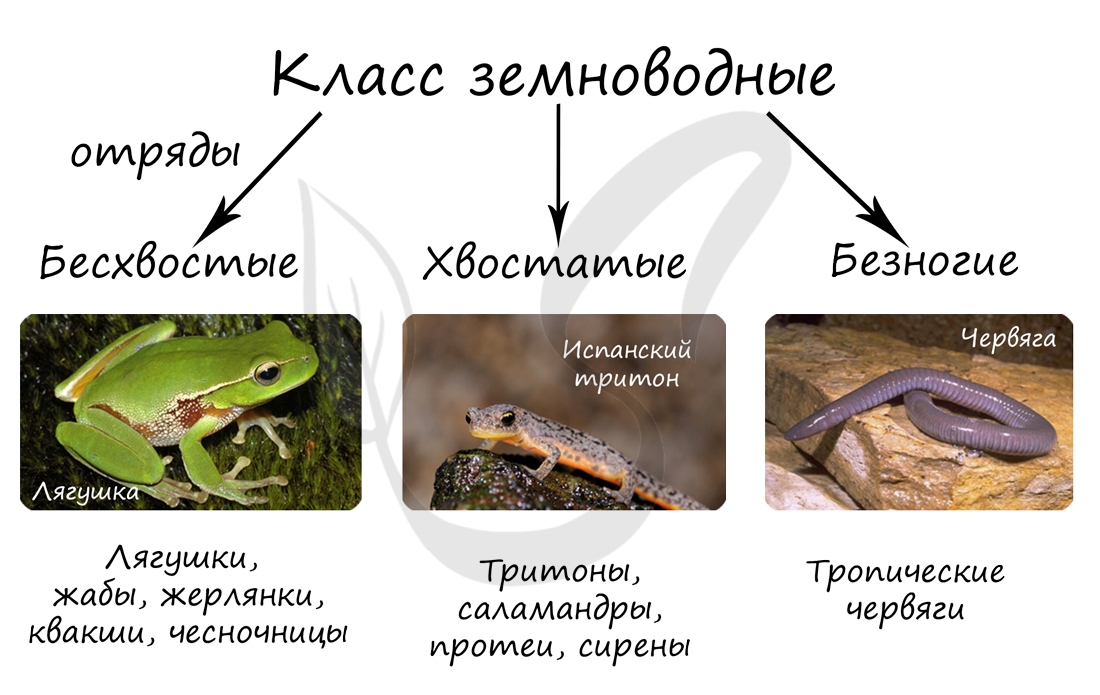
Для успешного изучения земноводных предлагаю начать с их классификации. Помните, что классификации это именно то, что раскладывает знания в голове "по полочкам", относитесь к ним с должным вниманием.
Давайте перечислим ароморфозы, которыми сопровождалось появление земноводных.
- Появление конечности пятипалого типа
У ранее обсуждаемой нами рыбы латимерии (целоканта) плавники напоминали конечность пятипалого типа. У земноводных они преобразовываются в скелет передней и скелет задней конечностей, которые мы обсудим в данной статье подробно.

Мышцы дифференцируются (делятся) на мышечные пучки, становятся возможны более сложные и точные движения.

Легкие земноводных представлены в виде тонкостенных мешков, относительно небольшого объема.
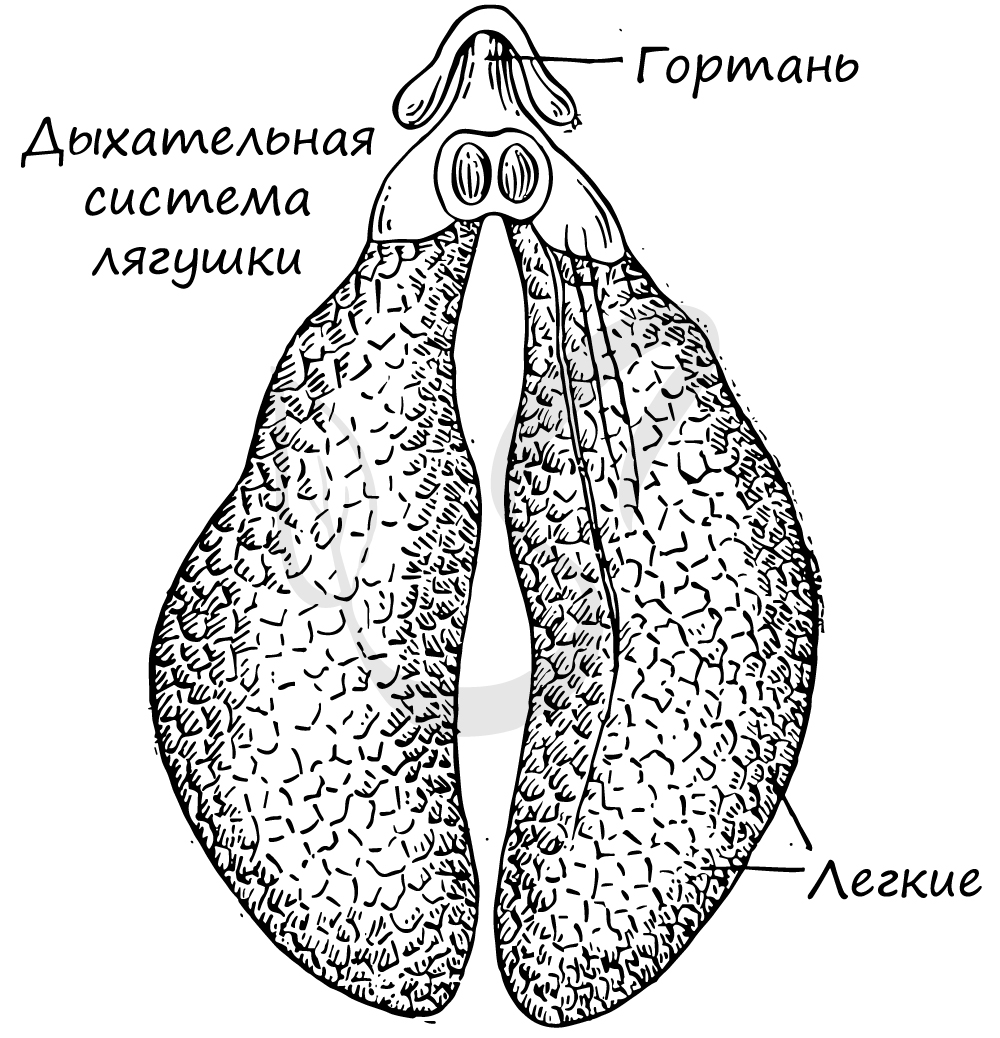
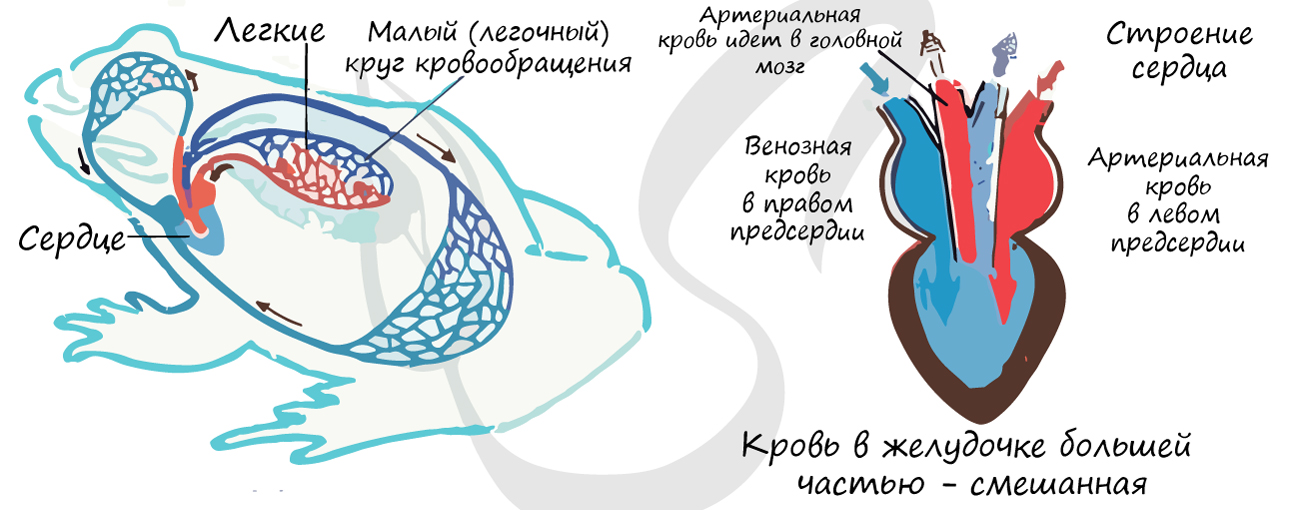
Появление легочного круга кровообращения связано с тем, что земноводные осваивают новый тип дыхания, наземное дыхание - дыхание атмосферным воздухом.
Появление шейного и крестцового отделов позвоночника
Благодаря появлению шейного отдела становится возможным поворот головы, что было невозможным для рыб.

Традиционно строение земноводных изучается на примере типичного представителя - лягушки озерной, ее мы возьмем за основу данной статьи.

- Покровы, опорно-двигательная система
Покров двуслойный, состоит из эпидермиса и дермы. Эпидермис снабжен густой капиллярной сетью, благодаря которой у лягушки возможен газообмен через кожу. Кожа голая, содержит большое количество желез: она должна быть постоянно увлажнена для интенсивного газообмена.
Около 65% всего кислорода поступает через увлажненную кожу. Увлажнение кожи крайне важно, рекомендую запомнить следующий факт: при пересыхании кожи лягушки газообмен в ней прекращается, и лягушка погибает от недостатка кислорода (гипоксии).

Особой окраской обладают ядовитые виды лягушек, их яркая окраска называется - предупреждающей. Съев такую, хищник серьезно пожалеет о своей оплошности, которая может закончиться летальным исходом. Поэтому животных с такой окраской хищники обычно не трогают.
Индейцы используют лягушек кокои (листолаз ужасный) для изготовления отравленных стрел: одна лягушки делает смертельным оружием около 50 стрел. Они готовят яд высушивая лягушку на костре и смазывая наконечники стрел выступающей на поверхности коже жидкостью - ядом.

Тело состоит из головы, туловища и двух пар конечностей. Передние конечности короче и слабее задних. Задние конечности служат для плавания, они удлинены и имеют плавательные перепонки между пальцами. Хорошо развита мышечная система: мышцы дифференцированы на отдельные пучки. В теле амфибий насчитывается до 350 мышц.
Скелет лягушки состоит из скелета головы, туловища, поясов конечностей и свободных конечностей. Скелеты поясов конечностей малоподвижны, в отличие от скелета свободной конечности, построенного по типу рычагов.
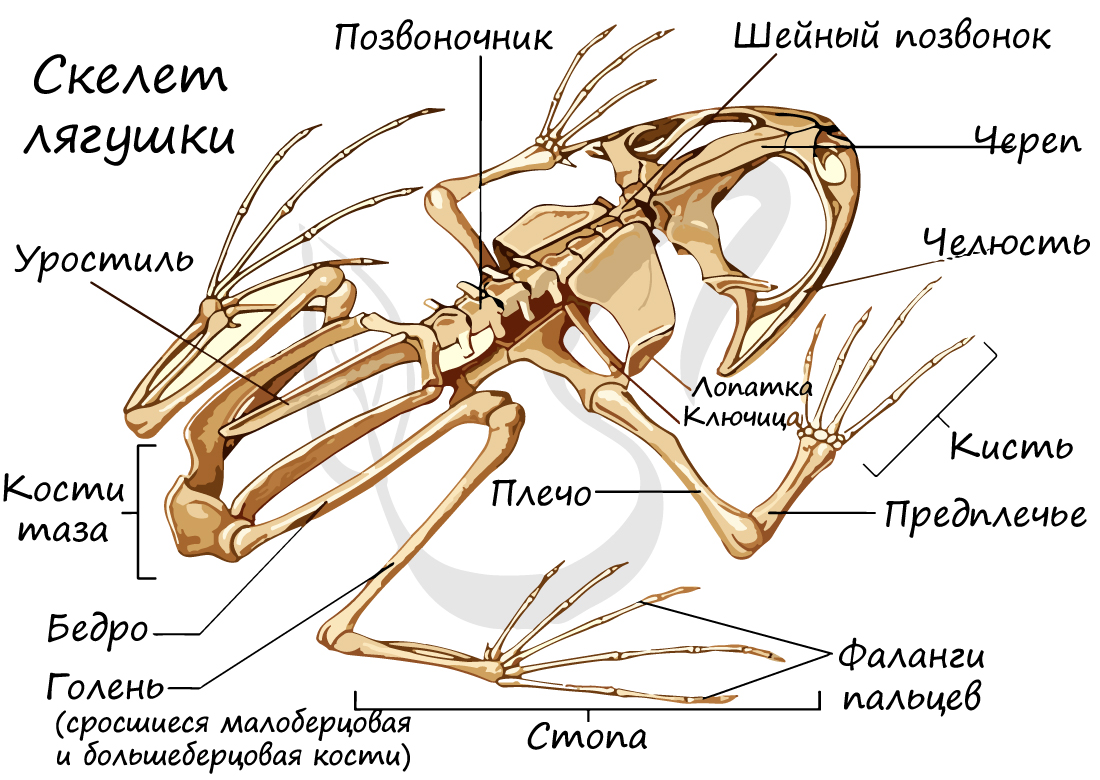
Имеются туловищный (7 позвонков) и хвостовой отделы позвоночника, доставшиеся земноводным еще от рыб. Однако появляются и новые отделы: крестцовый и шейный, оба содержащие по одному позвонку. Благодаря возникновению шейного отдела, у земноводных становится возможным поворот головы, что было невозможным для рыб.
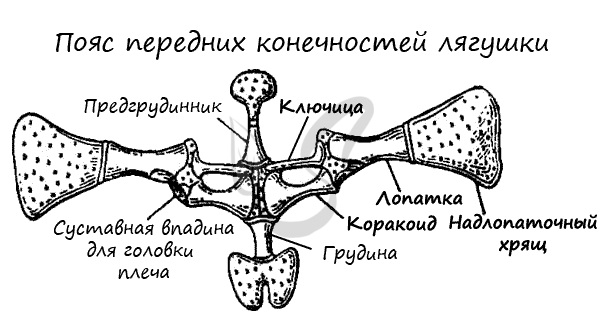
Скелеты поясов конечностей служат опорой для конечностей. В состав плечевого (переднего) пояса входят: лопатки, ключицы, вороньи кости (коракоиды), хрящи. Ребер нет, вследствие чего грудная клетка отсутствует, и передний пояс лежит свободно в толще мышц.
Пояс задних конечностей (тазовый) образован: подвздошными и седалищными костями, лобковым хрящом.
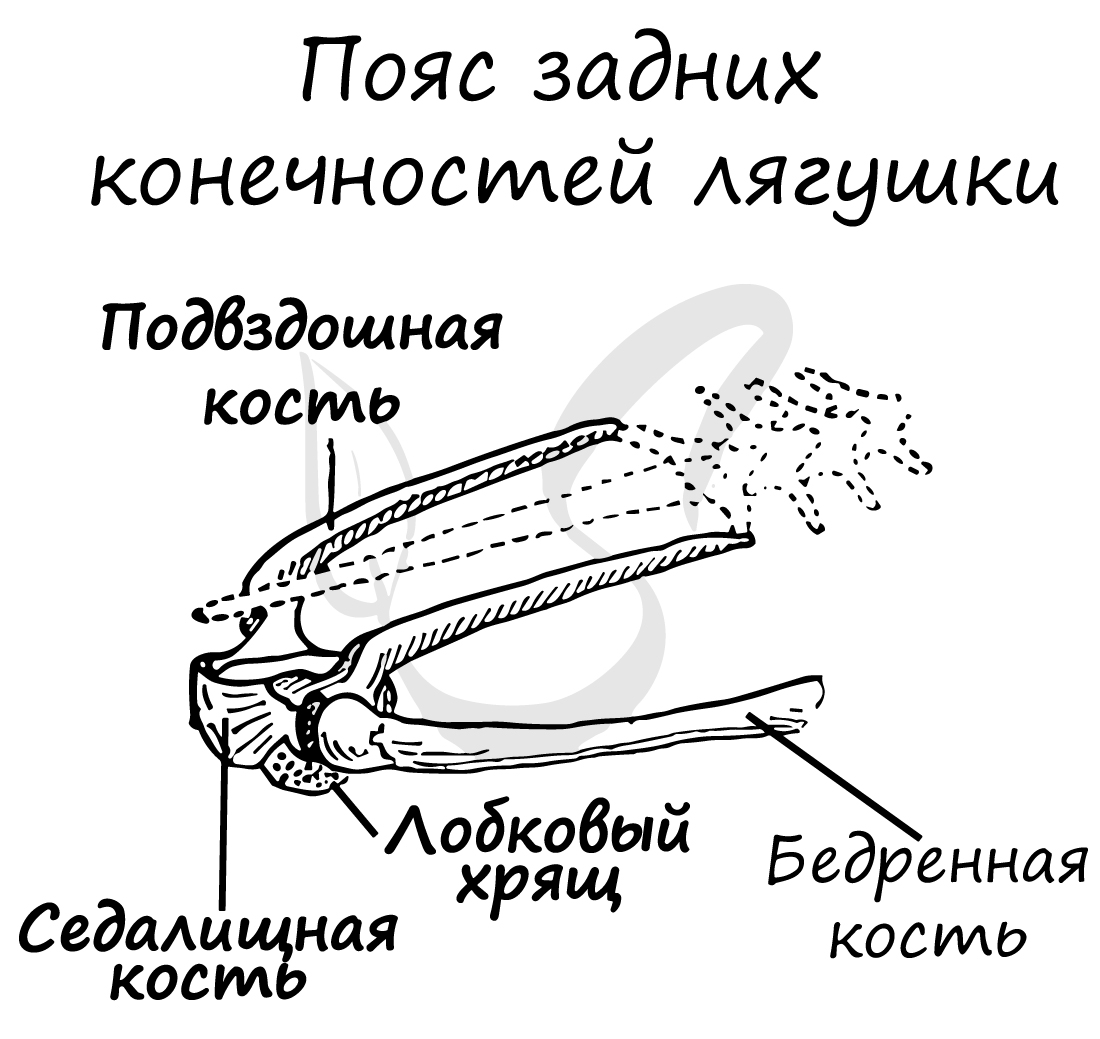
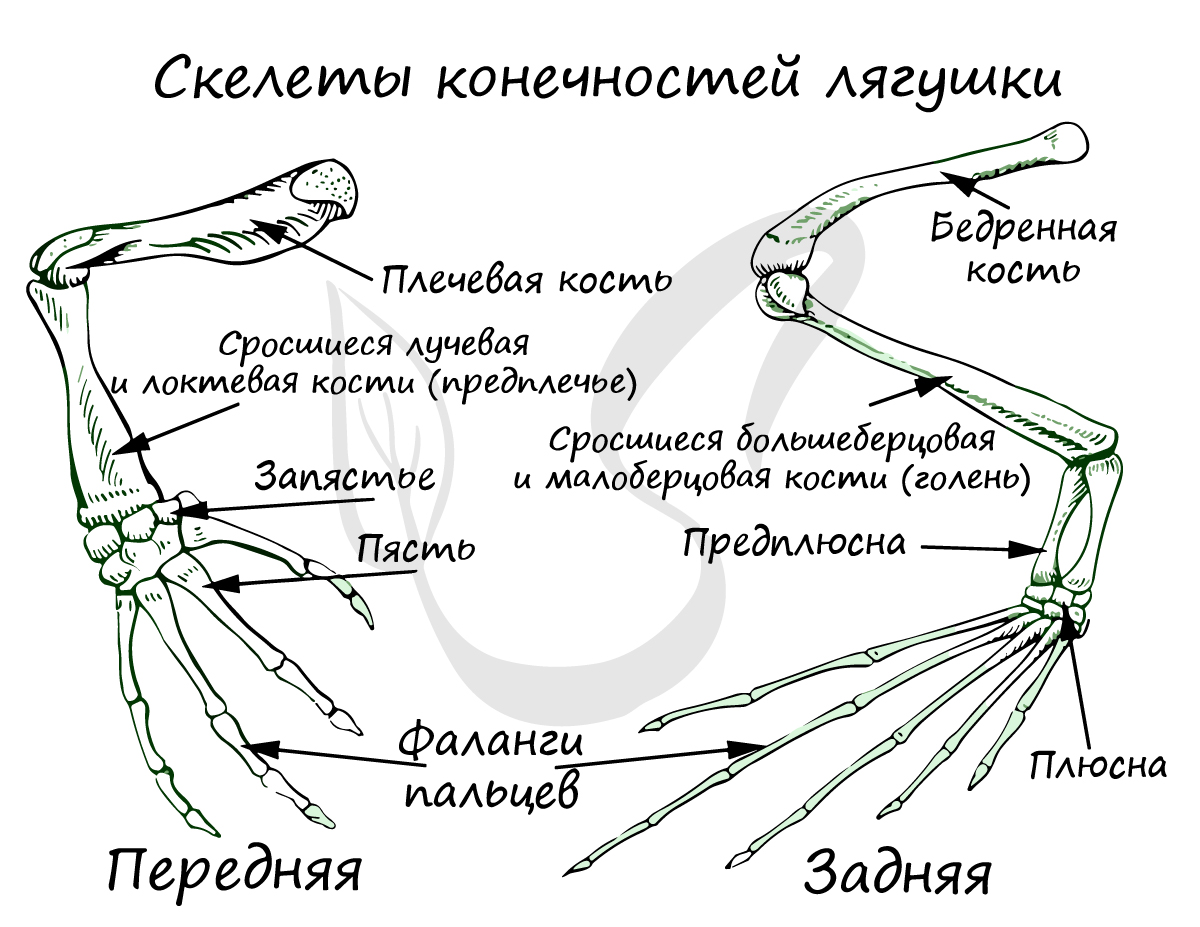
Скелет передней свободной конечности состоит из: плечевой кости, предплечья (образовано сросшимися лучевой и локтевой костями) и кисти (состоящей из запястья, пястья и фаланг пальцев).
Скелет задней свободной конечности образован бедром, голенью (состоит из сросшихся малоберцовой и большеберцовой костей), стопы (включает предплюсну, плюсну и фаланги пальцев).
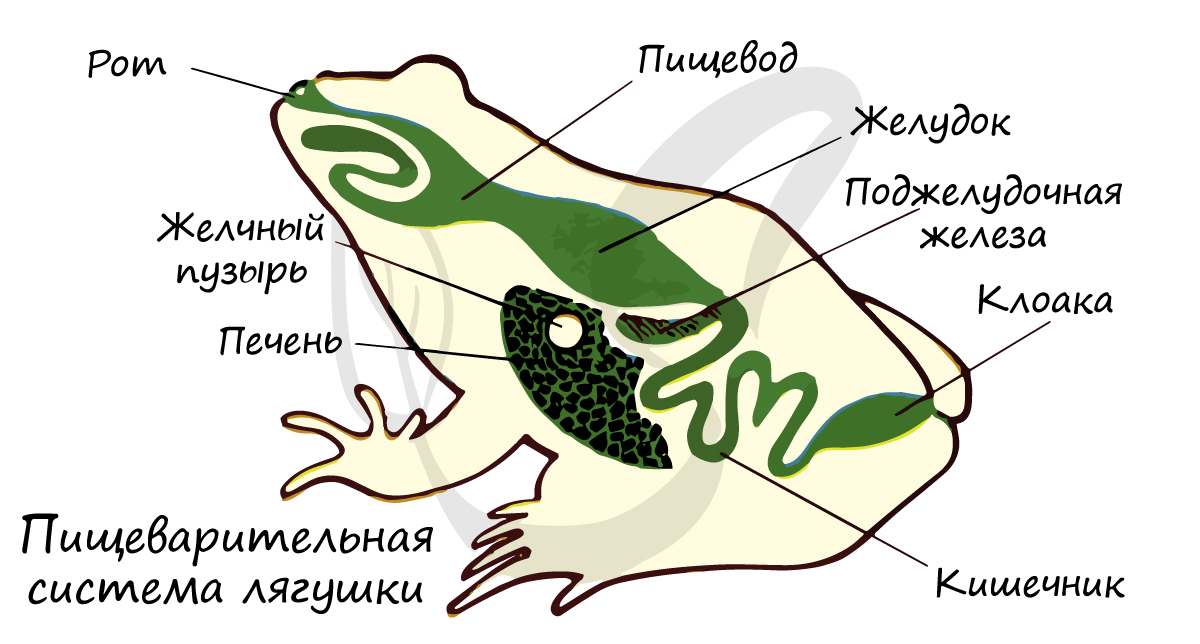
Состоит из ротоглоточной полости, в которую открываются протоки слюнных желез, секрет которых предназначен только для смачивания и формирования пищевого кома. Слюнные железы впервые возникли именно у амфибий, и пока еще не участвуют в химической обработке пищи. Слюнные железы были и у насекомых, однако членистоногие - отдельная эволюционная ветвь, хордовые никаким образом не произошли от членистоногих :)
Имеются хоаны - отверстия, с помощью которых полость носа сообщается с полостью глотки. В ротоглоточную полость открываются также отверстия евстахиевых труб, которые соединяют полость среднего уха с ротоглоткой и уравнивают в них давление. Значительную часть места занимает липкий длинный язык, служащий для ловли добычи.

Короткий пищевод переходит в желудок, продолжающийся в тонкий кишечник, куда открываются протоки желчного пузыря, а также пищеварительных желез: печени и поджелудочной. Тонкая кишка переходит в толстую кишку, оканчивающуюся клоакой.
Клоака (от лат. cloaca - труба для стока нечистот) - расширенная часть задней кишки, куда открываются мочеточники и мочевой пузырь, половые протоки.
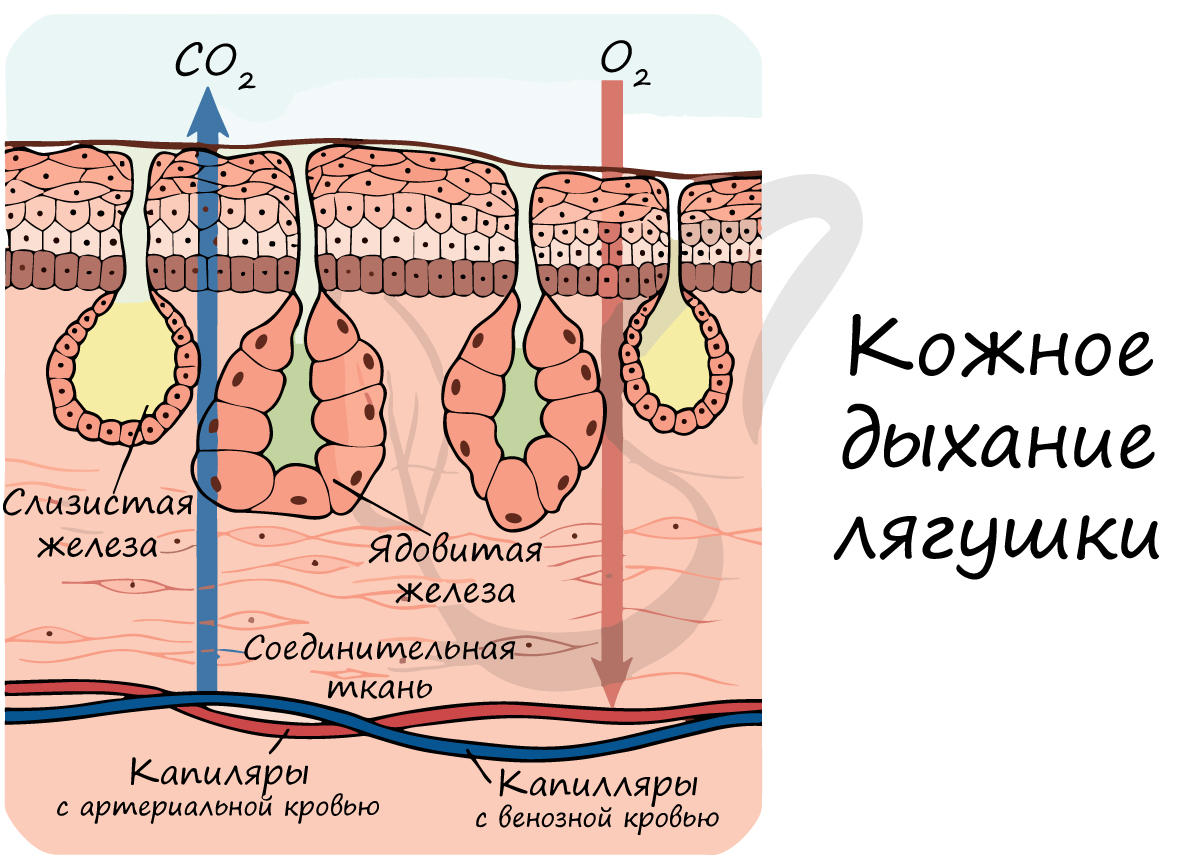
У лягушки дыхание осуществляется двумя основными способами:
-
Легочное дыхание
Легкие представленные тонкостенными мешками, стенка которых густо оплетена капиллярами, в которых происходит газообмен. Дыхательная поверхность легких невелика, они не способны полностью удовлетворить нуждам организма в кислороде, поэтому жизнь лягушки невозможна без кожного дыхания.
Воздух поступает в легкие и удаляется из них благодаря сокращениям мышц дна ротовой полости.

Осуществляется только при увлажненной коже, на поверхности которой растворяется кислород, после чего он проникает в капилляры кожи. Кожа помогает дышать лягушкам при погружении в воду: кислород поступает в кровь напрямую из воды.
При пересыхании кожи животное погибает, так как слабо развитые легкие не в состоянии обеспечить потребности организма в кислороде.
Также дыхание осуществляется эпителием, покрывающим ротовую полость, но это вносит незначительный вклад в процесс дыхания.
Изменения в кровеносной системе земноводных, по сравнению с рыбами, обусловлены заменой жаберного дыхания на легочное. Сердце имеет 3 камеры: 2 предсердия и 1 желудочек, сердце более крупное. Возникает второй (легочный) круг кровообращения.
Правое предсердие заполнено венозной кровью, а левое - артериальной. Перегородка в желудочке отсутствует, кровь внутри него большей частью смешанная. Из-за смешения крови интенсивность метаболизма уменьшается, это делает невозможным поддержание постоянной температуры тела. Эритроциты у амфибий с ядрами, безъядерные эритроциты возникнут позже, у млекопитающих.
Земноводные относятся к пойкилотермным животным (от греч. poikilos - различный и therme - тепло) - холоднокровным животным, температура тела которых непостоянна и зависит напрямую от температуры окружающей среды.
Физиологически и анатомически сокращение сердца происходят таким образом, что более насыщенная кислородом артериальная кровь изгоняется из него в последнюю очередь и попадает в сонные артерии, идущие к головному мозгу. Таким образом, к головному мозгу направляется более насыщенная кислородом кровь, чем к остальным органам.
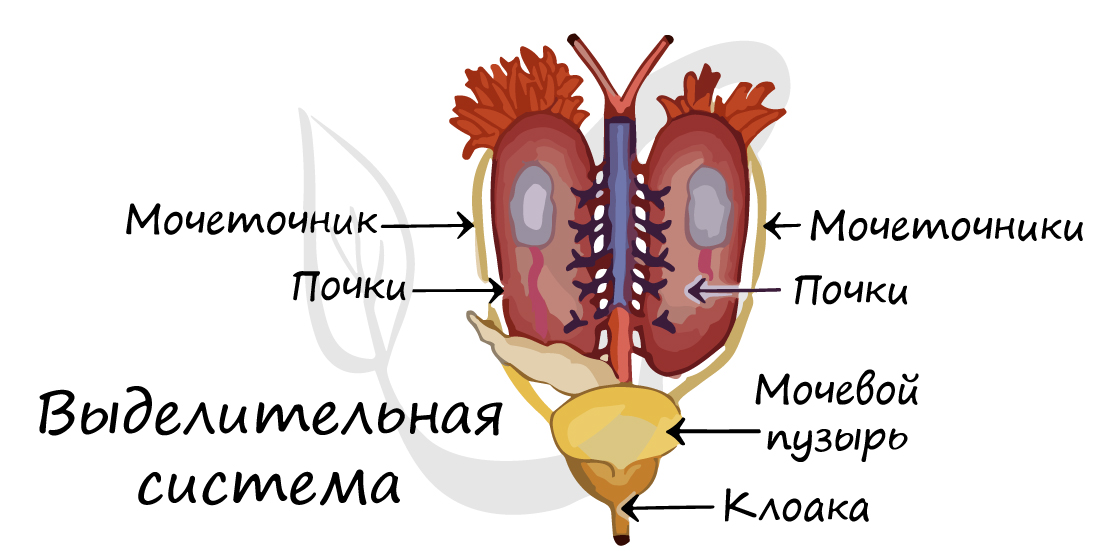
Так же, как и у рыб, выделительная система амфибий состоит из парных первичных (туловищных) почек, называемых мезонефрос. В них фильтруются продукты обмена веществ, главным образом - мочевина.
По мере образования в почке мочи, она поступает в мочеточники, ведущие в клоаку. Попавшая в клоаку моча стекает в мочевой пузырь, через стенки которого осуществляется всасывание воды. При сокращении стенок мочевого пузыря концентрированная моча выводится в клоаку, откуда выбрасывается во внешнюю среду.
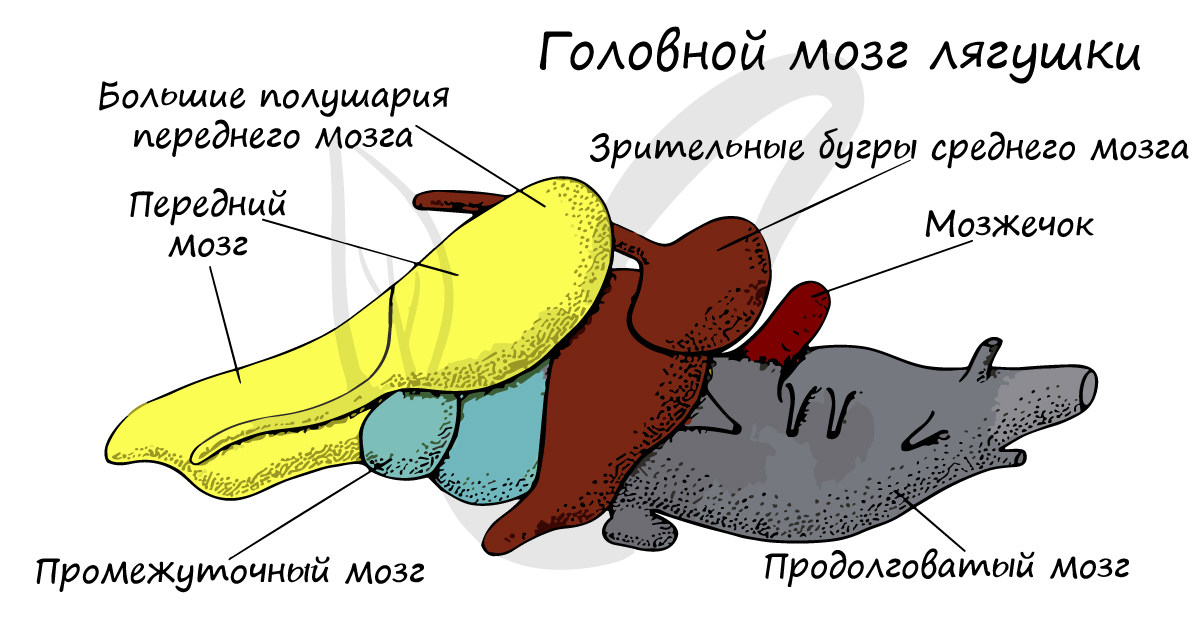
Головной мозг земноводных отличается от мозга рыб полностью разделенным на два полушария и более развитым передним мозгом.
Мозжечок и средний мозг развиты гораздо слабее, так как земноводные малоподвижны и их движения относительно простые: они передвигаются по плоскости в "2D измерении". Перемещение рыб в таком случае можно обозначить как в "3D измерении". Это упрощенное, но вполне справедливое сравнение.
Органы чувств земноводных адаптированы к наземному образу жизни. На суше возникает серьезная проблема, которой не было у рыб в воде - сухой воздух. Чтобы адаптироваться к нему у земноводных возникает подвижное веко и мигательная перепонка, движения которых смачивают поверхность глаза и препятствуют высыханию.
Меняется форма хрусталика. Если у рыб он был шарообразный, то у земноводных его форма становится более выпуклой (как и форма роговицы). Хрусталик приобретает форму двояковыпуклой линзы. Амфибии видят дальше рыб.
Однако аккомодация, настройка глаза на наилучшее видение объекта, остается на том же уровне, что и у рыб: достигается только за счет перемещения хрусталика вперед и назад. Эти перемещения помогает совершать особая ресничная мышца, возникшая у земноводных.

Наземная среда меняет и характер передачи звуков: теперь он передается через колебания воздуха. Если рыбам в воде было достаточно внутреннего уха для улавливания колебаний воды, то земноводным этого мало. У них впервые возникает среднее ухо, которое снабжено барабанной перепонкой, чувствительной к колебаниям воздуха.
Среднее ухо состоит из одной слуховой косточки - стремени. Евстахиева труба соединяет полость среднего уха с с ротглоточной полостью, служит для уравнивания давления между ними.

Лягушки раздельнополые животные, оплодотворение наружное. Размножение и начальные этапы развития земноводных происходят в воде, поэтому амфибий крайне редко можно встретить в засушливых местах, где водоемы отсутствуют.
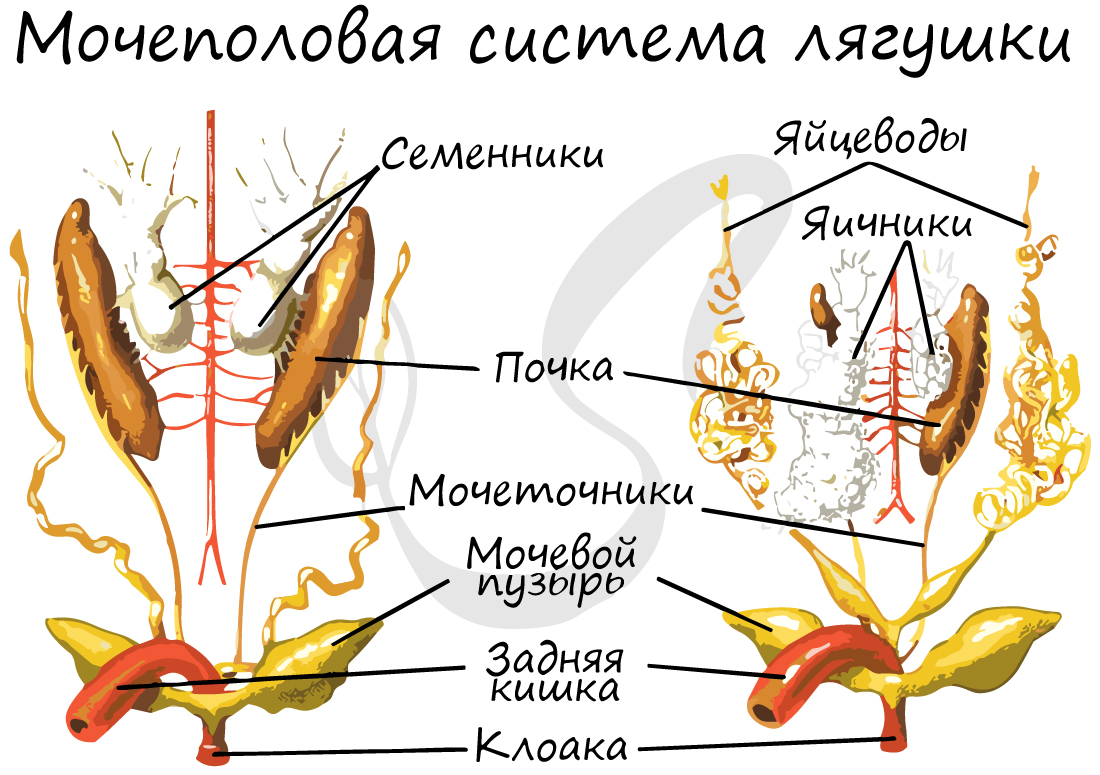
Мужские половые железы представлены парными семенниками с отходящими от них семявыносящими протоками, которые впадают в мочеточники, открывающиеся в клоаку. Женские половые железы - яичники с отходящими от них яйцеводами, открывающимися в клоаку.
Во время оплодотворения самец удерживает самку сзади и надавливает передними конечностями на ее брюшко, что приводит к выметыванию икры. Эта икра сразу же оплодотворяется семенной жидкостью самца.
Развитие происходит с метаморфозом - значительной перестройкой большинства органов. Личиночная стадия лягушки еще раз подчеркивает эволюционные корни этого класса, а именно то, что земноводные произошли от рыб. Головастики требуют нашего особого внимания.
Головастик - личинка бесховстых земноводных. Развивается из оплодотворенной яйцеклетки, живет в воде.

Во многом строение головастика напоминает строение рыбы, что подтверждает родство классов земноводных и рыб. Головастик характеризуется следующими чертами:
- Дыхание осуществляется с помощью жабр
- Форма тела рыбообразная, изначально конечности отсутствуют
- Имеется хвостовой плавник
- Двухкамерное сердце, один круг кровообращение
- Присутствует орган боковой линии
По мере изучения вышеуказанных признаков становится очевидно, что предками земноводных являются кистеперые рыбы, обитавшие мелких континентальных водоемах. От кистеперых рыб произошли древние земноводные - стегоцефалы, появившиеся в конце девонского периода и вымершие вначале мезозоя.

Земноводные, как и все живые организмы, являются звеном в цепи питания (консументами). Они уничтожают многих кровососущих насекомых, а также насекомых, которые наносят вред культурным растениям.
Их яд используют для изготовления лекарств, так что не забудем отметить их медицинское значение. Лягушка - классический объект для лабораторных исследований, в таком качестве лягушки используются повсеместно, и благодаря им совершены тысячи открытий.

Данная статья написана Беллевичем Юрием Сергеевичем и является его интеллектуальной собственностью. Копирование, распространение (в том числе путем копирования на другие сайты и ресурсы в Интернете) или любое иное использование информации и объектов без предварительного согласия правообладателя преследуется по закону. Для получения материалов статьи и разрешения их использования, обратитесь, пожалуйста, к Беллевичу Юрию.
У млекопитающих скелет шеи образован 7 позвонками за небольшим исключением (у ленивца – 6-9, у ламантина – 6). Они делятся на типичные – схожие по строению друг с другом (по счету 3, 4, 5, 6), и нетипичные (1, 2, 7).
Характерным признаком типичных шейных позвонков (рис. 1) является наличие двуветвистых (раздвоенных) поперечнореберных отростков (4) и межпоперечных (поперечных) отверстий – foramen transversarium (5), - расположенных у их основания. У типичных шейных позвонков к поперечным отросткам прирастают зачатки ребер, поэтому эти отростки называются не только поперечными, но и поперечнореберными – processus costotransversarius .

Рис. 1. Типичный шейный позвонок лошади
1 – головка позвонка; 2 – ямка позвонка; 3 – остистый отросток; 4 – поперечнореберные отростки; 5 – межпоперечное отверстие; 6 – краниальные суставные отростки; 7 – каудальные суставные отростки;
Особенности:
У крупного рогатого скота типичные шейные позвонки имеют сравнительно короткие тела (позвонки почти кубовидны), головки полушаровидной формы, остистые отростки короткие, округлые, на концах утолщены, высота их постепенно увеличивается с 3 по 7, хорошо выражены вентральные гребни.
У свиньи позвонки короткие, дужки узкие, междужковые отверстия широкие (расстояние между дужками рядом лежащих позвонков), головки и ямки плоские, остистые отростки относительно хорошо развиты, вентральные гребни отсутствуют, у основания поперечнореберных отростков имеются дорсовентральные отверстия (боковые позвоночные отверстия – foramen vertebrale laterale .
У лошади тела позвонков длинные, головки полушаровидной формы, остистые отростки в виде шероховатых гребешков, вентральные гребни хорошо развиты (кроме 6-го позвонка).
У собаки тела позвонков относительно длинные, головки и ямки плоские, косо поставлены по отношению к телу. Остистый отросток на 3-м позвонке отсутствует, а на остальных высота их постепенно увеличивается в каудальном направлении.
7-й шейный позвонок (рис.2) отличие от типичных имеет короткий неветвящийся поперечнореберный отросток (1), без межпоперечного отверстия в нем. Остистый отросток развит сильнее, чем на типичных шейных позвонках. На каудальном конце тела расположены каудальные реберные ямки (3) для сочленения с головками первой пары ребер.
Особенности:
У крупного рогатого скота остистый отросток высокий и широкий, стоит отвесно, суставные отростки широки и расставлены друг от друга, головка и ямки рельефно выражены (полушаровидной формы).
У свиньи головка и ямка позвонка плоские. Есть боковые позвоночные отверстия, идущие в дорсовентральном направлении.
У лошади остистый отросток относительно слабо развит, головка и ямка хорошо выражены, полушаровидной формы.
У собаки остистый отросток шиловидной формы, головка и ямка плоские, поставлены косо по отношению к телу.

Рис. 2 . Седьмой шейный позвонок лошади
1 – поперечнореберные отростки; 2 – остистый отросток; 3 – каудальные реберные ямки; 4 – краниальные суставные отростки; 5 – каудальные суставные отростки;
Первый шейный позвонок – или атлант – atlas (рис.3) – характерен отсутствием тела. Он имеет кольцевидную форму. На атланте различают дорсальную и вентральную дужки (дуги) - arcus dorsalis et ventralis с дорсальными и вентральными бугорками - tuberculum dorsale (1) et ventrale (2). Вентральная дуга заменяет тело атланта. Она со стороны позвоночного отверстия несет фасетку (ямку) для зубовидного отростка 2-го шейного позвонка - fovea dentis (3). Сбоку атланта располагаются крылья - ala atlantis (4), представляющие собой видоизмененные поперечные и суставные отростки, слитые в боковую массу - massa lateralis . На вентральной поверхности крыльев находится крыловая ямка - fossa atlantis (5). На краниальном конце атланта имеются краниальные суставные ямки - fovea articularis cranialis s . atlantis (6) для соединения с мыщелками затылочной кости, а на каудальном – каудальные суставные ямки - fovea articularis caudalis (7) – для соединения со 2-м шейным позвонком. На переднем конце крыла атланта находится крыловое отверстие - foramen alare (8), соединяющееся желобом с межпозвоночным отверстием - foramen intervertebrale (9). На каудальном конце крыльев имеется поперечное отвестие - foramen transversarium (10).

Рис. 3 . Атлант лошади
А – дорсальная поверхность; Б – вентральная поверхность.
1 – дорсальный бугорок; 2 – вентральный бугорок; 3 – фасетка (ямка) для зубовидного отростка 2-го шейного позвонка; 4 – крылья атланта; 5 – крыловая ямка; 6 – краниальные суставные ямки; 7 – каудальные суставные ямки; 8 - крыловое отверстие; 9 – межпозвоночное отверстие; 10 – поперечное отверстие.
Особенности:
У крупного рогатого скота крылья массивные со слабо выраженной ямкой, лежат горизонтально, поперечное (межпоперечное) отверстие отсутствует.
У свиньи крылья узкие и толстые, крыловая ямка мелкая, поперечное отверстие располагается на каудальном крае атланта, имеет форму канала и открывается в крыловую ямку. Ямка для зубовидного отростка глубокая. Вентральный бугорок направлен каудально в виде отростка.
У лошади крылья атланта тонкие и загнуты вентрально, вследствие чего крыловые ямки глубокие. Поперечное отверстие расположено на дорсальной поверхности крыла. Из трех отверстий оно более крупное.
У собаки крылья атланта плоские, тонкие и длинные, вытянуты латеро-каудально, поставлены почти горизонтально. Дорсальная дуга широкая и без бугорка. Крыловое отверстие заменено вырезкой (11).

4. Первый шейный позвонок (атлант)
А – атлант крупного рогатого скота; Б – атлант свиньи; В – атлант собаки.
Второй шейный позвонок – осевой, или эпистрофей – axis s . epistropheus (рис.8) – самый длинный из семи. Характеризуется наличием, вместо головки – зубовидного отростка, или зуба - dens (1) , остистый отросток в виде гребня – crista (2), со слабыми неветвящимися поперечнореберными отростками (3) с поперечными отверстиями (4) в виде канала и краниальными межпоперечными отверстиями (5).

Рис. 5 . Второй шейный позвонок (эпистрофей)
А – эпистрофей лошади; Б – эпистрофей крупного рогатого скота; В – эпистрофей свиньи; Г – эпистрофей собаки.
Особенности:
У крупного рогатого скота зубовидный отросток имеет вид полого полуцилиндра, а гребень – вид квадратной пластинки с приподнятым каудальным краем.
У свиньи зубовидный отросток тупой, конусообразной формы, гребень высокий, задний край его приподнят дорсально, передний – скошен. Встречаются дорсовентральные отверстия (6).
У лошади зубовидный отросток полуконической формы с плоской дорсальной поверхностью и выпуклой – вентральной. Мощный гребень каудально раздваивается и срастается с каудальными суставными отростками. Хорошо выражен вентральный гребень.
У собаки зубовидный отросток длинный, цилиндрической формы. Гребень нависает над зубовидным отростком в виде клюва, а каудально сливается с каудальными суставными отростками. Краниальные межпозвоночные отверстия заменены вырезками.
Отличительные признаки типичного шейного позвонка

Особенности типичных шейных позвонков




Особенности не типичных шейных позвонков

Позвоночник млекопитающих делится на пять отделов: шейный, грудной, поясничный, крестцовый и хвостовой (рис. 1). Соприкасающиеся поверхности этих позвонков плоские (платицельный тип), между позвонками залегают межпозвоночные диски, или мениски. Верх ние дуги хорошо выражены; в грудной области эти дуги продолжаются каждая в довольно длинный верхний костистый отросток. У оснований дуг с каждой стороны расположены суставные отростки. Ребра имеют двойное причленение к позвонкам. В шейной области они совершенно рудиментарны и срастаются с телами позвонков и с нижними частями поперечных отростков, причем между редуцированным ребром и поперечным отростком образуется небольшое отверстие. В грудном отделе каждое ребро причленяется головкой между двумя соседними телами позвонков, а бугорком—к нижней части поперечного отростка.
Ребро слагается из двух отделов: позвоночного и грудного. Последний остается хрящевым в течение всей жизни (за исключением однопроходных и неполнозубых, у которых он окостеневает). Поперечные отростки отходят от боковых оснований верхних дуг и только в поясничной области они иногда сидят на телах позвонков.
Рис. 1 . Скелет кролика.
1 —нижняя челюсть; 2 —верхняя челюсть; 3 —скуловая дуга; 4 —верх глазницы; 5 —шейные; 6—грудные; 7—поясничные; 8 —крестцовые и 9 —хвостовые позвонки; 10 —подвздошная кость; 11 —седалищная кость; 12—лонно-седалищное отверстие; 13 —лонная или лобковая кость; 14— бедро; 15 —коленная чашка; 1 6—большая берцовая кость; 17 —малая берцовая кость; 18 —пяточная кость; 19 —кости предплюсны; 20—ребра; 21 —локтевая кость; 22 —плечевая кость; 23 —лучевая кость; 24 —лопатка; 25 —грудина; 26 —ключица.
В шейном отделе у огромного большинства млекопитающих насчитывают 7 позвонков (только у ламантина и у одного вида ленивца, Choloepus hoffmani, их 6, а у ленивцев рода Bradypus —от 8 до 10). Длина шеи зависит, таким образом, не от числа позвонков, а от длины тела каждого из них. У китов шея почти совершенно отсутствует и слагается из 7 совершенно плоских позвонков, которые к тому же нередко срастаются между собой; у жирафы шея поддерживается 7 очень вытянутыми позвонками.
В грудном отделе насчитывают от 12 до 15 позвонков, в наибольшем числе случаев их 13 (у броненосца рода Tatusia и у клюворылого кита Hyperoodon их 9, у ленивца Choloepus их 24).
Число поясничных позвонков вариирует от 2 до 9. В состав их поперечных отростков входят и рудиментарные ребра.
У большинства млекопитающих имеется 4 крестцовых позвонка (реже 3 —у хищных). В крестце утконоса только 2 позвонка (как у рептилий).
Количество хвостовых позвонков подвержено большой вариации: у обезьян-гиббонов и у некоторых летучих мышей лишь 3 позвонка, у длиннохвостого танрека (из насекомоядных)—47, у длиннохвостого ящера—49.
Пояс передних конечностей млекопитающих
Грудина млекопитающих слагается из нескольких отделов. Расширенный передний (praesternum, или manubrium) связан с первой парой ребер и ключицами (если таковые имеются); средний отдел (mesostemum) состоит из нескольких костных частей, связывающих между собой концы нескольких пар ребер. Наконец, в нижней части грудина заканчивается мечевидным отростком (xypbisternum) яйцеродных млекопитающих существует непарное окостенение, лежащее впереди грудины и имеющее форму буквы Т. К отросткам упомянутого окостечения прилегают ключицы. Это—кость хрящевого происхождения, формирующаяся из трех зачатков. Вследствие указанных данных ее невозможно сравнивать с episternum рептилий, как это делалось прежними авторами (рис. 2, Ер).

Форма и очертания лопатки крайне вариируют. Пожалуй, наиболее часто эта кость имеет вид вытянутого неправильного треугольника (например у кролика). Вдоль верхней части, ближе к наружному краю, тянется гре бень лопатки, заканчивающийся спереди акромиальным отростком (processus acromialis). На относительно суженном нижне-переднем отделе лопатки заметна вогнутая сочленовная поверхность для головки плечевой кости, а над этой вогнутостью возвышается искривленный коракоидный отросток (дериват бывшего коракоида). У яйцеродных млекопитающих имеются хорошо развитые столбчатые метакоракоиды, похожие на таковые у птиц (рис. 2, Со). По интересному наблюдению Брума, у детенышей сумчатых куниц и кускусов (из Marsupialia), обретающихся еще в сумке матери, обнаруживаются коракоиды, соединяющиеся с грудиной; позднее они редуцируются и сливаются с лопаткой, образуя упомянутые отростки.
Рис. 2. Плечевой пояс утконоса.
Cl—ключица; G —сочленовная ямка для головки плечевой кости; Со —коракоид (метакор акоид); Со — эпикоракоид; Ер —episternum; S t—грудина; S —лопатка.
Ключицы имеют вид тонких, слегка искривленных костей. Они отсутствуют у копытных и хоботных; среди сумчатых они рудиментарны только у бандику; у насекомоядных отсутствуют только у выдровой землеройки; у большинства грызунов имеются; рудиментарны или отсутствуют у хищных.
Сравнение грудного пояса птицы и млекопитающего. В заключение полезно сделать сравнение частей грудного пояса птицы и млекопитающего . В прилагаемой схеме на звания костей, соединенных у данных животных сочленениями, объединены простыми стрелками, сросшиеся кости—двойными стрелками

Строение передней конечности млекопитающих
Конечности характеризуются значительной длиной их основных отделов. Плечевая кость отличается разнообразием своей длины и толщины. Так, она вытянута и тонка у летучих мышей, сравнительно весьма коротка и расширена у землероев, например, у крота. Концевой отдел плечевой кости несет расширенный сустав (trochlea) для соединения с предплечьем, состоящим из локтевой и лучевой костей. У многих зверей эти кости приблизительно одинаковой толщины и длины. Они весьма укорочены у китообразных, например, у дельфинов, и крайне длинны и вытянуты у ленивцев. Лучевая кость иногда снабжена сильным угловым отростком (processus olecranon); он особенно велик у крота и утконоса. У летучих мышей лучевая кость значительно больше и массивнее локтевой, при этом лучевая кость длиннее плечевой. Очень слаба локтевая кость у лошадей и жвачных и хорошо развита у свиней, бегемотов, тапиров и носорогов. В передней конечности (например, у человека) развивается способность лучевой кости вместе с кистью вращаться вокруг локтевой, так что рука из положения ладонью вверх и вперед (при параллельном положении костей предплечья)— супинация—может переходить в расположение ладонью вниз и назад— пронация (с перекрестом локтевой кости лучевой).
Кисть передней конечности относительно примитивна; обычно имеются все пять пальцев, нередки рудименты переднего шестого (praepollex). Присутствие центральной косточки запястья может считаться примитивным признаком. Она имеется у грызунов, китообразных и многих неполнозубых; отсутствует или слабо развита у многих копытных (за исключением даманов, тапиров, свиней), имеется у молодых слонов, у которых позднее сливается с radiale.

Здесь дана новая сравнительно-анатомическая номенклатура названных костей. Для сопоставления с анатомией человека и со старыми руководствами привожу прежние наименования костей:
Carpale 1 ……..trapezium, multangulum majus
Garpale 3 ……..trapezoideum, multangulum minus
Carpale 3+4 ……hamatum, uncinatum
Сообразно с разнообразием функций структура конечностей весьма различна. У роющих млекопитающих (кротов, броненосцев) лапы широки, у плавающих (киты, сирены)—ластообразны. У летучих мышей чрезвычайно удли няются пальцы (особенно вытянуты пястные кости). Большинство млекопитающих при ходьбе упирается на всю кисть и ступню (стопоходящие; быстро бегающие); многие грызуны, хищники становятся лишь на концы пальцев (пальцеходящие). У лучших бегунов среди копытных животное ступает либо на концы средних пальцев (третьего и четвертого у парнокопытных), либо на один третий палец (у непарнокопытных).
Строение таза млекопитающих
Таз слагается из трех костей: подвздошной (ileum), седалищной (ischium) и лобковой (pubis). Кроме них, нередко развивается еще осо бое окостенение—небольшая вертлужная кость (os acetabulare), лежащая в передней части вертлужной впадины. Обычно у взрослых животных все названные кости срастаются между собой в одну безыменную кость (os innominatum). У млекопитающих наблюдается широкое сращение, соединяющее лобковый и седалищный отделы (у большинства насекомоядных, у броненосцев, приматов названное сращение имеется лишь в передней, лонной, части тазового пояса).
У однопроходных и сумчатых с лобковыми костями связана пара косточек (ossa marsupialia). к поддержанию сумки эти кости не имеют прямого отношения. Вероятно, они служат в качестве пассивного упора для мускулатуры при давлении мускулов на молочные соски при лактации (М. Weber, 1928).
Структура задней конечности
Основной отдел бедра несет два-три выступа для прикрепления мышц. В голени малая берцовая кость развита слабее большой, иногда совершенно редуцируется и прирастает к последней. В колене наблюдается сухожильная кость коленная чашечка (patella). В предплюсне расположены лишь две кости: наружная пяточная (fibulare -calcaneus) несет большой пяточный выступ; внутренняя таранная кость (astragalus) представляет обычно комплекс из слияния промежуточной (intermedium), центральной (centrale) костей, а также редуцированной tibiale. Общая схема задней ступни такова:

Статья на тему Строение позвоночника млекопитающих
Читайте также: