Кортиев орган эмбриона. Развитие кортиева органа плода
Добавил пользователь Алексей Ф. Обновлено: 28.01.2026
Внутреннее ухо состоит из костного лабиринта и включенного в него перепончатого лабиринта.
Лабиринт подразделяется на 3 отдела - средний - преддверие, кзади от него - система полукружных каналов, впереди от преддверия - улитка.
Перепончатая улитка - спиралевидный канал с расположенным внутри него рецепторным аппаратом - спиральным (или кортиевым органом). На поперечном разрезе улитковый ход имеет треугольную форму; образован вестибулярной стенкой (обращенной к лестнице преддверия; представляет собой очень тонкую мембрану Reissneri); наружная стенка образована спиральной связкой с расположенными на ней эпителиальными клетками сосудистой полоски; нижняя тимпанальная стенка обращена к барабанной лестнице и представлена основной мембраной , на которой лежит спиральный орган - периферический рецептор кохлеарного нерва.
Спиральный (кортиев) орган состоит из нейроэпителиальных наружных и внутренних волосковых клеток, поддерживающих клеток (Дейтерса, Гензена, Клаудиуса), наружных и внутренних столбиковых клеток, образующих кортиевы дуги. Внутренние и наружные столбиковые клетки образуют кортиев тоннель. Кнутри от внутренних столбиковых клеток располагается ряд внутренних волосковых клеток (количеством до 3500); снаружи от наружных столбиковых клеток - 4 ряда наружных волосковых клеток (по 5000 в каждом ряду), поддерживаемых клетками Дейтерса, Гензена и Клаудиуса. Волосковые клетки охватываются нервными волокнами, исходящими из биполярных клеток спирального ганглия. Между клетками кортиева органа имеются внутриэпителиальные пространства, заполненные жидкостью (кортилимфой). Она имеет связь с перелимфой и довольно близка к ней по химическому составу. Считается, что основная функция кортилимфы - трофическая (т.к. кортиев орган не имеет собственной васкуляризации).
Над кортиевым органом расположена покровная мембрана - мягкая, упругая пластинка, состоящая из протофибрилл, имеющих продольное и радиальное направление. Эластичность этой мембраны различна в продольном и поперечном направлениях. В покровную мембрану проникают волоски нейроэпителиальных волосковых клеток. При колебаниях основной мембраны происходит напряжение и сжатие этих волосков, что служит моментом трансформации механической энергии в энергию электрического нервного импульса.
Периферический рецептор слухового анализатора - спиральный орган. Сюда (к волосковым клеткам) подходят дендриты клеток спирального ганглия, расположенного в улитке лабиринта. Аксоны проникают в полость черепа через внутреннее слуховое отверстие, входят в ствол мозга в мостомозжечковом углу и заканчиваются в вентральном и дорсальном ядрах моста - вторые нейроны слухового пути. От оливы начинается третий нейрон. Подкорковые слуховые центра - в задних бугорках четверохолмия и медиальном коленчатом теле. Корковый коней слухового анализатора - в заднем отделе верхней височной извилине и извилине Гешля.
Резонансная теория Геймгольца.
В XIX веке, когда господствующим в медицине было морфологическое направление, в качестве основного критерия, определяющего звуковосприятие, бралась определенная деталь строения слухового органа. Основная мембрана, на которой расположен спиральный орган, при осмотре ее с помощью увеличительной оптики имеет поперечную исчерченность, как бы состоит из «струн» разной длины. Исходя из этого факта, Гельмгольц в 1863 г. создал так называемую резонансную теорию слуха. Согласно этой теории, в улитке возникают явления механического резонанса в отношении звуковых колебаний различных частот. По аналогии со струнными инструментами звуки высокой частоты приводят в колебательное движение участок основной мембраны, с короткими волокнами у основания улитки, а звуки низкой частоты — в колебательное движение участок мембраны с длинными волокнами у верхушки улитки. При подаче и восприятии сложных звуков одновременно начинает колебаться несколько участков мембраны. Чувствительные клетки спирального органа воспринимают эти колебания и передают по нерву слуховым центрам. На основании изучения теории Гельмгольца можно сделать три вывода: 1) улитка является тем звеном слухового анализатора, где возникает первичный анализ звуков; 2) каждому простому звуку присущ определенный участок на основной мембране; 3) низкие звуки приводят в колебательное движение участки основной мембраны, расположенные у верхушки улитки, а высокие — у ее основания.
Таким образом, теория Гельмгольца впервые позволила объяснить основные свойства уха, т. е. определение высоты, силы и тембра. В свое время эта теория нашла много сторонников и до сих пор считается классической. Действительно, вывод Гельмгольца о том, что в улитке происходит первичный пространственный анализ звуков, полностью соответствует теории И. П. Павлова о способности к первичному анализу как концевых приборов афферентных нервов, так и в особенности сложных рецепторных аппаратов.
Резонансная теория Гельмгольца получила подтверждение и в клинике. Гистологическое исследование улиток умерших людей, страдавших островковыми выпадениями слуха, позволило обнаружить изменения кортиева органа в участках, соответствующих утраченной части слуха. Вместе с тем современные знания не подтверждают возможность резонирования отдельных «струн» основной мембраны; следовательно, необходимы более точные объяснения пространственной рецепции звуков в улитке.
2. Травмы и инородные тела полости носа, ринолиты. Диагностика. Методы удаления инородных тел. Осложнения.
Тема 29. СТРОЕНИЕ ОРГАНА СЛУХА И РАВНОВЕСИЯ
У 22-х дневного эмбриона на уровне ромбовидного мозга появляются парные утолщения эктодермы - слуховые плакоды. Путем инвагинации и последующего отделения от эктодермы формируется слуховой пузырек. С медиальной стороны к слуховому пузырьку прилежит зачаток слухового ганглия, из которого впоследствии дифференцируется ганглий преддверия и ганглий улитки. По мере развития в слуховом пузырьке появляются две части - эллипсовидный мешочек (утрикулюс с полукружными каналами) и сферический мешочек (саккулюс) с зачатком канала улитки.
Строение органа слуха
Наружное ухо включает в себя ушную раковину, наружный слуховой проход и барабанную перепонку, передающую звуковые колебания на слуховые косточки среднего уха. Ушная раковина образована эластическим хрящом, покрытым тонкой кожей. Наружный слуховой проход выстлан кожей, содержащей волосяные фолликулы, типичные сальные железы и церуминозные железы - видоизмененные сальные железы, вырабатывающие ушную серу. Наружная поверхность барабанной перепонки покрыта кожей. Изнутри, со стороны барабанной полости, барабанная перепонка выстлана однослойным кубическим эпителием, который отделен от наружного слоя тонкой соединительно-тканной пластинкой.
Среднее ухо содержит слуховые косточки - молоточек, наковаленку и стремечко, которые передают колебания с барабанной перепонки на мембрану овального окна. Барабанная полость выстлана многослойным эпителием, который переходит в однослойный цилиндрический мерцательный у отверстия слуховой трубы. Между эпителием и костью располагается прослойка плотной волокнистой соединительной ткани. Кость медиальной стенки барабанной полости имеет два окна - овальное и круглое, которые отделяют барабанную полость от костного лабиринта внутреннего уха.
Внутреннее ухо образовано костным лабиринтом височной кости, который содержит повторяющий его рельеф перепончатый лабиринт. Костный лабиринт - система полукружных каналов и сообщающаяся с ними полость-преддверие. Перепончатый лабиринт - система тонкостенных соединительно-тканных трубок и мешочков, расположенная внутри костного лабиринта. В костных ампулах перепончатые каналы расширяются. В преддверии перепончатый лабиринт образует два сообщающихся между собой мешочка: улюс (эллиптический мешочек), в который открываются перепончатые каналы и саккулюс (сферический мешочек). Перепончатые полукружные каналы и мешочки преддверия заполнены эндолимфой и сообщаются с улиткой, а также с расположенным в полости черепа эндолимфатическим мешком, где эндолимфа резорбируется. Эпителиальная выстилка эндолимфатического мешка содержит цилиндрические клетки с плотной цитоплазмой и ядрами неправильной формы, а также цилиндрические клетки со светлой цитоплазмой, высокими микроворсинками, многочисленными пиноцитозными пузырьками и вакуолями. В просвете мешка присутствуют макрофаги и нейтрофилы.
Строение улитки. Улитка - это спирально закрученный костный канал, развившийся как вырост преддверия. Улитка образует 2,5 завитка длиной около 35 мм. Базилярная (основная) и вестибулярная мембраны, расположенные внутри канала улитки, делят его полость на три части: барабанную лестницу, вестибулярную лестницу и перепончатый канал улитки, (среднюю лестницу или улитковый ход). Эндолимфа заполняет перепончатый канал улитки, а перилимфа - вестибулярную и барабанную лестницы. Барабанная лестница и вестибулярная лестница сообщаются у вершины улитки с помощью отверстия (геликотремы). В перепончатом канале улитки на базилярной лестнице расположен рецепторный аппарат - спиральный (или кортиев) орган.
Концентрация К+ в эндолимфе в 100 раз больше, чем в перилимфе; концентрация Na+ в эндолимфе в 10 раз меньше, чем в перилимфе.
Перилимфа по химическому составу близка к плазме крови и си жидкости и занимает промежуточное положение между ними по содержанию белка.
Строение кортиевого органа. Кортиев орган содержит несколько рядов волосковых клеток, связанных с текториальной (покровной мембраной). Различают внутренние и наружные волосковые и поддерживающие клетки.
Волосковые клетки - рецепторные, образуют синаптические контакты с периферическими отростками чувствительных нейронов спирального ганглия. Внутренние волосковые клетки образуют один ряд, имеют расширенное основание, 30 - 60 неподвижных микроворсинок (стереоцилий), проходящих через кутикулу в апикальной части. Стереоцилии расположены полукругом, открытым в сторону наружных структур кортиева органа. Внутренние волосковые клетки - первичные сенсорные клетки, которые возбуждаются в ответ на звуковой раздражитель и передают возбуждение афферентным волокнам слухового нерва. Смещение покровной мембраны вызывает деформацию стереоцилий, в мембране которых открываются механочувствительные ионные каналы и возникает деполяризация. В свою очередь, деполяризация способствует открытию потенциалочувствительных Са 2 + и К+-каналов, встроенных в базолатеральную мембрану волосковой клетки. Возникающее повышение в цитозоле концентрации Са 2 + инициирует секрецию (наиболее вероятен глютамат) из синаптических пузырьков с последующим его воздействием на постсинаптическую мембрану в составе афферентных терминалей слухового нерва.
Наружные волосковые клетки расположены в 3 - 5 рядов, имеют цилиндрическую форму и стереоцилии. Миозин распределяется вдоль стереоцилии волокнистой клетки.
Поддерживающие клетки. Среди поддерживающих клеток различают внутренние фаланговые клетки, внутренние клетки-столбы, наружные фаланговые клетки Дейтерса, наружные клетки-столбы, клетки Гензена, клетки Беттхера. Фаланговые клетки вступают в контакт с волосковыми на базальной мембране. Отростки наружных фаланговых клеток проходят параллельно наружным волосковым клеткам, не соприкасаясь с ними на значительном протяжении, и на уровне апикальной части волосковых клеток вступают с ними в контакт. Поддерживающие клетки связаны щелевыми контактами, образованными мембранным белком щелевого контакта коннексином-26. Щелевидные контакты участвуют в восстановлении уровня К+ в эндолимфе в ходе следовых реакций после возбуждения волосковых клеток.
Путь передачи слухового раздражения
Цепочка передачи звукового давления выглядит следующим образом: барабанная перепонка далее слуховые косточки - молоточек, наковаленка, стремечко, далее - мембрана овального окна, перилимфа базилярная и текториальная мембраны и мембрана круглого окна.
При смещении стремечка частицы перелимфы перемещаются по вестибулярной лестнице и затем через геликотрему по барабанной лестнице - к круглому окну.
Жидкость, сдвинутая смещением мембраны овального окна, создает избыточное давление в вестибулярном канале. Под действием этого давления базальный участок основной мембраны смешается в сторону барабанной лестницы. Колебательная реакция в виде волны распространяется от базальной части основной мембраны к геликотреме. Смещение текториальной мембраны относительно волосковых клеток при действии звука вызывает их возбуждение. Смещение мембраны относительно сенсорного эпителия отклоняет стереоцилии волосковых клеток, что открывает механочувствительные каналы в клеточной мембране и приводит к деполяризации клеток. Возникающая электрическая реакция, названная микрофонным эффектом, по своей форме повторяет форму звукового сигнала.
Строение и функционирование органа равновесия
В ампулярном расширении полукружного канала находятся кристы (или гребешки). Чувствительные области в мешочках называются пятнами.
В состав эпителия пятен и крист входят чувствительные волосковые и поддерживающие клетки. В эпителии пятен киноцилии распределяются особым образом. Здесь волосковые клетки образуют группы из нескольких сот единиц. Внутри каждой группы киноцилии ориентированы одинаково, однако ориентация самих групп различна. Эпителий пятен покрыт отолитовой мембраной. Отолиты - кристаллы карбоната кальция. Эпителий крист окружен желатинообразным прозрачным куполом.
Волосковые клетки присутствуют в каждой ампуле полукружных каналов и в пятнах мешочков преддверия. Различают два типа волосковых клеток. Клетки типа I обычно расположены в центре гребешков, а клетки типа II - по периферии. Клетки обоих типов в апикальной части содержат 40 - 110 неподвижных волосков (стереоцилий) и одну ресничку (киноцилию), расположенную на периферии пучка стереоцилий. Самые длинные стереоцилии находятся вблизи киноцилии, а длина остальных уменьшается по мере удаления от киноцилии.
Волосковые клетки чувствительны к направлению действия стимула (дирекционная чувствительность). При направлении раздражающего воздействия от стереоцилии к киноцилии волосковая клетка возбуждается. При противоположном направлении стимула происходит угнетение ответа. Клетки типа I имеют форму амфоры с закругленным дном и размещены в бокалообразной полости афферентного нервного окончания. Эфферентные волокна образуют синаптические окончания на афферентных волокнах, связанных с клетками I типа. Клетки типа II имеют вид цилиндров с округлым основанием. Характерная особенность этих клеток заключается в их иннервации: нервные окончания здесь могут быть как афферентными (большинство), так и эфферентными.
При сверхпороговом звуковом раздражении (акустическая травме) и при действии некоторых ототоксических препаратов (антибиотиков стрептомицина, гентамицина) волосковые клетки погибают. Возможность их регенерации из клеток-предшественниц нейросенсорного эпителия имеет важное практическое значение, считается установленным для птиц и интенсивно изучается на млекопитающих.
Вестибулярный нерв образован отростками биполярных нейронов в составе вестибулярного ганглия. Периферические отростки этих нейронов подходят к волосковым клеткам каждого полукружного канала, утрикулюса и саккулюса, а центральные направляются в вестибулярные ядра продолговатого мозга.
Данный текст является ознакомительным фрагментом.
Продолжение на ЛитРес
6. СКЕЛЕТ СВОБОДНОЙ ВЕРХНЕЙ КОНЕЧНОСТИ. СТРОЕНИЕ ПЛЕЧЕВОЙ КОСТИ И КОСТЕЙ ПРЕДПЛЕЧЬЯ. СТРОЕНИЕ КОСТЕЙ КИСТИ
6. СКЕЛЕТ СВОБОДНОЙ ВЕРХНЕЙ КОНЕЧНОСТИ. СТРОЕНИЕ ПЛЕЧЕВОЙ КОСТИ И КОСТЕЙ ПРЕДПЛЕЧЬЯ. СТРОЕНИЕ КОСТЕЙ КИСТИ Плечевая кость (humerus) имеет тело (центральную часть) и два конца. Верхний конец переходит в головку (capet humeri), по краю которой проходит анатомическая шейка (collum anatomikum).
8. СТРОЕНИЕ СКЕЛЕТА СВОБОДНОЙ ЧАСТИ НИЖНЕЙ КОНЕЧНОСТИ. СТРОЕНИЕ БЕДРЕННОЙ КОСТИ, НАДКОЛЕННИКА И КОСТЕЙ ГОЛЕНИ. СТРОЕНИЕ КОСТЕЙ СТОПЫ
8. СТРОЕНИЕ СКЕЛЕТА СВОБОДНОЙ ЧАСТИ НИЖНЕЙ КОНЕЧНОСТИ. СТРОЕНИЕ БЕДРЕННОЙ КОСТИ, НАДКОЛЕННИКА И КОСТЕЙ ГОЛЕНИ. СТРОЕНИЕ КОСТЕЙ СТОПЫ Бедренная кость (os femoris) имеет тело и два конца. Проксимальный конец переходит в головку (caput ossis femoris), посередине которой расположена
2. СТРОЕНИЕ ПОЛОСТИ РТА. СТРОЕНИЕ ЗУБОВ
2. СТРОЕНИЕ ПОЛОСТИ РТА. СТРОЕНИЕ ЗУБОВ Полость рта (cavitas oris) при сомкнутых челюстях заполнена языком. Ее наружными стенками является язычная поверхность зубных дуг и десен (верхних и нижних), верхняя стенка представлена небом, нижняя - мышцами верхней части шеи, которые
5. Боевые повреждения органа зрения
5. Боевые повреждения органа зрения Боевые повреждения органа зрения имеют ряд существенных особенностей по сравнению с травмами мирного времени. В отличие от бытовых травм все боевые повреждения являются огнестрельными. Чаще ранения глаза наносятся осколками, пулевые
ЭВОЛЮЦИЯ ОРГАНА ЗРЕНИЯ
ЭВОЛЮЦИЯ ОРГАНА ЗРЕНИЯ В ходе филогенетического развития организмов под влиянием условий внешней среды орган зрения претерпел большие изменения. Из примитивного органа зрения, который состоит из светочувствительных клеток, располагающихся в наружных покровах
ГЛАВА 8 ОЖОГИ ОРГАНА ЗРЕНИЯ
ГЛАВА 8 ОЖОГИ ОРГАНА ЗРЕНИЯ Ожоги глаза и его придаточного аппарата могут быть вызваны химическими, термическими и лучевыми факторами. Тяжесть поражения зависит от свойств повреждающего вещества, длительности его воздействия, своевременности и качества оказания
О естестве органа и его частей
О естестве органа и его частей Мы говорим: органы суть тела, рождающиеся из первого смешения достохвальных соков, так же, как соки суть тела, рождающиеся из первого смешения элементов.Среди органов есть органы простые и есть сложные. Простые органы суть те, любая ощутимая
Глава 5. ТРАВМЫ ОРГАНА ЗРЕНИЯ
Глава 5. ТРАВМЫ ОРГАНА ЗРЕНИЯ ВИДЫ ПОВРЕЖДЕНИЙ ГЛАЗ Травмы органа зрения встречаются довольно часто. На их долю приходится 5-10 % всех болезней глаз.Травмы глаз являются одной из основных причин слепоты и инвалидности. По различным данным, примерно в 60-86 % случаев
ОЖОГИ ОРГАНА ЗРЕНИЯ
ОЖОГИ ОРГАНА ЗРЕНИЯ Ожоги глаза и его придаточного аппарата могут быть вызваны следующими факторами:— химическими;— термическими;— лучевыми.Тяжесть поражения зависит от свойств повреждающего вещества, длительности его воздействия, своевременности и качества
Глава 7. ПЕСНЯ БОЛЬНОГО ОРГАНА
Глава 7. ПЕСНЯ БОЛЬНОГО ОРГАНА Пока я писал эту книгу о геморрое, я усвоил для себя одну очень важную вещь: самое главное для каждого человека, желающего быть здоровым, — это умение управлять своим телом, не позволяя собственным органам бездействовать и болеть (что, в
ПОВРЕЖДЕНИЯ ОРГАНА ЗРЕНИЯ
ПОВРЕЖДЕНИЯ ОРГАНА ЗРЕНИЯ Глаза в силу своего поверхностного расположения чрезвычайно уязвимы, а тяжесть их поражения во многом определяется потребностью их постоянного функционирования. Если у пострадавшего имеется большое количество различных повреждений, но ни
Травмы органа зрения
Травмы органа зрения Приблизительно 10 % всех болезней в глазу происходят из-за травм. Результатом их может стать и полная слепота, и инвалидность. От 60 до 80 % случаев повреждений глаз происходят на производстве. Мужчины чаще наносят себе травмы, чем женщины. Как правило,
Тренировка органа спокойствия
Тренировка органа спокойствия Шестьдесят процентов населения России находятся на грани нервного срыва. Это означает, что имеется острая нехватка спокойствия. А врачи говорят, что примерно 60 % всех болезней возникает на нервной почве. Обратите внимание: и там и тут - 60 %.Я
Неоселен и заболевании органа зрения
Неоселен и заболевании органа зрения Катаракта, макулодистрофия, атрофия зрительного нерва. Прогрессирование этих заболеваний может остановить прием селена. На остроту зрения влияние небольшое, но у некоторых людей отмечался значительный положительный результат.
Анализаторы слуха и равновесия (орган слуха и равновесия, ухо)
Анализаторы слуха и равновесия (орган слуха и равновесия, ухо) Во все времена ухо не менее почитаемо, чем глаз. И даже больше. Ведь маленький принц Антуана де Сент-Экзюпери уверен, что самое главное невидимо для глаз. А король Лир говорит ослепленному Глостеру: «Чтоб видеть
Кортиев орган
Кортиев орган — рецепторная часть слухового анализатора, расположенная внутри перепончатого лабиринта. В процессе эволюции возникает на основе структур органов боковой линии.
Воспринимает колебания волокон, расположенных в канале внутреннего уха, и передаёт в слуховую зону коры больших полушарий, где и формируются звуковые сигналы. В кортиевом органе начинается первичное формирование анализа звуковых сигналов.
Содержание
История изучения
Открыт итальянским гистологом Альфонсо Корти (Alfonso Corti; 1822—1876).
Анатомия
Расположение
Кортиев орган располагается в спирально завитом костном канале внутреннего уха — улитковом ходе, заполненном эндолимфой и перилимфой. Верхняя стенка хода прилегает к т. н. лестнице преддверия и называется рейснеровой перепонкой; нижняя стенка, граничащая с т. н. барабанной лестницей, образована основной перепонкой, прикрепляющейся к спиральной костной пластинке.
Структура и функции
К. о. расположен на основной перепонке и состоит из внутренних и наружных волосковых клеток, внутренних и наружных опорных клеток (столбовых, клеток Дейтерса, Клаудиуса, Гензена), между которыми находится туннель, где проходят направляющиеся к основаниям волосковых клеток отростки нервных клеток, лежащих в спиральном нервном ганглии. Воспринимающие звук волосковые клетки располагаются в нишах, образуемых телами опорных клеток, и имеют на поверхности, обращенной к покровной перепонке, по 30—60 коротких волосков. Опорные клетки выполняют также трофическую функцию, направляя поток питательных веществ к волосковым клеткам.
Функция Кортиева органа — преобразование энергии звуковых колебаний в процесс нервного возбуждения.
Физиология
Звуковые колебания воспринимаются барабанной перепонкой и через систему косточек среднего уха передаются жидким средам внутреннего уха — перилимфе и эндолимфе. Колебания последних приводят к изменению взаиморасположения волосковых клеток и покровной перепонки Кортиева органа, что вызывает сгибание волосков и возникновение биоэлектрических потенциалов, улавливаемых и передаваемых в центральную нервную систему отростками нейронов спирального ганглия, подходящими к основанию каждой волосковой клетки.
По другим представлениям, волоски звуковоспринимающих клеток — лишь чувствительные антенны, деполяризующиеся под действием приходящих волн за счёт перераспределения ацетилхолина эндолимфы. Деполяризация вызывает цепь химических превращений в цитоплазме волосковых клеток и возникновение нервного импульса в контактирующих с ними нервных окончаниях. Различающиеся по высоте звуковые колебания воспринимаются различными отделами Кортиевого органа: высокие частоты вызывают колебания в нижних отделах улитки, низкие — в верхних, что связано с особенностями гидродинамических явлений в ходе улитки.
Добавьте ссылки на источники, в противном случае она может быть выставлена на удаление.
Дополнительные сведения могут быть на странице обсуждения.
Таким образом улитка является механическим измерителем АЧХ, и по действию схожа с АЧХ-метром, а не с микрофоном. Это позволяет мозгу сразу реагировать на конкретный звук, а не производить преобразование Фурье математически (на что, впрочем, у него не хватит вычислительных способностей), с целью разложения воспринимаемого звука на отдельные источники.
По поляризации звуковых гармоник можно судить о направлении(угловом) источника звука. Таким образом ухо позволяет получить информацию о амплитуде и поляризации каждой гармоники звуковых колебаний. Для низких частот (десятки герц) ухо и мозг успевают также извлечь информацию о фазе гармоник, что позволяет определить направление (как расстояние от головы по оси, проходящей через уши) низкочастотного колебания, если вычислить разность фаз сигнала от правого и левого уха.
Особенность дополнительного сжатия акустической информации позволяет значительно сократить время на анализ полученных данных. Закрученность улитки позволяет снимать спектр, совмещая октавы, то есть ось частоты в АЧХ звуковых колебаний закручивается, амплитуды октав совмещаются, что даёт возможность значительно сократить количество необходимых информационных каналов. Эта физическая основа слуха служит причиной восприятия музыки человеком.
Кортиев орган [organum spirale (PNA), organon spirale (JNA), organon spirale (Cortii) (BNA); по имени итал. анатома и гистолога A. Корти; син. спиральный орган] — периферический, или рецепторный, аппарат слухового анализатора. Слуховой рецептор представляет собой совокупность волосковых (сенсорно-эпителиальных) клеток, расположенных на базилярной пластинке улиткового протока, которые осуществляют преобразование звукового раздражения в физиологический акт слуховой рецепции. Физиологическая активность Кортиевого органа зависит от колебательных процессов в прилежащих мембранах и окружающих жидкостях, а также от метаболизма всего комплекса тканей улитки, в особенности сосудистой полоски.
История
Электронно-микроскопически строение Кортиевого органа было изучено Энгстремом (H. Engstrom, 1960) и Спондлином (Spoendlin, 1966). Раух (S. Rauch, 1964) опубликовал результаты исследований по биохимическому составу жидкостей и тканей внутреннего уха.
Сравнительная анатомия и эмбриология
У низших позвоночных не имеется дифференцировки нейроэпителия внутреннего уха на слуховой и вестибулярный. Механизм слухового восприятия у рыб, земноводных и пресмыкающихся иной. Гомолог органа слуха млекопитающих появляется у птиц, однако в нем нет еще разделения волосковых (сенсорно-эпителиальных) клеток на внутренние и наружные.
Зачаток внутреннего уха у человека появляется на ранних стадиях эмбриональной жизни и имеет эктодермальное происхождение. Проходя отдельные стадии развития (слуховая плакода, слуховой пузырек), внутреннее ухо разделяется на 2 мешочка. Из нижнего мешочка формируется улитковый проток, спиральный рост к-рого у эмбрионов длиной 40—50 мм приводит к образованию 2,5 завитков. Общий эпителиальный зачаток рецепторов внутреннего уха образуется у эмбрионов длиной 5 мм. Кубический эпителий зачатка вступает в связь с ганглиозными клетками. Разделение слухового пузырька на мешочки приводит также и к разделению эпителиального зачатка. Дифференцировка эпителия на волосковые (сенсорно-эпителиальные и поддерживающие) клетки завершается у эмбрионов длиной 70 мм. Полного развития К. о. достигает примерно через 2 нед. после рождения ребенка. С 40—45-летнего возраста наблюдается постепенная возрастная дистрофия К. о., сопровождающаяся понижением слуховой чувствительности к дискантовым звукам.
Анатомия и гистология
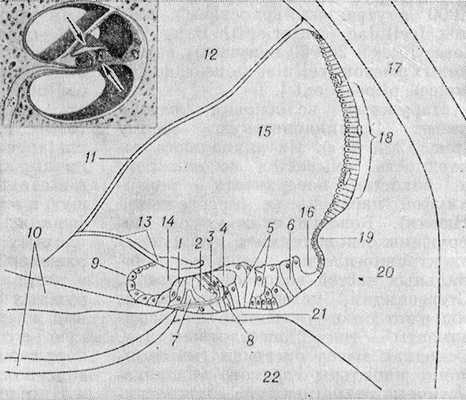
Рис. 1. Схема строения кортиева органа (поперечный срез): 1— внутренние волосковые клетки; 2— паратуннель (пространство Нюэля); 3 — наружные волосковые клетки; 4— наружный туннель; 5— наружные пограничные клетки (клетки Гензена); 6— наружные поддерживающие клетки (клетки Клаудиуса); 7— туннель наружный (кортиев туннель); 8 — наружные фаланговые клетки (клетки Дейтерса); 9— лимб костной спиральной пластинки; 10— костная спиральная пластинка; 11—пред-дверная мембрана (вестибулярная мембрана); 12— лестница преддверия; 13 — покровная мембрана; 14 — внутренняя спиральная борозда; 15 — улитковый канал; 16—наружная спиральная борозда; 17— улитка; 18—сосудистая полоска; 19—спиральный выступ; 20— спиральная связка улитки; 21—базилярная пластинка; 22— барабанная лестница. Слева вверху схема поперечного среза улитки, расположение кортиева органа указано стрелками.
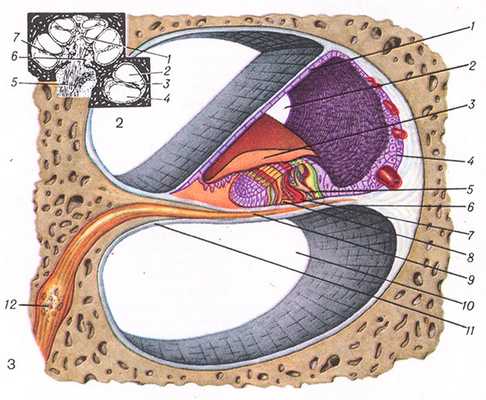
Рис. 2. Разрез улитки: 1 — ganglion spirale cochleae; 2 — scala vestibuli; 3 — ductus cochlearis; 4 — scala tympani; 5 —pars cochlearis n. vestibulocochlearis; 6 — modiolus; 7 — organon spirale. Рис. 3. Поперечное сечение витка улитки: 1 — membrana vestibularis; 2 — ductus cochlearis; 3 — membrana tectoria; 4 — stria vascularis; 5 —cellulae phalangae ext.; 6 — membrana basilaris с расположенным на ней спиральным органом; 7 — cellula pilaris ext.; 8 —cellula pilaris int.; 9 — pars cochlearis n. vestibulocochlearis; 10 — scala tympani; 11 — lamina spiralis secundaria; 12 — ganglion spirale.
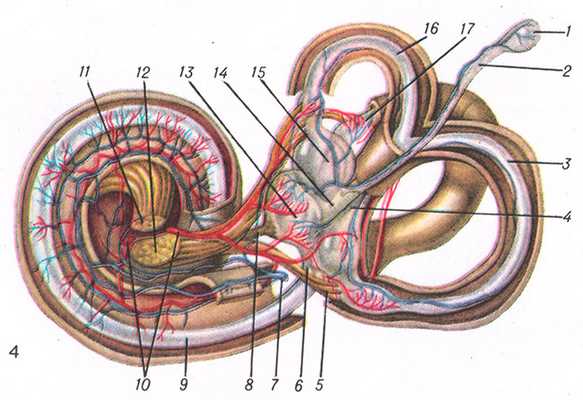
Рис. 4. Правый костный лабиринт (вскрыт) и заключающийся в нем перепончатый лабиринт: 1 — saccus endolymphaticus; 2 —ductus endolymphaticus; 3 — ductus semicircularis post.; 4 — crus membranaceum commune; 5 — n. ampullaris post.; 6 — ductus reuniens; 7 — v. labyrinthi; 8 — n. saccularls; 9 — ductus cochlearis; 10 — a. labyrinthi; 11 — pars cochlearis n. vestibulocochlearis; 12 — pars vestibularis n. acustici; 13 — sacculus; 14 — ductus utriculosaccularis; 15 — utriculus; 16 — ductus semicircularis ant.; 17 — dyctus semicircularis lat.
Улитка (cochlea) представляет собой передний отдел внутреннего уха (см.). Завитки улитки обвивают костный стержень (modiolus), несущий в себе сосуды и нервы (цветн. рис. 2—4). На поперечном срезе (рис. 1) в каждом завитке различают два перилимфатических канала — лестницу преддверия (scala vestibuli), находящуюся выше преддверной (рейсснеровой) мембраны (membrana vestibularis), и барабанную лестницу (scala tympani), расположенную ниже базилярной пластинки (lamina basilaris). Обе лестницы соединены у верхушки улитки отверстием улитки (helicotrema). Эндолимфатическое пространство в пределах улитки — улитковый проток (ductus cochlearis) — ограничено снизу базилярной пластинкой, сверху преддверной мембраной и латерально сосудистой полоской (stria vascularis), спиральным выступом (prominentia spiralis) и наружной спиральной бороздкой (sulcus spiralis ext.).
Кортиев орган занимает большую часть эндолимфатической поверхности базилярной пластинки, тянущейся между костной спиральной пластинкой (lamina spiralis ossea) медиально и спиральной связкой улитки (lig. spirale cochleae) латерально. Базилярная пластинка расширяется по направлению к верхушке улитки. Она состоит из четырех слоев волокон. Над К. о. нависает покровная мембрана (membrana tectoria), медиально связанная с эпителием вестибулярной губы лимба костной спиральной пластинки. Она сохраняет свое положение благодаря наличию среди ее волокнистых структур прочных коллагеновых волокон. Покровная мембрана имеет свободный контакт со стереоцилиями (волосками) волосковых клеток. На поперечном разрезе клеточный массив К. о. разделен на две части — наружную и внутреннюю — треугольным пространством внутреннего (кортиева) туннеля, заполненного так наз. кортилимфой, по своему хим. составу приближающейся к перилимфе. Через туннель проходят безмякотные волокна наружного спирального сплетения и частично эфферентного оливокохлеарного пучка. К. о. состоит из двух видов нейроэпителиальных клеток — волосковых (сенсорно-эпителиальных) клеток [cellulae (sensoriepitheliales) pilosae] и поддерживающих (cellulae sustentantes). По пространственному отношению к внутреннему туннелю волосковые клетки делятся на внутренние и наружные. У человека 3500— 4000 внутренних волосковых клеток [cellulae (sensoriepitheliales) pilosae int.] и 20 000 наружных волосковых клеток [cellulae (sensoriepitheliales) pilosae ext.].
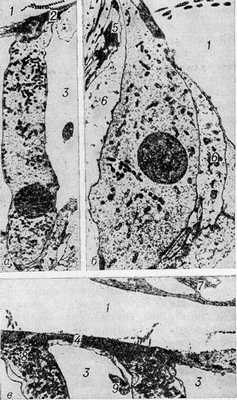
Рис. 2. Электронограммы волосковых клеток кортиева органа: а— наружная волосковая клетка (х 3680); б— внутренняя волосковая клетка (х 4290); в— апикальная область наружных волосковых клеток (X 4320); 1— эндолимфатическое пространство; 2— кутикула; 3— паратуннель (пространство Нюэля); 4— наружная фаланговая клетка (клетка Дейтерса); 5 - клетка-столб; 6— внутренняя опорная клетка; 7— текториальная мембрана; 8 — пластинчатый комплекс (комплекс Гольджи); 9— поперечный срез фаланги наружной фаланговой клетки.
Наружные волосковые клетки имеют цилиндрическую форму (рис. 2, а и в). Их апикальная поверхность омывается эндолимфой, а боковые поверхности — кортилимфой паратуннеля (пространства Нюэля). Волоски этих клеток погружены основаниями в кутикулу— электронно-плотное вещество апикальных частей волосковых клеток. Стереоцилии, помимо сердцевины, содержат многочисленные микрофиламенты — тончайшие плотные нити. Основная масса органелл (митохондрии, канальцы гладкого эндоплазматического ретикулума, микротрубочки, мультивезикулярные тельца, рибосомы и др.) распределена в цитоплазме наружных волосковых клеток, в отличие от внутренних волосковых клеток, неравномерно: в верхней части, вдоль плазматической мембраны и в базальной части клеток. Характерным для волосковых клеток является наличие так наз. пластинчатых каналов, идущих параллельно боковой плазмолемме и связывающих апикальную и базальную части клетки. Последняя богата органеллами, включая так наз. покрытые пузырьки, и контактирует с афферентными и эфферентными нервными окончаниями. Тела афферентных окончаний небольшие, светлые, содержат единичные пузырьки диам. 40—50 нм и митохондрии. Пре- и постсинаптические мембраны имеют примембранные утолщения, а у некоторых видов и синаптическую полоску. Эфферентные нервные окончания намного крупнее, темные от множества пузырьков диам. 30—35 нм. Эфферентный синапс имеет субсинаптическую цистерну. Электронно-микроскопически прослежено вскрытие пузырьков эфферентного окончания с выбросом содержимого в синаптическую щель, а электронно-цитохимически — наличие ацетилхолинэстеразы в области эфферентных и афферентных синапсов, в базальной части волосковых клеток и в стереоцилиях.
Внутренние волосковые клетки имеют кувшинообразную форму (рис. 2, б) и со всех сторон окружены клеточными элементами, за исключением апикальной поверхности, омываемой эндолимфой. Они имеют принципиально сходное с наружными волосковыми клетками строение, однако гораздо беднее их органеллами. Внутренние волосковые клетки связаны с поддерживающими клетками простой межклеточной связью, тогда как наружные фиксированы плотной межклеточной связью в апикальной области и специализированным усиленным контактом в базальной области.
Опорные элементы К. о. [внутренние и наружные клетки столбов (cellulae int. columnarum et cellulae ext. columnarum), наружные фаланговые клетки Дейтерса (cellulae phalangeae ext.) и наружные пограничные клетки Гензена (cellulae limitantes ext.) ] выполняют поддерживающую в отношении волосковых клеток функцию за счет развитой системы плотных межклеточных связей, выраженной тонофибриллярной сети в цитоплазме клеток столбов и наружных фаланговых клеток. Опорные клетки также несут трофическую функцию, обеспечивая транспорт веществ за счет аппарата микро-ворсинок.
Кортиев орган не имеет сосудов. Ближайший сосуд — vas spirale — располагается на барабанной поверхности базилярной пластинки. Основное значение в трофике К. о. принадлежит сосудистой полоске. Она состоит из трех видов эпителиальных клеток (маргинальных, интермедиарных и базальных), тесно связанных морфологически и функционально с эндотелиальными клетками. Эндолимфатическую поверхность сосудистой полоски составляют маргинальные клетки, цитоплазма которых насыщена органеллами, в особенности митохондриями. Донные части этих клеток представляют собой сложнейшую и разветвленную систему отростков, пронизывающую всю толщину сосудистой полоски и, как щупальцами, охватывающую капилляры. Энзиматическая активность сосудистой полоски чрезвычайно высока, в особенности это относится к ферментам окислительного метаболизма. По совр, представлениям сосудистая полоска создает в улитке постоянный потенциал покоя, обеспечивает насыщение эндолимфы кислородом, определяет ее состав (в частности, своеобразное, интрацеллюлярное распределение в этой среде ионов калия и натрия). Нарушение ионного равновесия в эндо- и перилимфе ведет к нарушению слуховой функции. Конгенитальная патология сосудистой полоски лежит в основе врожденной глухоты, ее экспериментальное повреждение вызывает глубокое нарушение функции К. о. вплоть до его гибели.
Эндолимфа резорбируется в эндолимфатическом мешке (saccus endolymphaticus). Она изоосмотична с перилимфой, хотя и имеет количественно отличный от нее состав. Обмен между жидкостями возможен гл. обр. через преддверную мембрану. В происхождении перилимфы основная роль принадлежит внутрилабиринтным источникам — процессу ультрафильтрации из сосудистых зон. Обе жидкости функционально едины и представляют собой целостную жидкостную систему внутреннего уха. Нарушения циркуляции, хим. состава, давления лабиринтной лимфы лежат в основе многих заболеваний, в частности болезни Меньера (см. Меньера болезнь), кохлеарного неврита (см. Преддверно-улитковый нерв), старческой тугоухости (см. Пресбиакузис), акустической травмы (см.), поздних стадий отосклероза (см.), тугоухости травматического происхождения и др. Гомеостаз внутренних лабиринтных сред обеспечивается функциональной активностью гематолабиринтного барьера. По сравнению с гематоэнцефалическим и гематоофтальмическим барьерами стабильность этого гистогематического барьера весьма высока: он является преградой для многих соединений, сохраняет свою инертность при значительных общих расстройствах гемодинамики. Однако при терапии антибиотиками (см.) группы аминогликозидов наступает нарушение проницаемости барьера. Отмечается феномен истинной кумуляции стрептомицина, неомицина, канамицина и других антибиотиков в лабиринтной лимфе К. о. с избирательным и необратимым поражением первоначально наружных, а затем и внутренних волосковых клеток.
В физиологических условиях в наружных волосковых клетках возникает микрофонный потенциал — переменный биоток с частотой, синхронной тону озвучивания. Амплитуда потенциала в известных пределах пропорциональна интенсивности звукового раздражения. Форма звукового стимула в точности воспроизводится в форме волны микрофонного потенциала. Благодаря электропроводности перилимфы и эндолимфы регистрация микрофонного потенциала в эксперименте осуществима с окна улитки. Запись этого биотока (кохлеограмма) и ее анализ имели большое значение для развития экспериментальной отологии. Проводятся успешные попытки регистрации этого потенциала у человека с промонториальной стенки барабанной полости и даже барабанной перепонки.
В улитке возникают также и другие биоэлектрические явления. Суммационный потенциал отражает функц, активность внутренних волосковых клеток, акционный потенциал — ганглиозных слуховых клеток, а эндолимфатический потенциал покоя — клеток сосудистой полости. Дейвис полагает, что эндолимфатический потенциал играет роль усилителя микроэлектрических явлений в К. о. и определяет его колоссальную чувствительность. Акционный потенциал представляет собой первичную кодированную информацию о звуке. Механизм раздражения К. о., по мнению Дейвиса, состоит в сгибании стереоцилий волосковых клеток; Я. А. Винников и Л. К. Титова считают, что механизм раздражения волосковых клеток К. о. состоит в деполяризации мембраны волосков ацетил холином, растворенным в эндолимфе.
Кортиев орган — самый сложный прибор рецепции млекопитающих. Это один из двух основных дистантных рецепторов. Чувствительность органа близка к предельной, т. к. он способен реагировать на колебательный процесс, приближающийся по своим параметрам к броуновскому движению. Слуховой рецептор функционирует в течение всей жизни постоянно, без пауз. В нем осуществляется совершенный первичный анализ звука, т. е. математическое разложение сложного звука на составляющие, благодаря дифференцированному восприятию частот отдельными его участками. После трансформации механической энергии звука в волосковых клетках в нервный импульс последний по проводящим путям поступает в кору височной доли, где осуществляется высший анализ звука (см. Слуховой анализатор). Высокие тона воспринимаются у основания улитки, а низкие — у ее верхушки (резонансная теория Гельмгольца). Биофиз, обоснование феномена резонанса в К. о. остается предметом исследований (см. Слух).
Патология К. о. наблюдается при многих общих и местных (ушных) заболеваниях. Клиническая диагностика кохлеарного поражения осуществляется с помощью аудиометрии (см.). Лечебные меры в основном направлены на сохранение остаточной слуховой функции.
Библиография: Андреев А. М., Арапова А. А. и Гершуни Г. В. О потенциалах улитки у человека, Физиол, журн. СССР, т. 26, № 2-3, с. 205, 1939; Винников Я. А. и Титова Л. К. Кортиев орган, М.— Л., 1961, библиогр.; Лавдовский М. Д. Гистология концевого аппарата улиткового нерва, дисс., Спб., 1874; Многотомное руководство по оториноларингологии, под ред. А. Г. Лихачёва, т. 1, с. 161 и др., М., 1960; Вekesу G. Experiments in hearing, L., 1960, bibliogr.; Claudius M. Bemerkungen uber der Bau des hautigen Spiralleiste der Schnecke, Z. wiss. Zool., Bd 8, S. 154, 1855; Сorti А. Recherches sur l’organe de l’ouie des mammiferes, ibid., Bd 3, S. 109, 1851; Davis H. A mechano-electrical theory of cochlear action, Ann. Otol. (St Louis), v. 67, p. 789, 1958; Deiters O. Beitrage zur Kenntniss der Lamina spiralis membranacea, Z. wiss. Zool., Bd 10, S. 1, 1860; Helmholtz H. Die Lehre von den Tonempfindungen als physiologische Grundlage fiir die Theorie der Musik, Braunschweig, 1868; HensenV. Zur Morphologie der Schnecke des Menschen und der Saugethiere, Z. wiss. Zool., Bd 13, S. 481, 1863; он же, Uber Boettcher’s Entwicklung und Bau des Gehorlabyrinths nach eigenen Untersuchungen, Arch. Ohrenheilk., Bd 6, S. 1, 1871; Hunter-Duvar I. M. Electron microscopic assessment of the cochlea, Acta oto-laryng. (Stockh.), Suppl. 351, 1978; Neural mechanisms of the auditory and vestibular systems, ed. by G. L. Rasmussen a. W. F. Windle, Springfield, 1960; Nuel, Beitrag zur Kenntniss der Sauge-thierschnecke, Arch. mikr. Anat., Bd 8, S. 200, 1872; Rauch S. Biochemie des Hororgans, Stuttgart, 1964; Wever E. G. a. Bray C. W. Aetion currents in the auditory nerve in response to acoustical stimulation, Proc. nat. Acad. Sci. (Wash.), v. 16, p. 344, 1930.
периферическая часть звуковоспринимающего аппарата (рецептор слухового анализатора (См. Слуховой анализатор)) у млекопитающих животных и человека. Открыт итальянским гистологом А. Корти (A. Corti; 1822—76). В процессе эволюции возникает на основе структур боковых органов (См. Боковые органы). К. о. располагается в спирально завитом костном канале внутреннего уха (См. Внутреннее ухо) — улитковом ходе, заполненном эндолимфой. Верхняя стенка хода прилегает к т. н. лестнице преддверия и называется рейснеровой перепонкой; нижняя стенка, граничащая с т. н. барабанной лестницей, образована основной перепонкой, прикрепляющейся к спиральной костной пластинке. Лестницы преддверия и барабанная заполнены перилимфой. Наружная стенка улиткового хода, или сосудистая полоска, содержит много кровеносных сосудов. К. о. расположен на основной перепонке и состоит из внутренних и наружных волосковых клеток, внутренних и наружных опорных клеток (столбовых, клеток Дейтерса, Клаудиуса, Гензена), между которыми находится туннель, где проходят направляющиеся к основаниям волосковых клеток отростки нервных клеток, лежащих в спиральном нервном ганглии. Воспринимающие звук волосковые клетки располагаются в нишах, образуемых телами опорных клеток, и имеют на поверхности, обращенной к покровной перепонке, по 30—60 коротких волосков. Опорные клетки выполняют также трофическую функцию, направляя поток питательных веществ к волосковым клеткам. Функция К. о. — преобразование (трансформация) энергии звуковых колебаний в процесс нервного возбуждения. Звуковые колебания воспринимаются барабанной перепонкой и через систему косточек среднего уха передаются жидким средам внутреннего уха — перилимфе и эндолимфе. Колебания последних приводят к изменению взаиморасположения волосковых клеток и покровной перепонки К. о., что вызывает сгибание волосков и возникновение биоэлектрических потенциалов (См. Биоэлектрические потенциалы), улавливаемых и передаваемых в центральную нервную систему отростками нейронов спирального ганглия, подходящими к основанию каждой волосковой клетки. По др. представлениям, волоски звуковоспринимающих клеток — лишь чувствительные антенны, деполяризующиеся под действием приходящих волн за счёт перераспределения ацетилхолина эндолимфы. Деполяризация вызывает цепь химических превращений в цитоплазме волосковых клеток и возникновение нервного импульса в контактирующих с ними нервных окончаниях. Различающиеся по высоте звуковые колебания воспринимаются различными отделами К. о.: высокие частоты вызывают колебания в нижних отделах улитки, низкие — в верхних, что связано с особенностями гидродинамических явлений в ходе улитки.
Лит.: Винников Я. А., Титова Л. К., Кортиев орган. Гистофизиология и гистохимия, М.— Л., 1961; Wandsronk С., On the mechanism of hearing, [Leiden], 1961; Vinnikov Ya. A., The ultrastructural and cytochemical bases of the mechanism of function of the sense organ receptors, в кн.: The structure and function of nervous tissue, v. 2, N. Y.— L., 1969, p. 265—392.
Разрез улитки и кортиева органа (схема): 1 — лестница преддверия; 2 — барабанная лестница; 3 — улитковый ход; 4 — рейснерова перепонка; 5 — сосудистая полоска; 6 — спиральная пластинка; 7 — основная перепонка; 8 — внутренние и 9 — наружные волосковые клетки; 10 — внутренние и 11 — наружные опорные клетки; 12 — клетки Дейтерса; 13 — клетки Клаудиуса; 14 — клетки Гензена; 15 — туннель; 16 — спиральный ганглий; 17 — покровная перепонка.
Большая советская энциклопедия. — М.: Советская энциклопедия . 1969—1978 .
Читайте также: