Опорно-двигательная система клетки. Промежуточные филаменты клетки.
Добавил пользователь Alex Обновлено: 28.01.2026
Цитоскелет (опорно-двигательная система, иди каркасно-двигательная система клетки). Опорно-двигательная система клетки образована тремя основными компонентами: микротрубочками, микрофиламентами и промежуточными филаментами.
Функции: 1) обеспечивает поддержание формы клеток, осуществляет изменение объема и формы клетки, передвижение ее в пространстве, 2) образует опорный каркас для всех клеточных структур, обеспечивает фиксацию составных частей клетки в определенном положении и перемещение их относительно друг друга; 3) участвует в образовании других органоидов (ресничек, жгутиков, центриолей и др.) и межклеточных контактов; 4) через цитоскелет происходит взаимодействие с белками внеклеточного матрикса (фибронектином, ламинином).
14. Микрофиламенты и промежуточные филаменты
Микрофиломенты (от греч. micros - малый и лат. filamentum — нить) - органоиды, состоящие из двух спирально закрученных цепочек, образованных короткими молекулами белка актина, на долю которого приходится более 10% всех белков клетки.
Тонкие микрофиламенты (МФ) - очень тонкие белковые нити диаметром 4-7 нм, состоящие из белка актина. В клетке актин присутствует в двух состояниях: в виде отдельных глобулярных (шаровидных) субъединиц или в виде филаментов (нитей), образованных в результате полимеризации глобулярного актина. В состав МФ могут входить также тропонин, тропомиозин и другие белки. МФ полярны - они имеют плюс-конец, который растет за счет полимеризации актина и минус-конец, где происходит деполимеризация.
Функции микрофиламентов:
1) образуют пучки, служащие опорой для различных внутриклеточных структур;
2) образуют сократительные системы, обеспечивая клеточную подвижность. Формируют временные органоиды перемещения (псевдоподии, ламеллоподии). Лежат в основе мышечного сокращения (нити миозина скользят относительно нитей актина). Участвуют в обеспечении всех форм движения. Влияют на действие
3) МФ образуют кортикальную сеть, которая близко подходит к плазмалемме и при помощи
белка винкулина (с участием а-актинина и спектрина) присоединяется к белку интегрину
цитоплазматической мембраны; 4) МФ образуют нити натяжения, расположенные по радиусам
клетки. Нити представляют собой спирально скрученные волокна.
5) актиновые МФ принимают участие в образовании микроворсинок (рис.8). Микроворсинки
находятся на апикальных поверхностях эпителиоцитов тонкой кишки.
Функции ПФ изучены недостаточно; установлено, однако, что они не влияют ни на движение, ни на деление клетки. Выполняют главным образом структурные функции, например, противодействуют растягивающим силам. К их основным функциям относятся: • структурная ;• обеспечение равномерного распределения сил деформации; * участие в образовании рогового вещества; • формообразующая - поддержание формы отростков нервных клеток; • удержание миофибрилл в мышечной ткани.
- Опорно-двигательная система клетки
Одним из важнейших свойств любой клетки является подвижность. Подвижность клетки проявляется различными формами движения (внутриклеточное, амебовидное, мерцательное, сократительное). Несмотря на разнообразие форм, в основе любых двигательных реакций лежат общие молекулярные механизмы. Кроме того, двигательные аппараты (реснички, жгутики) структурно связаны с существованием опорных внутриклеточных структур. Это позволяет говорить об опорно-двигательной системе клеток.
1. Центриоли, структура, репликация, участие в делении клетки
Клеточный центр, или центросома, представляет собой немембранный органоид, локализованный около ядра и состоящий из двух центриолей и центросферы. Причем постоянным и наиболее важным компонентом клеточного центра являются центриоли. Этот органоид обнаружен в клетках животных, низших растений и грибов.
Центриоли (от лат. centrum - срединная точка, центр) представляют два перпендикулярно расположенных друг к другу цилиндра, стенки которых образованы микротрубочками и соединены системой связок. Конец одного цилиндра (дочерняя центриоль) направлен к поверхности другого (материнская центриоль). Совокупность сближенных между собой материнской и дочерней центриолей называетя диплосомой. Впервые центриоли были обнаружены и описаны в 1875 В. Флемингом. В интерфазных клетках центриоли часто располагаются возле комплекса Гольджи и ядра.
Ультрамикроскопическое строение центриолей было изучено только с помощью электронного микроскопа. Стенку центриолей составляют расположенные по окружности 9 триплетов микротрубочек, образующих полый цилиндр. Системы микротрубочек центриоли можно описать формулой (9X3) + 0, подчеркивая отсутствие микротрубочек в центральной части. Ширина центриоли составляет около 0,2 мкм, длина - 0,3-0,5 мкм (однако, есть центриоли, достигающие в длину нескольких микрометров). Кроме микротрубочек в состав центриоли входят дополнительные структуры - "ручки", соединяющие триплеты.
Центросфера - плотный слой цитоплазмы вокруг центриолей, в котором часто содержатся микротрубочки, расположенные лучами.
Центриолярный цикл. Строение и активность центриолей меняются в зависимости от периода клеточного цикла. Это позволяет говорить о центриолярном цикле. В начале периода G1 от поверхности материнской центриоли начинается рост микротрубочек, которые растут и заполняют цитоплазму. По мере роста микротрубочки теряют связь с областью центриолей и могут находиться в цитоплазме длительное время. В периоде S или G2 происходит удвоение числа центриолей. Этот процесс заключается в том, что центриоли в диплосоме расходятся и около каждой из них происходит закладка процентриолей. В начале вблизи и перпендикулярно исходной центриоли закладываются девять одиночных микротрубочек. Затем они преобразуются в девять дуплетов, а потом в девять триплетов микротрубочек новых центриолей. Этот способ увеличения числа центриолей был назван дупликацией. Следует отметить, что удвоение числа центриолей не связано с их делением, почкованием или фрагментацией, а происходит путем образования процентриолей. Таким образом, в результате дупликации в клетке содержатся четыре попарно связанные центриоли. В этом периоде материнская центриоль продолжает играть роль центра образования цитоплазматических микротрубочек. В периоде G2 обе материнские центриоли покрываются фибриллярным гало (зона тонких фибрилл), от которого в профазе начнут отрастать митотические микротрубочки. В этом периоде в цитоплазме происходит исчезновение микротрубочек и клетка стремиться приобрести шаровидную форму. В профазе митоза диплосомы расходятся к противоположным полюсам клетки. От фибриллярного гало материнской центриоли отходят микротрубочки, из которых формируется веретено деления митотического аппарата. Таким образом, центриоли являются центрами организации роста микротрубочек. В телофазе происходит разрушение веретена деления. Следует отметить, что в клетках высших растений, некоторых водорослей, грибов, ряда простейших центры организации роста микротрубочек центриолей не имеют. У некоторых простейших центрами индукции образования микротрубочек выступают плотные пластинки, связанные с мембраной.
2. Строение ресничек и жгутиков эукариотических клеток
Реснички и жгутики - органоиды специального назначения, выполняющие двигательную функцию и выступающие из клетки. Различий в ультрамикроскопическом строении ресничек и жгутиков нет. Жгутики отличаются от ресничек лишь длиной. Длина ресничек составляет 5-10 мкм, а длина жгутиков может достигать 150 мкм. Диаметр их составляет около 0,2 мкм. Причем клетки, имеющие реснички и жгутики, в свободном состоянии обладают способностью двигаться. Неподвижные клетки, благодаря движению ресничек, способны перемещать жидкости и частички веществ.
Жгутик - это органоид движения у бактерий, ряда простейших, зооспор и сперматозоидов. В клетке обычно бывает от 1 до 4 жгутиков.
Ресничка - это органоид движения или рецепции у клеток животных и некоторых растений. Траектория движения ресничек очень разнообразна. В различных клетках это движение может быть маятникообразным, крючкообразным, воронкообразным или волнообразным.
Ресничка представляет собой тонкий цилиндрический вырост цитоплазмы, покрытый цитоплазматической мембраной. Внутри выроста расположена аксонема ("осевая нить"), состоящая в основном из микротрубочек. В основании реснички находится базальное тело, погруженное в цитоплазму. Диаметры аксонемы и базального тельца одинаковы (около 150 нм). Базальное тельце, как и центриоли, состоит из 9 триплетов микротрубочек и имеет "ручки". Часто в основании реснички лежит не одна, а пара базальных телец, располагающихся под прямым углом друг к другу подобно диплосоме - центриоли. Аксонема в отличие от базального тельца или центриоли имеет 9 дублетов микротрубочек с "ручками", образующих стенку цилиндра аксонемы. Кроме периферических дублетов микротрубочек, в центре аксонемы располагается пара центральных микротрубочек. В целом систему микротрубочек реснички описывают как (9 х 2) + 2 в отличие от (9 х 3) + 0 системы центриолей и базальных телец. Базальное тельце и аксонема структурно связаны друг с другом и составляют единое целое: две микротрубочки триплетов базального тельца являются микротрубочками дублетов аксонемы.
Для объяснения способа движения ресничек и жгутиков используется гипотеза "скользящих нитей". Считается, что незначительные смещения дублетов микротрубочек друг относительно друга могут вызвать изгиб всей реснички. Если такое локальное смещение будет происходить вдоль жгутика, то возникает волнообразное движение.
3. Фибриллярные структуры цитоплазмы
Если Вам понравилась эта лекция, то понравится и эта - 6.1. Структура HTML-документов.
Цитоплазма клетки представляет собой вязкую жидкость, поэтому из-за поверхностного натяжения клетка должна иметь шаровидную форму. Однако помимо шаровидной встречается множество других форм клеток (кубические, призматические, звездчатые, дисковидные, с разнообразными отростками и другие). Форма определяется с помощью жестких, параллельно расположенных волокон. Эти волокна называются фибриллярными структурами цитоплазмы. К ним относятся микротрубочки, микрофиламенты и промежуточные филаменты. Эти структуры образуют цитоскелет клетки (опорно-двигательная система). Цитоскелет определяет форму клетки, участвует в передвижении клетки, во внутриклеточном транспорте органоидов и отдельных соединений.
Микротрубочки - немембранные органоиды, представляющие собой полые цилиндры длиной около 200 нм и толщиной около 25 нм. Микротрубочки можно обнаружить в цитоплазме практически всех эукариотных клеток. В больших количествах они находятся в цитоплазматических отростках нервных клеток, фибробластов и других изменяющих свою форму клеток. Местом организации роста микротрубочек цитоскелета в интерфазной клетке является центриоль. Микротрубочки различного происхождения (реснички простейших, клетки нервной ткани, веретено деления) имеют сходный состав и содержат белки - тубулины. Очищенные тубулины при определенных условиях способны собираться в микротрубочки. Добавление алкалоида колхицина предотвращает самосборку микротрубочек или приводит к разборке уже существующих. В клетке тубулины существуют в двух формах - свободной и связанной. Сдвиг равновесия между этими формами приводит или к диссоциации микротрубочек, или к их росту. Ни тубулины в чистом виде, ни построенные из них микротрубочки не способны к сокращению. В клетках микротрубочки принимают участие в создании ряда временных (цитоскелет интерфазных клеток, веретено деления) или постоянных структур (центриоли, базальные тельца, реснички, жгутики). Кроме того, микротрубочки способствуют ориентированному движению внутриклеточных компонентов.
Микрофиламенты представляют собой тонкие (около 7 нм) белковые нити, встречающиеся практически во всех типах эукариотических клеток. Они расположены пучками или слоями в кортикальном слое цитоплазмы, непосредственно под плазмолеммой. Сеть микрофиламентов выявлена в большинстве клеток. В состав микрофиламентов входят сократительные белки: актин, миозин, тропомиозин, альфа-актинин. Функции микрофиламентов заключаются в образовании цитоскелета и обеспечении большинства внутриклеточных форм движения (токи цитоплазмы, движение вакуолей, митохондрий).
Промежуточные филаменты имеют толщину около 10 нм и также являются белковыми структурами. Это тонкие неветвящиеся, часто располагающиеся пучками нити. Характерно, что их белковый состав различен в разных тканях. В эпителии в состав промежуточных филаментов входит кератин. Пучки кератиновых промежуточных филаментов в эпителиальных клетках образуют тонофибриллы. В состав промежуточных филаментов клеток мезенхимальных тканей (например, фибробластов) входит другой белок - виментин, в мышечные клетки - десмин, в нервных клетках в состав их нейрофиламентов также входит особый белок. Промежуточные филаменты выполняют опорно-каркасную функцию.
Промежуточные филаменты
Промежуточные филаменты состоят из большой группы родственных белков, которую делят на четыре типа. Первый тип - кератины, кислые и нейтральные, встречающиеся в эпителиальных клетках; они образуют гетерополимеры из этих двух подтипов. Кератины, кроме того, имеют некоторую гетерогенность, зависящую от тканевого источника. Так, в эпителиях встречается до 20 форм кератинов, 10 форм других кератинов найдено в волосах и ногтях. Молекулярный вес кератинов колеблется от 40 до 70 тыс.
Второй тип белков ПФ включает в себя три вида белков, имеющих сходный молекулярный вес (45-53 тыс.). Это - виментин, характерный для клеток мезенхимного происхождения, входящий в состав цитоскелета клеток соединительной ткани, эндотелия, клеток крови. Десмин - характерен для мышечных клеток, как гладких, так и исчерченных. Глиальный фибриллярный белок входит в состав ПФ некоторых клеток нервной глии - в астроциты и некоторые Шванновские клетки. Периферин - входит в состав периферических и центральных нейронов.
Третий тип - белки нейрофиламентов (мол. вес от 60 до 130 тыс.) встречается в аксонах нервных клеток.
И наконец, четвертый тип - белки ядерной ламины. Хотя эти последние имеют ядерную локализацию, они сходны по строению и свойствам со всеми белками промежуточных филаментов.
Как уже говорилось, промежуточные филаменты, построены из фибриллярных белков наподобие каната. При этом некоторые белки могут образовывать сополимеры, например виментин с десмином, или виментин с глиальными белками.
Все белки промежуточных филаментов обладают сходной аминокислотной последовательностью из 130 остатков в центральной части фибриллярной молекулы, которая обладает a-спиральным строением. Концевые же участки молекул имеют разные последовательности аминокислот, разную длину, и не имеют a-спирального строения. Наличие протяженных a-спиральных участков позволяет двум молекулам образовывать двойную спираль, подобно тому, что приводит к образованию палочковидного димера, длиной около 48 нм. Два димера, объединяясь бок о бок, образуют короткий протофиламент, тетрамер, толщиной около 3 нм. Такие протофиламенты могут объединяться в более толстые и длинные фибриллы и в конечном итоге в промежуточный полный филамент, состоящий из 8 продольных протофиламентов (рис. 242).
Белки ядерной ламины образуют рыхлую прямоугольную решетку. Такие слои ламины быстро разрушаются во время митоза при фосфорилировании ламинов.
Цитоплазматические промежуточные филаменты относятся к самым стабильным и долгоживущим элементам цитоскелета.
Особенности строения и химическая устойчивость промежуточных филаментов определяет их физическую устойчивость. Они служат как бы истинно опорной системой в клетках подвергающихся значительным физическим нагрузкам. В клетках кожного эпидермиса промежуточные филаменты образуют пучки (тонофиламенты), связанные с десмосомами, и создают жесткую внутриклеточную сеть (рис. 243). Так, в нервных аксонах, простирающихся на многие десятки сантиметров, ПФ или нейрофиламенты создают жесткую основу, обеспечивающую гибкость и целостность тонких цитоплазматических отростков нервных клеток. В поперечно исчерченных мышечных клеток десминовые филаменты входят в состав z-дисков и связывают их друг с другом как в составе саркомера, так и в соседних миофибриллах, а также с плазматической мембраной.
Вопрос 4. Опорно-двигательная система клетки (Цитоскелет)
Цитоскелет - это совокупность фибриллярных компонентов цитоплазмы эукариотических клеток. Основными типами фибрилл в составе цитоскелета являются актиновые филаменты, микротрубочки и промежуточные филаменты. Относительно функций промежуточных филаментов известно очень мало. Предполагается, что промежуточные филаменты играют механическую роль в межклеточных взаимодействиях и в организации тканевой структуры. В состав промежуточных филаментов у клеток из различных тканей входят следующие белки: виментин, десмин, глиальный фибриллярный кислый белок, белки нейрофиламентов, кератины, ламины.
Схема строения актиновых микрофиламентов на Рис. 11 (А).
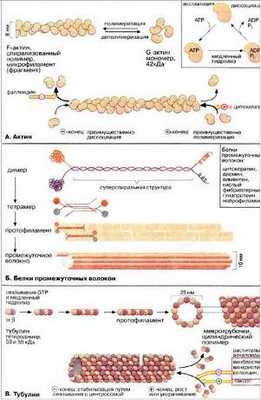
Актиновые филаменты или фибриллярный актин (F-актин) представляют собой тонкие фибриллы диаметром 6-8 нм. Они являются результатом полимеризации глобулярного актина - G-актина. Актиновые филаменты играют ключевую роль в сократительном аппарате мышечных и немышечных клеток, а также принимают участие во многих других клеточных процессах, таких как подвижность, поддержание формы клеток, цитокинез. Существуют системы в которых движение органелл происходит по актиновым филаментам. Движение органелл по актиновым филаментам опосредуется миозинами (актин-связывающими белками), которые объединяются в одну группу благодаря наличию общего "головного" домена, обладающего АТФазной активностью.
Схема строения микротрубочки на Рис. 12.
Белком микротрубочек является тубулин, полимеризация которого приводит к образованию этих структур. Еще есть белки, ассоциированные с микротрубочками (MAP) и белки - транслокаторы. Тубулин - очень консервативный белок, связывающий две молекулы ГТФ, и может также связывать ионы кальция и магния. Функция динеина в биении ресничек и жгутиков, преобразуя энергию АТР в механическое усилие. Микротрубочки имеют две основные функции - двигательную и структурную. Двигательная функция заключается в том, что по микротрубочкам с помощью специальных транспортных белков - транслокаторов - осуществляется движение клеточных органелл. Структурная функция состоит в поддержании с помощью микротрубочек определенной формы клетки или ее части.
В клетках концы микротрубочек, как правило, ассоциированы со специальными структурами - ЦОМТ (центр организации микротрубочек). Некоторые ЦОМТ (например, центриоли, базальные тельца) содержат систему сложно организованных микротрубочек. Другие же представляют собой скопления аморфного электронно-плотного материала. Эти скопления могут быть ассоциированы с другими клеточными компонентами, такими как центриоли, базальные тельца, ядерная оболочка, плазматическая мембрана. Типы ЦOМТ: центросомы, базальные тельца, кинетохоры. Рис. 13 (Б, кружочек).
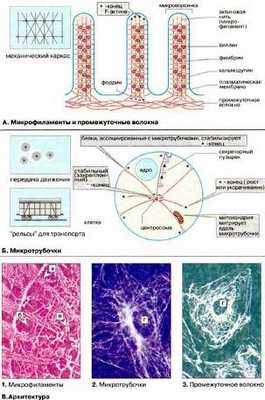
Рис. 13 (Б, кружочек)
В большинстве животных клеток область, где впервые образуется веретено, содержит центриоли. Каждая пара центриолей в митозе становится частью митотического центра, от которого лучами расходятся микротрубочки. Между тем у многих организмов, в том числе у высших растений, функционально полноценное веретено образуется при полном отсутствии центриолей. Кроме того, если у живой клетки разрушить лазерным микролучом центриоли, то митотическое веретено продолжает нормально функционировать. По-видимому, центриоли не являются структурами, необходимыми для сборки микротрубочек веретена; но если центриоли в клетке есть, то они играют роль фокусов, в которых сходятся микротрубочки.
Опорно-двигательная система клетки
К собственно двигательным компонентам клеток относятся различные микрофиламенты и белки, ассоциированные с микротрубочками.
К опорным или скелетным внутриклеточным структурам относят микрофибриллы и микротрубочки.
Микрофиламенты представляют собой тонкие белковые нити, встречающиеся в различных участках клеток, и являются элементарными структурами, обеспечивающими движение. Примером микрофиламентов является миофибрилла поперечнополосатой мышечной ткани, состоящая из актина, миозина, α-актинина, тропомиозина. Движение осуществляется в результате энергозависимого скольжения нитей актина и миозина относительно друг друга.
Микрофибриллы, или промежуточные филаменты, являются более тонкими структурами, представляют собой пестрый класс белков. Они связываются с многочисленными местами на поверхности цитоплазматической мембраны и являются скелетными структурами, обеспечивающими механическую прочность клеток, постоянно подвергающихся большим деформирующим нагрузкам. В эпителиальной ткани известны кератаны, в соединительной - виментин, в мышечной - десмин, в нервной - нейрофибрилл. Эти белки могут сополимеризоваться, в одном типе клеток могут существовать два рода микрофибрилл, например, виментиновая сеть вокруг ядра и одновременно кератиновые филаменты на периферии клетки. Вероятно, что промежуточные филаменты каким-то образом участвуют в заякоривании ядра в объеме цитоплазмы.
Микротрубочки - самые тонкие опорно-двигательные структуры, состоят из белков и не имеют мембранного строения, встречаются в цитоплазме всех эукариотических клеток и не обнаруживаются у прокариот. Микротрубочки представляют собой прямые, не ветвящиеся длинные полые цилиндры. Самосборка трубочек идет при наличии двух видов белков тубулинов (α и β), ГТФ, дополнительных белков полимеризации, ионов магния и при отсутствии ионов кальция. В цитоплазме микротрубочки выполняют две функции - скелетную (стабилизируют форму клетки) и двигательную (создание системы упорядоченного движения типа рельс). В живых клетках процесс новообразования микротрубочек связан с так называемыми центрами организации микротрубочек, центриолями.
Центриоли
Центриоли характерны для клеток животных, их нет у высших растений, низших грибов и некоторых простейших. В делящихся клетках центриоли принимают участие в формировании веретена деления и располагаются на его полюсах. В неделящихся клетках центриоли часто определяют полярность клеток и лежат рядом с ядром. Часто с центриолями ассоциированы мембраны аппарата Гольджи. Возможно, это отражает участие элементов клеточного центра в движении вакуолей.
Основу строения центриолей составляют расположенные по окружности девять триплетов микротрубочек, образующих таким образом полый цилиндр. Первая микротрубочка триплета (А-микротрубочка) состоит из 13 глобулярных единиц. Вторая и третья (В и С) микротрубочки отличаются тем, что они являются неполными, содержат 11 субъединиц и вплотную примыкают к своим соседям. От А-микротрубочек отходят так называемые ручки, выросты, один из которых (внешний) направлен к С-микротрубочке соседнего триплета, а другой (внутренний) - к центру цилиндра. Центральная часть цилиндра центриоли занята структурой, напоминающей тележное колесо. Такие структуры внутри центриоли расположены в одном из ее концов, проксимальном, что делает строение цилиндра центриоли полярным. На дистальном конце центриоли внутри таких структур нет. Систему микротрубочек центриоли обычно описывают формулой 9+0, или (9×3)+0.
Обычно в интерфазных клетках всегда присутствуют две центриоли, располагающиеся рядом друг с другом, образуя диплосому. В диплосоме центриоли располагаются под прямым углом по отношению друг к другу. Из двух центриолей различают материнскую и дочернюю. В дистальном участке материнской центриоли на триплетах располагается аморфный материал в виде шпор. Вокруг каждой центриоли расположен бесструктурный, или тонковолокнистый матрикс. Внутри материнской центриоли образуется особая зона - центросфера - светлая цитоплазма, от которой отходят радиально микротрубочки. Совокупность центриолей и центросферы носит название клеточного центра.
Клеточный центр участвует в процессе расхождения хромосом при делении клеток. Микротрубочки в веретене деления образуются в результате полимеризации тубулинов в зоне центриолей и около специальных структур хромосом - кинетохоров. Кинетохорные микротрубочки перекрываются полюсными и расхождение хромосом происходит в результате их скольжения относительно друг друга.
Формы активности клеточного центра:
формирование веретена митотического аппарата, состоящего из микротрубочек, в это время на материнских центриолях нет шпор, а цитоплазма теряет микротрубочки;
образование цитоплазматических микротрубочек в интерфазных клетках, на центриоли возникают шпоры и от них начинают расти микротрубочки, по мере роста они теряют связь с областью центриолей и могут находиться в цитоплазме длительное время;
удвоение числа центриолей при наступлении S-периода; закладка процентриолей происходит на проксимальных концах центриолей, сперва закладываются девять синглетов микротрубочек, затем они преобразуются в девять дуплетов, а потом в девять триплетов; в этот период материнская центриоль продолжает быть центром образования цитоплазматических микротрубочек.
Читайте также:
- Интраэпителиальная неоплазия конъюнктивы: признаки, гистология, лечение, прогноз
- Лучевые изменения брюшной полости при обструкции верхней полой вены (ВПВ)
- Причины боли в пятке: подошвенный фасцит и пяточная шпора. Диагностика и лечение
- Мужская свобода. Ограничения и поощрения мужчин
- Влияние эстрогенов на организм