Строение комплекса ядерной поры (ЯПК, ядерного порового комплекса)
Добавил пользователь Евгений Кузнецов Обновлено: 21.01.2026
П.А.Я. или кариотека ) имеет микроскопическую толщину и поэтому виден в световой микроскоп. Поверхностный аппарат ядра включает:
а)ядерную оболочку, или кариолемму;. б)паровые комплексы; в)периферическую плотную пластинку (ППП), или ламину.
(1) Ядерная оболочка (кариолемма).состоит из 2 мембран - наружной и внутренней, разделённых перинукляеарным пространством. Обе мембраны имеют такое же жидкосто-мозаичное строение, как и плазматическая мембрана, и различаются по набору белков. Среди этих белков имеются ферменты, переносчики и рецепторы. Наружная ядерная мембрана является продолжением мембран грЭПС и может быть усеяна рибосомами, на которых идёт синтез белка. Со стороны цитоплазмы наружная мембрана окружена сетью промежуточных (ви-ментиновых) фипаментов. Между наружной и внутренней мембранами находится перинуклеарное пространство -полость шириной 15-40 нм, содержимое которого сообщается с полостями каналов ЭПС. По составу перинуклеарное пространство близко к гиалоплазме и может содержать синтезированные рибосомами белки. Главная функция кариолеммы - изоляция гиалоплазмы от кариоплазмы. Специальные белки ядерных мембран, расположенные в области ядерных пор, осуществляют транспортную функцию. Ядерная оболочка пронизана ядерными порами, через которые осуществляется связь кариоплазмы и гиалоплазмы. Для регуляции такой связи в порах находятся (2) поровые комплексы.Они занимают 3-35% поверхности ядерной оболочки. Число ядерных пор с поровыми комплексами является изменчивой величиной и зависит от активности ядра. В области ядерных пор наружная и внутренняя ядерные мембраны сливаются. Совокупность структур, связанных с ядерной порой, называется комплексом ядерной поры. Типичный поровый комплекс представляет собой сложную белковую структуру - содержит более 1000 молекул белка. В центре поры расположена центральная белковая глобула (гранула), от которой по радиусу отходят тонкие фибриллы к периферическим белковым глобулам, образуя диафрагму поры. По периферии ядерной поры находятся две параллельные кольцевые структуры диаметром 80-120 нм (по одному с каждой поверхности кариолеммы), каждое из которых образовано 8 белковыми гранулами (глобулами).
Белковые глобулы перового комплекса подразделяются на центральные и периферические. С помощью периферических глобул осуществляется транспорт макромолекул из ядра в гиалоплазму. (фиксируются в мембране специальным интегральным белком. От этих гранул к центру сходятся белковые фибриллы, формирующие перегородку - диафрагму поры)
В нем участвуют специальные белки периферических глобул - нуклеопорины. В периферических глобулах имеется особый белок - переносчик молекул т-РНК
Центральная глобула специализируется на транспорте и-РНК из ядра в гиалопдазму. В её составе имеются ферменты, участвующее в химической модификации иРНК - ее процессинге.
Гранулы поровых комплексов структурно связаны с белками ядерной ламины, которая участвует в их организации
Функции комплекса ядерной поры:
1. Обеспечение регуляции избирательного транспорта в-в между цитоплазмой и ядром.
2. Активный перенос в ядро белков
3. Перенос в цитоплазму субъединиц рибосом
(3) ППП или ламина
слой толщиной 80-300 нм. прилегает изнутри к внутренней ядерной мембране. Внутренняя ядерная мембрана гладкая, ее интегральные белки связаны с ламиной (периферической плотной пластинкой). Ламина состоит из специальных переплетенных белков-ламинов, образующих периферический кариоскелет. Белки-ламины относятся к классу промежуточных филаментов (скелетных фибрилл). У млекопитающих известно 4 вида этих белков - это ломимы А, В,, В2 и С. Эти белки поступают в ядро из цитоплазмы. Ламины разных видов взаимодействуют между сбой и образуют белковую сеть под внутренней мембраной ядерной оболочки. С помощью ламинов «В» ППП соединяется со спец интеграл белкомядерн оболочки. С ППП взаимодействуют и белки приферич голобул «внутр кольца» порового комплекса. К ламину «А» присоед теломерн участки хромосом.
Функции ламины: 1)поддерд форму ядра. (даже есл бое мембраны разруш, то ядро за счет ламины сохр свою форму и поровые комп-сы ост на своём месте.
2)служит компонентом кариоскелета
3)участв в сборке ядерн оболочки (формирование кариоллемы) при делен клетки.
4)в интерфазном ядре к ламине прикрепл хроматин. таким образом ламина обеспеч функцию фиксации хроматина в ядре (обеспеч упорядочн укладку хроматина, участвует в пространственной организации хроматина в интерфазном ядре). Ламин «А» взаимодейств с теломерными участками хромосом.
5)обеспеч структур организацию поровых комплексов.
импорт и экспор белков.
В ядро через ядерные поры поступают: синтезированные цитоплазматическими рибосомами белки-ферменты, которые участвуют в процессах репликации и репарации (восстановления повреждений в ДНК); белки-ферменты, участвующие в процессе транскрипции; белки-репрессоры, которые регулируют процесс транскрипции; белки-гистоны.(которые связаны с молекулой ДНК и образуют хроматин); белки, входящие в состав субъединиц рибосом: белки ядерного матрикса, образующие кариоскелет; нуклеотиды; ионы минеральных солей, в частности, ионы Са и Mg .
Из ядра в цитоплазму выходят и-РНК. т-РНК и субъединицы рибосом, которые представляют собой рибонуклеопротеидные частицы (р-РНК, связанные с белками).
ЯДЕРНЫЕ ПОРОВЫЕ КОМПЛЕКСЫ
Ядерная оболочка клеток млекопитающих содержит 3-4 тысячи пор (примерно 10 пор на 1 квадратный мкм). Через ядерные поры происходит обмен веществами между ядром и цитоплазмой. Действительно, РНК, синтезируемые в ядре, а также рибосомные субъединицы и белки, содержащие сигналы ядерного экспорта, транспортируются через ядерные поры в цитоплазму, а гистоны, компоненты репликативной системы, многие другие белки импортируются через ядерные поры из цитоплазмы в ядро. Поры окружены большими кольцевыми структурами, называемыми поровыми комплексами (их внутренний диаметр составляет приблизительно 80 нм, а мол. масса -50-100 млн. Каждый комплекс образован набором больших белковых гранул, сгруппированных в октагональную структуру. Поровой комплекс пронизывает двойную мембрану, связывая по окружности поры липидный бислой внутренней и внешней мембран в единое целое. "Дыра" в центре каждого комплекса (ядерная пора) представляет собой водный канал, сквозь который водорастворимые молекулы курсируют между ядром и цитоплазмой. Ядерный поровой комплекс содержит заполненный водой цилиндрический канал диаметром около 9 нм. Большие ядерные белки взаимодействуют с белками-рецепторами, расположенными на границе ядерных пор, и эти рецепторы активно переносят белки в ядро, увеличивая канал поры.
Количество ядерных пор зависит от типа клетки, стадии клеточного цикла и конкретной гормональной ситуации. Для ядерной поры характерна симметрия восьмого порядка, поэтому многие белки ядерной поры представлены в ее составе в количестве, кратном восьми. В электронный микроскоп видны выпуклые кольца. Кольцо, находящееся с ядерной стороны, несет структуру, называемую корзиной (basket). Это образование состоит из обращенных в нуклеоплазму фибрилл и прикрепленного к ним терминального кольца. К просвету канала обращены восемь симметричных образований (spoke complex). В центре комплекса виден вход в канал ядерной поры. Иногда в канале оказывается видна электронноплотная гранула. Некоторые исследователи полагают, что это какой-то транспортирующийся комплекс в момент пересечения ядерной мембраны. Другие считают, что эта структура является функциональной деталью ядерной поры. На основании этого последнего предположения была даже выдвинута не подтвердившаяся впоследствии гипотеза, согласно которой ядерная пора содержит не один, а восемь проницаемых каналов. Молекулы массой менее 5 кДа, проходят через ядерную пору свободно, и равновесие между ядерной и цитоплазматической концентрацией устанавливается за секунды. Для белков массой 17 кДа этот процесс занимает 2 минуты, белков массой 44 кДа (приблизительно 6 нм) - 30 минут. Белки массой более 60 кДа, по-видимому, вообще не могут пассивно проходить через ядерные поры. Проницаемый для гидрофильных макромолекул канал, через который происходит как активный, так и пассивный транспорт, в ядерной поре один, и он, по всей видимости, расположен в центре комплекса. Существуют специальные механизмы транспорта макромолекул внутрь ядра и из ядра в цитоплазму, однако до сих пор о них мало что известно.
ПОРИНЫ
Порины образуют поры, которые функционируют как молекулярные сита, опосредуя диффузию небольших гидрофильных молекул через наружную мембрану грамотрицательных бактерий. Молекулярная масса поринов варьирует от 28000 до 48000. В мембране обычно присутствуют в виде триммеров. Для поринов характерно высокое (до 60%) содержание β-слоев. Наиболее полно к настоящему времени охарактеризованы порины из Escherichia coli: ОmpF (порин матрикса), ОmpС, РhoE, LamB (мальтопорин). Эти белки имеют молекулярную массу ~35000. Их основной особенностью является то, что они образуют наполненный водой трансмембранный канал, причем этот канал образован в основном β-структурами. На рис.4. представлена одна из возможных моделей образования поринового канала из амфифильных β-цепей.
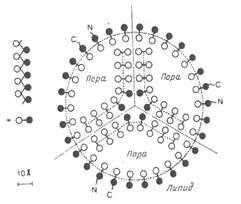
Рис.4.Модель поринового тримера (вид сверху).
Образуемые порином каналы различаются как по размерам (диаметр от 6 до 23Ǻ), так и по селективности. Селективность связана с наличием внутри или около входа заряженных аминокислотных остатков. В одних случаях порины образуют один большой канал, в других - три независимых (рис.4.).
Три из четырех поринов Escherichia coli имеют много общих структурных особенностей, а их аминокислотные последовательности в значительной степени гомологичны. Эти порины образуют поры диаметром 10-12 Ǻ (ОmpF, ОmpС, РhoE).
Порины представляют большой интерес, поскольку они показывают, что трансмембранные каналы могут образовывать не только из α-спиралей, но и из β-слоев.
Строение порового комплекса
В некоторых местах внутренняя и внешняя мембраны ядерной оболочки сливаются и образуют ядерные поры, через которые происходит материальный обмен между ядром и цитоплазмой. Ядерные поры занимают 3-35% поверхности ядерной оболочки. Они более многочисленны в ядрах интенсивно функционирующих клеток и отсутствуют в ядрах спермиев. Количество их зависит от интенсивности деления: чем клетка малдше, тем пор больше. Пора не является дыркой в ядре, а имеет сложную структуру: содержит 2 параллельных кольца (по одному с каждой поверхности кариолеммы), которые образованы 8 белковыми гранулами. От этих гранул к центру сходятся фибриллы, формирующие перегородку (диафрагму), в середине которой лежит центральная гранула. Совокупность структур, связанных с ядерной порой, называется комплексом ядерной поры. Это восьмиугольный цилиндр, состоящий из белков. Белки поровых комплексов структурно связаны с белками ядерной ламины, которая участвует в их организации. Он образует водный канал, по которому по градиенту концентрации свободно диффундируют молекулы воды, растворенные в ней газы и низкомолекулярные вещества, ионы. В результате содержимое ядра отличается от содержимого цитоплазмы по составу высокомолекулярных органических веществ, а по составу низкомолекулярных не отличается. Транспорт макромолекул происходит активно. На внутренней и наружной мембранах комплекса имеются рецепторы, обеспечивающие избирательный транспорт из ядра РНК, субъединиц рибосом (слишком велики для свободного прохождения пор - их транспорт, вероятно, сопровождается изменением конформации порового комплекса), а в ядро избирательно транспортируются синтезированные белки-гистоны и белки ламин, ферменты. Также комплекс ядерной поры обеспечивает регуляцию транспорта веществ между цитоплазмой и ядром.
Студопедия рекомендует:
Формы и виды права собственности Конституция РФ, а так же ГК РФ, предусмотрел 3 основных формы права собственности это.
Управленческие решения ТРЕБОВАНИЯ К УПРАВЛЕНЧЕСКИМ РЕШЕНИЯМ Один из важных показателей деятельности менеджера &mdash.
КОМПЛЕКСНЫЕ ЧИСЛА И ДЕЙСТВИЯ НАД НИМИ Содержание §1. КОМПЛЕКСНЫЕ ЧИСЛА И ДЕЙСТВИЯ НАД НИМИ §2 ПОСЛЕДОВАТЕЛЬНОСТИ КОМПЛЕКСНЫХ ЧИСЕЛ РЯДЫ С.
Профессиональная этика Кодексы профессиональной этики: история и современность. .
Тригонометрические функции, их свойства и графики. Все тригонометрические функции (синус, косинус, тангенс и котангенс) относятся к основным элементарным функциям.
Ядерный поровый комплекс (ЯПК) - восмиугольный цилиндр в состав которого входит ~100-200 белков, М=124x106 D.| Молекулы M9kD перемещаются путем активного транспорта используя ядерный сигнал с затратой энергии ГТФ. Ламина связана с липопротеиновым комплексом внутренней ядерной мембраны. Импортин - белок 60kD узнает сигнальную последовательность из 6 аминокислот - PPKKKKAKV в полипептиде который транспортирует через ядерную оболочку. В ядерном импорте участвуют различные цитоплазматические факторы. ядерные поры - d=7 нм небольшие белки легко проникают в ядро; белки >40 кДа проникают, если содержат сигнальную последовательность.
В начале профазы киназы фосфорилируют ламину, которая разрушается, липопротеиновый комплекс ядерной мембраны распадается на везикулы, наружная мембрана сливается с ЭР, содержимое ядра распределяется в цитозоле. В поздней анафазе цитоплазматические фосфатазы дефосфорилируют ламины, которые полимеризуются на поверхности хромосом, везикулы сливаются образуя ядерную оболочку.
Биосинтез NAD+ -предшественник кофермента, никотинамидмононуклеотид НАМ (NMN) синтезируется в цитоплазме и транспортируется в ядрышко для превращения в динуклеотид NAD+, который после этого возвращается в цитоплазму.
ЯПК - белковый комплекс массой ~125 МДа у метазой и ~66 МДа у грибов, состоящий примерно из 30 различных белков, называемых нуклеопорины.
review 17-19, 26-30. Нуклеопорины разделяют на имеющие FG-повторы, которые непосредственно участвуют в транспорте, и не имеющие FG-повторы, представляющие собой структурные компаненты ЯПК.
Помимо транспортной функции нуклеопорины имеют также дополнительные функциии:
Ядерная пора и ядерный поровый комплекс
Ядерные поры - гигантские макромолекулярные комплексы, которые обеспечивают активный обмен белков и рибонуклеопротеидов между ядром и цитоплазмой. Ядерный поровый комплекс(ЯПК) формирует цилиндр, приблизительно 1200 Ǻ в диаметре и 500 Ǻ толщиной и имеет восьмиугольную симметрию. ЯПК состоит из 100-200 белков; он имеет массу 124х106 дальтон, что примерно в 30 раз больше массы рибосомы.
Этот комплекс - основные ворота для веществ, которые постоянно перемещаются внутрь ядра и из него. Например, матричная РНК (мРНК), субъединицы рибосом, гистоны, рибосомные белки, факторы транскрипции, ионы и мелкие молекулы быстро обмениваются между ядром и полостью эндоплазматического ретикулума или цитозолем.
Механизм ядерного импорта и экспорта
Перемещение молекул из ядра и в него происходит путем активного транспорта, пассивной диффузии или путем специальной ядерной локализации, которая идет посредством сигнальной последовательности определенных белков. Пассивная диффузия и активный транспорт происходят через ядерный поровый комплекс. Мелкие молекулы и ионы (<9кДа) диффундируют через водный канал ЯПК, около 10нм в диаметре. Более крупные молекулы (>9кДа) перемещаются путем активного транспорта с вовлечением ядерного сигнала, а также по энергозависимому механизму.
Ядерный локализационный сигнал.
Роль импортина.
Белки, транспортируемые в ядро, несут ядерный локализационный сигнал (ЯЛС), который содержит значительно обогащенный промежуток из пяти или шести основных аминокислот. Пример- пролин-пролин-лизин-лизин- лизин- лизин-аланин- лизин-валин (Р-Р-К-К-К-К-А-К-V).
Группы основных аминокислот ЯЛС могут локализоваться в любом месте белка. Более того, ядерный локализационный сигнал не изменяется при транслокационных преобразованиях. Особое внимание привлекает тот факт, что 60 кДа белок импортинсвязывается с ЯЛС, инициирует и поддерживает импорт белков. В ядерном импорте также участвуют цитоплазматические факторы.
Растворение ядра и его восстановление
Интерфазные ядра полностью собраны вместе с комплексами пор. Ядерная пластина (ламина) - сетчая структура специальных промежуточных филаментов - формирует волосковую сетеподобную структуру, которая связана с липопротеиновым комплексом внутренней ядерной мембраны.
При вступлении клетки в начало профазы цитозольные киназы фосфорилируют субъединицы ядерных ламин. После фосфорилирования сетеподобная структура разрушается. Затем липопротеиновый компонент внутренней ядерной мембраны распадается на мелкие везикулы, так же как и наружная ядерная мембрана, которая состыкована с ЭР. Затем содержимое ядра распространяется в цитозоле.
Восстановление ядерной оболочки начинается в поздней анафазе, в тот момент, когда цитоплазматические фосфатазы начинают удаление фосфатных остатков из ядерных ламин. Эти белки начинают реполимеризоваться на поверхности конденсированных хромосом. В то же время везикулы, образовавшиеся из внутренней ядерной мембраны, начинают сливаться и формируют оболочку вокруг хромосом. К концу поздней телофазы происходит окончательное слияние внутренней ядерной мембраны. Эти слитые мембраны и дефосфорилированные ламины формируют сетевидную структуру на внутренней поверхности ядерной мембраны.
Митохондрии
Общая структура и функции
Митохондрии -это окруженные двойной мембраной органеллы, которые выполняют функцию метаболического центра клетки. Митохондрии являются местом синтеза аденозинтрифосфата (АТФ). Этот процесс требует участия многих ферментов, большинство из которых поступает из цитозоля.
Процесс импорта ферментов очень сложен и включает несколько этапов. Предполагается, что митохондрии - результат эволюции организмов, которые внедрились в примитивную прокариотическую клетку и сформировали симбиотические отношения с хозяином.
| Признаки | Значение |
| Происхождение | Считается, что митохондрии произошли врезультате эволюции от орагнизмов, которые внедрились в примитивную прокариотическую клетку и стали симбиотами с ней. |
| Форма | Эти орагнеллы могут принимать различные морфологические формы. Некоторые из них имеют сферическую форму, другие лентовидную. |
| Митохондриальная ДНК | Митохондриальная ДНК реплицируется в интерфазе, и этот процесс не синхронизирован с репликацией ДНК в ядре. Митохондриальная ДНК отличается от ядерной ДНК и кодирует особые митохондриальные гены. |
| Синтез белка | Количество транслируемых с митохондриальной мРНК белков ограничено; они формируют субъединицы крупных ферментных комплексов. Митохондрии имеют функционирующие рибосомы, переводящие информацию митохондриальной ДНК в белки, используемые в органелле. |
| Клеточное деление | Во время клеточного цикла митохондрии один раз делятся надвое, образуя при этом перетяжку. Перетяжка деления развивается, начиная с внутренней митохондриальной мембраны. |
Митохондриальная ДНК
В отличие от других орагнелл клетки, митохондрии обладают собственной ДНК, которая отличается от ядерной ДНК и кодирует особые митохондриальные гены. Свойства митохондриальной ДНК:
1) небольшая и содержит около 16,5 кб, то есть приблизительно в 105 раз меньше, чем ДНК, локализованная в ядре;
2) кольцевая и кодирует 2 рибосомные РНК, 22 транспортных РНК (тРНК) и 13 белков.
Генетический код митохондрий, определяющий отдельные аминокислоты, немного отличается от кода ядерной ДНК. Митохондриальный код, например, обладает измененными стоп-кодонами.
Эта органелла обладает функционирующими рибосомами, которые синтезируют белки, используемые в органелле и кодируемые митохондриальной мРНК белка, ограничено и формирует субъединицы более крупных ферментных комплексов. Митохондрии могут принимать различную форму. Обычно митохондрия делится, по крайней мере, один раз в течение клеточного цикла после репликации ее ДНК, которая происходит во время интерфазы. Эта репликация не связана с S-фазой клетки. Деление митохондрии происходит посредством перетяжки на две, которая начинается с образования кольцевой бороздки на внутренней митохондриальной мембране.
Читайте также: