
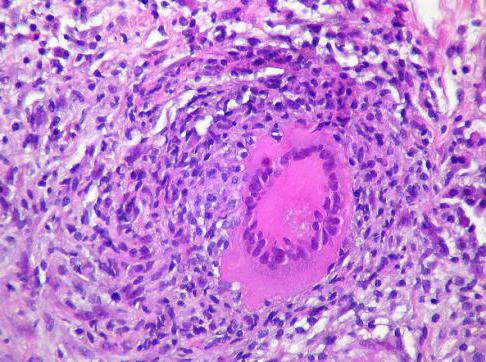
Гигантские клетки пирогова-лангханса при туберкулезе
По своей сути любая гранулема – это узелок (бугорок), образовавшийся в тканях организма по причине начавшегося в них воспаления. Все они подразделяются на неспецифические и специфические. Последние могут быть инфекционными и неинфекционными. В свою очередь, инфекционные классифицируются по типу вызвавшего их микроба - бактериальные или грибковые. Следуя этой классификации, туберкулезная гранулема является специфической, так как наблюдается только при заболевании туберкулез, инфекционной и бактериальной, так как в ее образовании повинна патогенная бактерия Mycobacterium tuberculosis complex. Второе ее название – палочка Коха. Локализуются такие гранулемы в основном в легких, по размеру на начальном этапе бывают с просяное зерно, то есть, милиарными (до 3 мм) или субмилиарными (до 1 мм). Но иногда встречаются туберкулезные гранулемы диаметром 1 и более сантиметров, то есть, солитарные. Без лечения и те, и другие, и третьи способны сливаться и образовывать крупные воспалительные очаги. Рассмотрим строение туберкулезной гранулемы, механизм ее образования, методы выявления и прогноз после лечения.
Как появляется гранулема в легких, ее особенности
Туберкулез может начаться во многих органах, но чаще всего диагностируется в легких. Вызывает его микроскопическое существо – бактерия палочкообразной формы, именуемая Mycobacterium tuberculosis. В организм здорового человека она попадает в основном воздушно-капельным путем при вдыхании воздуха, инфицированного больным туберкулезом человеком. Но возможны и другие пути заражения. Достигнув легких, микроб приступает к размножению и какое-то время ведет себя настолько смирно, что на него не реагирует иммунная система. Однако он, как любое живое существо, выделяет отходы своей жизнедеятельности, для человека являющиеся токсинами. Они в месте локализации бактерий вызывают раздражение тканей. 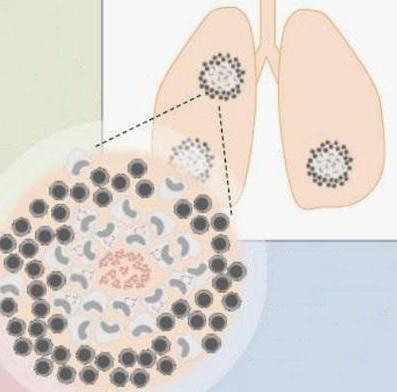
В ответ на это организм человека активизирует свою иммунную систему. К колониям бактерий устремляются антитела, макрофаги и различные типы лимфоцитов. В дальнейшем эти клетки определяют состав туберкулезной гранулемы, которая образуется в месте очага воспаления. Макрофаги созданы природой, чтобы уничтожать раздражители, тем самым помогая организму победить инфекцию. Но в случае с палочкой Коха этот процесс нарушается. Будучи проглоченными макрофагом, микробы не погибают, а продолжают успешно существовать. При этом они получают идеальную защиту от антител, а макрофаг трансформируют для своих нужд. Трансформация заключается в том, что микобактерии задерживают созревание фагосом – специфических органелл макрофагов. Незрелые фагосомы не связываются с лизосомами, то есть, не могут образовать органоид, разрушающий бактерию. Вместо этого незрелые фагосомы начинают взаимодействовать с другими органеллами - ранними эндосомами, в результате чего продуцируются вещества, нужные бактерии для ее питания.
Строение туберкулезной гранулемы
По своей природе гранулематозные узелки являются скоплением клеток, характерных для воспалительных процессов в тканях. Строение туберкулезных гранулем несколько иное. На первых этапах заболевания эти образования являются эпителиодно-клеточными, то есть, без некрозной зоны в центре. С прогрессированием недуга в центре туберкулезных гранулем появляется участок казеозного некроза. Вокруг него располагаются следующие клетки:
- аргирофильные (выявляющиеся при пропитывании серебром) волокна.
Капилляров в самих туберкулезных гранулемах нет, но они присутствуют в наружных зонах.
Некрозная зона
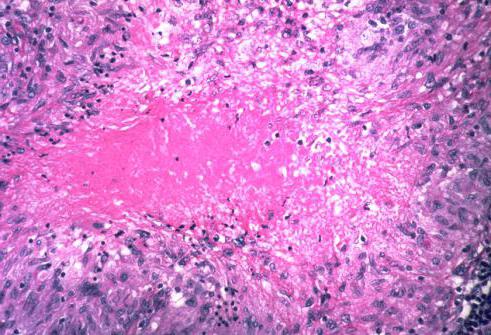
Присутствие казеозного (похожего на творог) центра обуславливает специфичность строения туберкулезной гранулемы. Причин казеификации может быть несколько. Среди них:
- прямое действие цитотоксичных веществ сенсибилизированных макрофагов и/или Т-лимфоцитов;
- слишком быстрая смерть макрофагов;
- спазмы сосудов, ведущие к коагуляционному некрозу.
При неблагоприятном течении заболевания казеификация достигает значительных размеров. Помимо центра гранулемы, такие творожистые очаги образуются в окружающих узелок тканях, пропитанных фибринозно-серозным экссудатом. Прогрессирование недуга способствует разжижению казеозного вещества. Больные начинают его откашливать. При этом в местах, из которых он удаляется, образуются специфические полости (каверны), служащие источником распространения палочек Коха по отделам легких.
Эпителиоидные клетки
Эти структуры есть не что иное, как макрофаги, утратившие способность уничтожать чужеродные агенты, в данном случае палочки Коха. Строение туберкулезной гранулемы таково, что эпителиоидные клетки в ней располагаются вокруг зоны некроза в несколько слоев (иногда эта картина напоминает частокол), образуя циркуляционный ряд. Они довольно крупные, уплощенные, имеют большое ядро, крупное ядрышко и зозинофильную цитоплазму. В тех клетках, что ближе к зоне некроза, наблюдаются деструкция и дистрофия. В процессе своего образования эпителиоидные клетки проходят две стадии: незрелой и зрелой, что отражает кинетику дальнейшего развития гранулематоза.
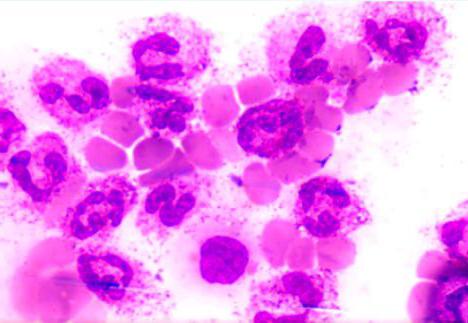
Клетки Пирогова-Лангханса
Эти структуры образуются из клеток эпителиоидных двумя способами:
1. Путем цитодиэреза, что означает деление ядра без деления самих клеток. В результате в каждой из них может насчитываться более 100 ядер, располагающихся по периферии. Благодаря цитодиэрезу, их размеры получаются очень внушительными - до 50 мкм (по другим данным до 300 мкм) . Из-за большой величины, а также из-за многоядерности эти клетки туберкулезной гранулемы получили название гигантских. Также их называют клетками Пирогова-Лангханса.
2. Путем соединения эпителиоидных клеток.
Они считаются показательными для туберкулеза, хотя могут образовываться и при других заболеваниях. Решающим фактором для постановки диагноза является обнаружение в их цитоплазме палочек Коха или их фрагментов.
В цитоплазме гигантских клеток обнаруживаются лизосомы, митохондрии и окислительно-восстановительные ферменты. Если иммунитет человека, подхватившего палочки Коха, высокий, эти макроорганизмы ведут себя как фагоциты, то есть, уничтожают бактерии. В таких случаях усиливается клеточная пролиферация, и воспалительный процесс затихает. Если иммунитет низок, гигантские клетки образуют с бактериями своеобразный симбиоз, в результате которого заболевание прогрессирует.
Классификация гранулем
В зависимости от того, какие клетки в туберкулезной гранулеме преобладают, различают такие виды гранулем:
Отметим, что у туберкулезных бугорков клеточный состав непостоянен. Он меняется под воздействием реактивности составляющих их макроорганизмов, работы иммунной системы и получаемого лечения. Замечено, что антибактериальная терапия способствует увеличению в гранулемах гигантских клеток.
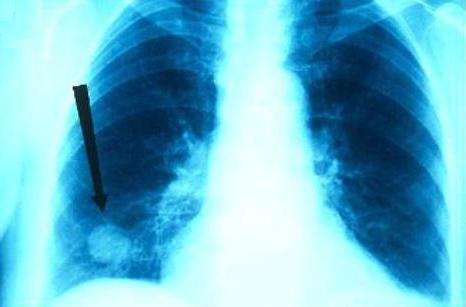
Диагностика
Туберкулезные гранулемы обнаружить при простом клиническом осмотре невозможно. Об их наличии в легких большинство пациентов даже не подозревают, и лишь у небольшой части инфицированных палочками Коха появляются кашель, не излечиваемый обычными противокашлевыми лекарствами, одышка, необъяснимо высокая утомляемость, головные боли. Выявляются туберкулезные узелки при флюорографии или рентгене. В вынесении окончательного вердикта фтизиатра большую роль играют диагностические клетки, входящие в состав туберкулезной гранулемы. Так, эпителиоидные клетки, окрашенные эозином и гематоксилином, при исследовании их световым микроскопом демонстрируют обильную розовую мелкогранулированную цитоплазму. В ней почти всегда обнаруживаются целые или фрагментарные палочки Коха.
Гигантские клетки образуются не только в туберкулезных бугорках, но и в гранулемах, имеющих другую этиологию. Однако при окрашивании их подогретым карболовым фуксином (метод Циля-Нильсена) присутствующие в них палочки Коха становятся малиново-красными на голубом фоне препарата. Это помогает установить точный диагноз.
Варианты развития
Исход туберкулезной гранулемы может быть различным, что зависит от способности организма человека сопротивляться инфекционным заболеваниям. Если резистентность низкая, развитие гранулемы прогрессирует, казеозный некроз развивается и увеличивается в объеме, гранулемы сливаются, образуя обширные зоны поражения, а воспалительный процесс захватывает новые зоны – паренхиму, кровеносные и лимфатические сосуды.
Если резистентность высокая, происходит затухание (регрессирование) процесса. При этом эпителиоидные клетки трансформируются в фибробласты, аргирофильные волокна заменяются коллагеновыми, а гранулема рубцуется. Очаг некроза может подвергнуться кальцинированию, окостенению либо рассосаться.
Гранулематозное воспаление — вариант продуктивного воспаления, при котором доминирующим типом клеток являются активированные макрофаги (или их производные), а основным морфологическим субстратом — гранулема.
Гранулема,или узелок(бугорок, по Р.Вирхову), — это очаговое скопление способных к фагоцитозу клеток моноцитарно-макрофагальной природы. Основным представителем клеток СМФ является макрофаг, который, как уже упоминалось, образуется из моноцита. На "поле" воспаления моноцит делится лишь один раз, а затем трансформируется, как показал опыт с культурой ткани, в макрофаг. Но на этом трансформации не заканчиваются. Через 7 дней после возникновения и размножения макрофаг превращается в эпителиоидную клетку. Для этого необходимы продукты активированных Т-лимфоцитов, особенно 7-интерферон. Эпителиоидные клетки по сравнению с макрофагами имеют более низкую фагоцитарную способность (у них отсутствуют вторичные лизосомы и макрофагальные гранулы), но лучше развитую бактерицидную и секреторную активность — они синтезируют факторы роста (ФРФ, ТФР), фиброне-ктин-1, ИЛ-1. На второй неделе эпителиоидные клетки трансформируются путем деления ядер без деления клетки (реже путем слияния между собой) в гигантские многоядерные клетки Пирогова — Лангханса, а через 2—3 нед — в гигантские клетки инородных тел.
Особенностями гигантских клеток Пирогова — Лангханса являются крупные размеры (до 40—50 мкм), наличие большого (до 20) количества ядер, которые располагаются эксцентрично с одной стороны в форме подковы. В гигантской клетке инородных тел ядер еще больше — до 30 (описывают даже до 100), но они располагаются преимущественно в центре клетки. Оба типа гигантских клеток отличает отсутствие лизосом, поэтому, захватывая различные патогенные факторы, гигантские клетки не в состоянии их переварить, т.е. фагоцитоз в них подменяется эндоцитобиозом. В случаях микробной инвазии эндоцитобиоз поддерживается наличием в цитоплазме секреторных гранул, например л ипидных включений при туберкулезе. Однако в основном секреторная функция их резко подавлена, факторы роста и цитокины, в частности, вообще не синтезируются.
Морфогенез гранулемы складывается из следующих четырех стадий:
▲ накопление в очаге повреждения ткани юных моноцитарных фагоцитов;
▲ созревание этих клеток в макрофаги и образование макрофагальной гранулемы;
▲ созревание и трансформация моноцитарных фагоцитов и макрофагов в эпителиоидные клетки и образование эпителиоидно-клеточной гранулемы;
▲ трансформация эпителиоидных клеток в гигантские (Пирогова — Лангханса и/или инородных тел) и формирование гигантоклеточных гранулем.
Таким образом, учитывая преобладающий клеточный состав гранулемы, по морфологическим признакам различают три вида гранулем: 1) макрофагальную гранулему (простую гранулему, или фагоцитому); 2) эпителиоидно-клеточную гранулему; 3) гигантоклеточную гранулему.
Этиологиягранулематоза. Различают эндогенные и экзогенные этиологические факторы развития гранулем. Кэндогенным относят труднорастворимые продукты поврежденных тканей, особенно жировой ткани (мыла), а также продукты нарушенного обмена, такие как ураты. К экзогенным факторам, вызывающим образование гранулем, относят биологические (бактерии, грибы, простейшие, гельминты), органические и неорганические вещества (пыли, дымы и т.п.), в том числе лекарственные.
В настоящее время гранулемы по этиологии разделяют на две группы: 1) гранулемы установленной этиологии и 2) гранулемы неустановленной этиологии [Струков А.И., Кауфман О.Я., 1989]. Первую группу в свою очередь подразделяют на две подгруппы: инфекционные и неинфекционные гранулемы.
К инфекционным относят гранулемы при сыпном брюшном тифах, бешенстве, вирусном энцефалите, актиномикозе, шистосомозе, туберкулезе, лепре, сифилисе и др.
Неинфекционные гранулемы развиваются при попадании в организм органической и неорганической пыли шерсть, мука, оксид кремния (IV), асбест и др., инородных тел медикаментозных воздействиях (гранулематозный гепатит, олео гранулематозная болезнь).
К гранулемам неустановленной этиологии относят гранулемы при саркоидозе, болезни Крона, первичном билиарном циррозе и др.
Патогенез гранулематоза.Далеко не полный перечень этиологических факторов выявляет совершенно очевидную закономерность — гранулематозное воспаление протекает, как правило, хронически и развивается при следующих двух условиях: 1 наличие веществ, способных стимулировать СМФ, созревание трансформацию макрофагов; 2) стойкость раздражителя по отношению к фагоцитам. Такой раздражитель в условиях незавершенного фагоцитоза и измененной реактивности организма оказывается сильнейшим антигенным стимулятором для макрофага и Т- и В-лимфоцитов. Активированный макрофаг с помощью ИЛ-1 еще в большей степени привлекает лимфоциты, способствуя их активации и пролиферации, — завязываются механизмы клеточно-опосредованного иммунитета, в частности механизмы ГЗТ (подробнее см. лекцию 17 "Реакции гиперчувствительности")- В этих случаях говорят об иммунной гранулеме.
Иммунные гранулемы чаще построены по типу эпителиоидно-клеточных узелков, но в них всегда имеется примесь довольно большого количества лимфоцитов и плазматических клеток. Развиваются они прежде всего при таких инфекциях, как туберкулез, лепра, сифилис, склерома. Иногда продукты тканевого повреждения становятся источником антигенного раздражения и в этих случаях могут подключаться аутоиммунные механизмы гранулемообразования. Наконец, гранулемы, вызванные органическими частицам пыли и аэрозолями, содержащими белки птиц, рыб, шерсть животных, как правило, по механизму их развития являются также антигенно-опосредованными. Хотя иногда возникают механизмы образования гранулем, опосредованные антителами.
К числу неиммунных гранулем относится большинство гранулем, развивающихся вокруг инородных тел, состоящих, прежде всего из частиц органической пыли (например, оксид бериллия (II) является соединением, вызывающим иммунные гранулемы саркоидного типа). Фагоцитоз в клетках неиммунных гранулем более совершенен, и построены они чаще по типу фагоцитомы либо гигантоклеточной гранулемы, состоящей из клеток инородных тел. При сравнении этих гранулем с иммунными отмечается меньшее количество лимфоцитов и плазматических клеток.
К числу критериев в оценке гранулем относят показатель клеточной кинетики, т.е. степени быстроты обмена (обновления) клеток внутри гранулемы, на основании которого выделяют быстро и медленно обновляющиеся гранулемы. Быстро обновляющиеся (за 1—2 нед) гранулемы продуцируют очень токсичные вещества (микобактерии туберкулеза, лепры), построены в основном по типу эпителиоидно-клеточных, характеризуются тем, что их клетки быстро погибают и заменяются новыми, а чужеродный материал лишь частично располагается в макрофагах — все это свидетельствует об интенсивности клеточного обновления. В медленно обновляющихся гранулемах патогенный агент целиком располагается в макрофагах, при этом кинетика обмена резко замедлена. Такие гранулемы возникают при воздействии инертными малотоксичными веществами
и построены чаще всего из гигантских клеток. Этот критерий важен для сравнения гранулем вокруг инородных тел экзогенного и эндогенного происхождения (шовный материал, места татуировок, неорганические пылевые частицы).
Некоторые гранулемы инфекционной этиологии обладают относительной морфологической специфичностью. Для подтверждения диагноза необходима идентификация возбудителя. Специфическими называют те гранулемы, которые вызываются специфическими возбудителями (микобактерии туберкулеза, лепры, бледная трепонема и палочка склеромы), характеризуются относительно специфичными морфологическими проявлениями (только для этих возбудителей и ни для каких других), причем клеточный состав, а иногда и расположение клеток внутри гранулем (например, при туберкулезе) также довольно специфичны.
Гранулемы всех четырех типов возникают при заболеваниях, которые носят хронический, причем волнообразный, характер течения, т.е. с периодами обострений и ремиссий. Как правило, при всех этих заболеваниях развивается особый вид некроза — казеозный некроз.
Туберкулезная гранулема имеет следующее строение: в центре ее расположен очаг казеозного некроза, за которым находится вал радиарно расположенных (вытянутых по длиннику от центра к периферии) эпителиоидных клеток; за ними видны единичные гигантские клетки Пирогова — Лангханса и, наконец, на периферии гранулемы располагается еще один вал лимфоидных клеток. Среди этих типичных клеток может быть примесь небольшого количества плазматических клеток и макрофагов. При импрегнации солями серебра среди клеток гранулемы обнаруживают тонкую сеть аргирофильных (ретикулярных) волокон. Кровеносные сосуды обычно в туберкулезной гранулеме не встречаются. При окраске по Цилю — Нильсену в гигантских клетках выявляют микобактерии туберкулеза.
Учитывая преобладание в описанной выше гранулеме эпителиоидных клеток, такую гранулему называют эпителиоидно-клеточной. Эта специфическая гранулема является иллюстрацией типичной инфекционной (по этиологии), иммунной (по патогенезу), эпителиоидно-клеточной (по морфологии) гранулемы.
Обычно туберкулезные гранулемы невелики — их диаметр не превышает 1—2 мм, чаще они обнаруживаются лишь микроскопически. Однако и макроскопически изменения довольно типичны — многочисленные сливающиеся гранулемы внешне напоминают мелкие, как просо, бугорки, отчего процесс принято называть милиарным (от лат. miliarius - просяной) туберкулезом.
Сифилитическая гранулема носит название "гумма" (от лат. gummi — камедь). Она, как и туберкулезная гранулема, в центре представлена очагом казеозного некроза, но гораздо более крупным по размерам. По периферии от некроза располагается множество лимфоцитов, плазматических клеток и фибробластов. Эти три типа клеток являются преобладающими, но в небольшом количестве в гумме могут встречаться эпителиоидные клетки, макрофаги и единичные гигантские клетки типа Пирогова — Лангханса. Для сифилитической гранулемы характерно быстрое в связи с пролиферацией фибробластов разрастание массивной плотной соединительной ткани, которая формирует подобие капсулы. С внутренней стороны этой капсулы среди клеток инфильтрата видны многочисленные мелкие, а снаружи — более крупные сосуды с явлениями продуктивного эндоваскулита. Крайне редко среди клеток инфильтрата удается выявить бледную трепонему путем серебрения по Левадити.
Гумма характерна для третичного периода сифилиса, который обычно развивается через несколько лет (4—5 и позже) после заражения и длится десятилетиями. При этом в разных органах — костях, коже, печени, головном мозге и др., — появляются солитарные (от лат. solitarius — склонный к одиночеству) узлы размером от 0,3—1,0 см на коже и размером до куриного яйца — во внутренних органах. При разрезе из этих узлов выделяется желеобразная масса желтого цвета, напоминающая клей гуммиарабик (аравийскую камедь), откуда и возникло наименование сифилитической гранулемы.
Помимо гумм, в третичном периоде сифилиса может развиваться гуммозная инфильтрация. Гуммозный инфильтрат обычно представлен теми же клетками, которые являются доминирующими и в гумме, т.е. лимфоцитами, плазмоцитами и фибробластами. При этом очень быстро выявляется тенденция к склерозу — разрастается грануляционная ткань. Среди клеток инфильтрата выявляется множество мелких, капиллярного типа сосудов: в этих сосудах также обнаруживают продуктивный васкулит. Подобные изменения чаще всего развиваются в восходящей части и в дуге грудного отдела аорты и носят название сифилитического мезаортита. Расположенный в средней и наружной оболочках аорты гуммозный инфильтрат вместе с пораженными vasa vasorum разрушает эластический каркас аорты — при окраске фукселином на месте бывших эластических волокон появляются своеобразные "плеши". На месте эластических волокон разрастается соединительная ткань. Именно в этих участках бывшего гуммозного инфильтрата внутренняя оболочка аорты становится неровной, морщинистой, со множеством Рубцовых втяжений и выбуханий, напоминая "шагреневую кожу". Малоэластичная стенка аорты в очагах поражения под давлением крови истончается, выбухает наружу и формируется аневризма грудного отдела аорты. Если гуммозный инфильтрат с аорты "спускается" на ее клапаны, то формируется аортальный порок сердца.
Диффузная гуммозная инфильтрация в печени имеет аналогичное строение и приводит к развитию дольчатой печени в связи со сморщиванием соединительной ткани, разрастающейся на месте специфического поражения. Аналогичные изменения на коже и слизистых оболочках иногда приводят к резкому обезображиванию лица — язвы, рубцы, разрушение носовой перегородки и пр.
Лепрозная гранулема (лепрома) имеет полиморфный клеточный состав: в ней видны в большом количестве макрофаги, эпителиоидные клетки, а также гигантские, плазматические клетки, фибробласты. Микобактерии Гансена — Нейссера в огромном количестве содержатся в макрофагах (установлено, что 1 г "цветущей" лепромы содержит 5 • 10 9 лепроматозных микобактерии). Последние, переполненные возбудителями, увеличиваются, как бы разбухают, в их цитоплазме появляются жировые включения. Такие макрофаги, называемые лепрозными клетками Вирхова, переполнены микобактериями, которые лежат в них строго упорядоченными рядами, напоминая сигареты в пачке, что особенно хорошо видно при окраске по Цилю — Нильсену. В последующем микобактерии, склеиваясь, образуют лепрозные шары. Макрофаг со временем разрушается, выпавшие лепрозные шары фагоцитируются гигантскими клетками инородных тел. Наличие в лепроме огромного количества микобактерии обусловлено незавершенным фагоцитозом в макрофагах при проказе.
Тканевые реакции при проказе находятся в тесной связи с резистентностью организма, которая целиком зависит от его взаимоотношения с микобактериями лепры и определяет все разнообразие клинических проявлений болезни. Различают несколько вариантов течения проказы, но наиболее ярко выяглядят две "крайние" клинико-анатомические формы: 1) с высокой резистентностью — туберкулоидная; 2) с низкой резистентностью — лепроматозная.
Туберкулоидная форма протекает клинически доброкачественно, иногда с самоизлечением, на фоне выраженного клеточного иммунитета. Поражение кожи диффузное, со множеством пятен, бляшек и папул, с последующей депигментацией пораженных участков. Морфологически обнаруживают эпителиоидно-клеточные гранулемы, а микобактерии выявляют в редких случаях. Все это является подтверждением развития лепромы по типу ГЗТ. Изменение нервов характеризуется диффузной инфильтрацией их эпителиоидными клетками, что проявляется ранними
нарушениями чувствительности. Изменения внутренних органов для этой формы нехарактерны.
Лепроматозная форма является полной противоположностью туберкулоидной формы. Поражение кожи нередко носит диффузный характер, вовлекаются, а затем полностью разрушаются придатки кожи — потовые и сальные железы, повреждаются сосуды. В лепроме обнаруживаются макрофаги, гигантские клетки и множество микобактерии. Диффузная инфильтрация кожи лица иногда приводит к полному обезображиванию внешности ("львиная морда"). Лепрозный неврит носит восходящий характер, развивается диффузная инфильтрация всех элементов чувствительных нервов макрофагами с постепенным замещением нервного волокна соединительной тканью. Гранулемы из макрофагов с высоким содержанием микобактерии обнаруживают в печени, селезенке, костном мозге, лимфатических узлах, слизистой оболочке верхних дыхательных путей, в эндокринных органах. Все перечисленное может быть свидетельством значительного угнетения клеточных реакций иммунитета при лепроматозной форме лепры, при этом отмечают выраженную дисфункцию гуморального звена.
Склеромная гранулема характеризуется скоплением макрофагов, лимфоцитов, большого числа плазматических клеток и продуктов их деградации — эозинофильных телец Русселя. Специфическими для склеромной гранулемы являются очень крупные одноядерные клетки с вакуолизированной цитоплазмой — клетки Микулича. Макрофаги интенсивно захватывают диплобациллы, однако фагоцитоз в них незавершенный. Часть макрофагов разрушается, а часть, укрупняясь, превращается в клетки Микулича, в которых и находят возбудителя склеромы — палочку Волковича - Фриша.
Склеромная гранулема обычно располагается в слизистой оболочке верхних дыхательных путей — носа, гортани, трахеи, реже — бронхов. Процесс заканчивается образованием на месте гранулем грубой рубцовой ткани, в результате слизистая оболочка деформируется, дыхательные пути резко суживаются и даже иногда полностью закрываются, вызывая опасность асфиксии.
Исходы гранулем.Возможны следующие исходы гранулема-тозного воспаления:
▲рассасывание клеточного инфильтрата. Это редкий вариант исхода, так как гранулематоз в основной массе является хроническим воспалением. Подобное возможно только в случаях малой токсичности патогенного фактора и быстрой элиминации его из организма. Примером служат такие острые инфекции, как бешенство, брюшной и сыпной тиф.
▲ фиброзное превращение гранулемы с образованием рубца или фиброзного узелка. Это наиболее частый и типичный вариант исхода гранулематоза. Развитие склероза стимулирует ИЛ-1, выделяемый макрофагами гранулемы, а нередко и сам патогенный агент.
▲ некроз гранулемы. Этот исход характерен прежде всего для туберкулезной гранулемы, которая может целиком подвергнуться казеозному некрозу, и еще для ряда инфекционных гранулем. В развитии некроза во всех случаях участвуют протеолитические ферменты макрофага, а также продукты, выделяемые патогенным агентом, которые обладают прямым токсическим действием на ткани. Эксперимент с гранулемами, развившимися после введения БЦЖ, позволил доказать антительный механизм некроза, при этом иммунные комплексы были обнаружены в стенках сосудов, где развивалась картина продуктивного васкулита. В случае некроза гранулем, расположенных на коже, слизистых оболочках, неизбежно происходит расплавление тканей с образованием язв.
▲ нагноение гранулем. Это встречается, как правило, при грибковых поражениях. При многих инфекциях (сап, иерсиниоз, туляремия) и грибковых поражениях на первых этапах появляется много нейтрофилов, но только в случае микотического поражения они не справляются с возбудителем, гибнут, а продукты их гибели, будучи хемоаттрактантами, привлекают макрофаги. Так возникают своеобразные гранулемы с абсцессом в центре.
Дата добавления: 2015-08-14 ; просмотров: 6289 ; ЗАКАЗАТЬ НАПИСАНИЕ РАБОТЫ
Гигантские клетки нередко образуются при продуктивном воспалении (см.), являясь характерной структурной особенностью многих специфических гранулем, при регенерации (см.), инфекционных процессах вирусной этиологии, в опухолях, в частности — костных (остеобластокластома), при современных методах лечения цитостатическими препаратами, рентгеновскими лучами. При ряде заболеваний (напр., при анемиях различного происхождения, при реактивных эритробластозах, остеомиелосклерозе) в очагах экстрамедуллярного кроветворения встречаются мегакариоциты. При воспалении, регенерации, нарушениях обмена, напр, при остеомиелитах, рахите, системной фиброзной остеодистрофии, костных переломах, встречаются остеокласты. При патологических процессах встречаются преимущественно два морфологических типа Гигантских клеток: инородных тел — некрофаги Подвысоцкого (1889) и Г. к. типа Пирогова—Лангханса (Th. Langhans, 1868).

В Г. к. инородных тел ядра разбросаны группами или расположены в центре клетки; количество их может колебаться в пределах от 3—5 до 100 и более; они светлые, овальные, могут быть и гиперхромными. Цитоплазма клетки гомогенная. Г. к. инородных тел образуются при попадании в организм труднорастворимых инородных тел, напр, кусочков дерева, стекла, обрывков операционного шелка, волокон ваты, нерастворимых газов (напр., при пневматозе кишечника). Иногда они образуются в ответ на скопление труднорастворимых продуктов, появляющихся в результате каких-либо патол. процессов в самом организме, напр, частицы эктопического или избыточного рогового вещества (цветн. рис. 7), хряща, кристаллов жирных кислот, продуктов некроза жировой клетчатки, в атероматозных бляшках при атеросклерозе, при рассасывании амилоида, при распаде эластической ткани артерий и т. д.

Гигантские клетки типа Пирогова—Лангханса характеризуются периферическим расположением овальных ядер; цитоплазма их делится на эндо- и эктоплазму. Характерным считается наличие клеточного центра с многочисленными центриолями, которые соединены между собой центродесмозами, образующими своеобразную сеть внутри клетки; в этих Г. к. также обнаруживается большое количество митохондрий (рис.), канальцы эндоплазматического ретикулума иногда заполнены жировыми вакуолями. В ядре отмечается увеличение размеров или количества ядрышек. Г. к. типа Пирогова—Лангханса могут достигать 300 мкм в диаметре, количество ядер в них ок. 200. Наличие Г. к. типа Пирогова—Лангханса является характерным для туберкулезного продуктивного воспаления; в их цитоплазме можно видеть фагоцитированные кислотоустойчивые туберкулезные микобактерии, хорошо окрашивающиеся по Циля — Нельсена методу (см.). Наличие микобактерий имеет решающее значение для диагноза, т. к. Г. к. типа Пирогова—Лангханса могут образоваться и при других инфекционных и неинфекционных воспалительных процессах, напр, при бруцеллезе, сифилисе, проказе (клетки Вирхова), микозах, склероме (клетки Микулича), симпатической офтальмии, болезни кошачьей царапины, саркоидозе, липогранулематозе и др.
При вирусных инфекциях, помимо Г. к. типа Пирогова Лангханса и Г. к. инородных тел, обнаруживают Г. к. эпителиального происхождения, что зависит от тропизма многих вирусов к эпителиальным клеткам. Образование большого количества Г. к. при вирусных инфекциях наблюдается преимущественно в детском возрасте.

Многоядерные Г. к. с причудливым расположением ядер (чаще гиперхромных) обнаруживают при кори в конъюнктиве глаза, в эпителии бронхов, бронхиол, среди альвеолярных клеток; они образуют так наз. гигантоклеточные пневмонии (цветн. рис. 8). Г. к. макрофагального происхождения находят в миндалинах, в лимфатическом аппарате червеобразного отростка в продромальном периоде кори. При ветряной оспе Г. к. образуются в эпителии кожи на ранних стадиях формирования ветряночной везикулы. При вирусном гепатите среди печеночного эпителия образование много-ядерных Г. к. свидетельствует о начинающихся процессах регенерации.

Диагностический интерес представляет обнаружение так наз. цитомегалических клеток размером 20—30 мкм при вирусном поражении слюнных желез (цветн. рис. 9). Это одноядерные Г. к., характеризующиеся крупным внутриядерным включением, к-рое связывают с наличием самого вируса. При генерализованной цитомегалии Г. к. наблюдаются в эпителии слюнных желез, в легких, печени, почках, поджелудочной железе, надпочечниках и т. д.; возникают они преимущественно из эпителиальных клеток. Однако их можно наблюдать и среди клеток нервной и мезенхимальной ткани (головной мозг, тимус, лимф. узлы).
При лимфогранулематозе наблюдается образование одноядерных и многоядерных Г. к. из макрофагов. Образование Г. к. соединительнотканного, эпителиального, неврогенного и мышечного происхождения часто наблюдается в опухолях. Морфология этих Г. к. разнообразна, гистогенез связан с гистогенезом самих опухолей.
Гигантские клетки были обнаружены в культурах тканей. Так, А. Д. Тимофеевский в 1914 г. в культуре лейкемической крови наблюдал образование Г. к. из гематогенных макрофагов вокруг случайно попавших инородных тел.
При введении в эту культуру тканей туберкулезных бацилл были получены типичные Г. к. типа Пирогова—Лангханса.
В редких случаях наблюдается образование своеобразных миогенных Г. к. в миокарде при патологических процессах, сопровождающихся гибелью мышечных волокон (дистрофии, расстройства кровообращения, воспалительные процессы).
Гистогенез. Г. к. чаще всего возникают из гистио- и гематогенных макрофагов (см.) и из эпителия;
значительно реже они образуются из мышечной и нервной ткани. Г. к. эпителиального происхождения в норме иногда встречаются в эпителии мочевого пузыря. Они бывают двухядерными или многоядерными, причем в некоторых случаях из переходного эпителия мочевого пузыря (при развитии рака) появляется большое количество атипических Г. к. При болезнях накопления (энзимопатиях) образуются Г. к. неврогенного гистогенеза (см. Ниманна — Пика болезнь). Особого рода глобоидные многоядерные Г.к. патогномоничны для болезни Краббе, характеризующейся недостаточностью энзима галактоцереброзидазы (см. Лейкодистрофии); их происхождение возможно из гистиоцитов, т. к. эти Г.к. располагаются преимущественно вокруг мелких сосудов.
Условно к Г. к. неврогенного происхождения могут быть отнесены подошвы двигательных бляшек, которые представляют собой цитоплазменные образования, содержащие несколько ядер двух типов — более вытянутые, напоминающие ядра мышечных волокон, и округлые, напоминающие ядра клеток шванновской оболочки.
Гигантские клетки мезенхимального происхождения встречаются в норме у человека в костном мозге; к ним относят мегакариоциты и остеокласты. Остеокласты возникают из камбиальных элементов надкостницы; их образование регулируется гормоном околощитовидной железы. Они в большом количестве обнаруживаются в растущих костях.
Г. к. типа Пирогова—Лангханса и Г. к. типа инородных тел образуются из эпителиоидных клеток (гистиогенных макрофагов).
Причины образования Г. к. типа Пирогова—Лангханса и Г. к. инородных тел еще недостаточно выяснены. Имеются наблюдения, что некоторые хим. фракции туберкулезных палочек (А3-фосфатид, фосфорная к-та), особенно в сочетании со смесью жирных кислот, дают в эксперименте образование особенно большого количества Г. к. Высказывается точка зрения, что образование Г. к. типа Пирогова—Лангханса обратно пропорционально вирулентности самого возбудителя: при введении маловирулентной культуры у экспериментальных животных образуется большое количество Г. к. и малое число Г. к. при введении вирулентной культуры.
При лечении антибиотиками, когда падает вирулентность туберкулезной палочки, отмечается повышенное образование Г. к. типа Пирогова—Лангханса. Эти клетки с очень высокой ферментативной активностью и большим содержанием функциональных групп белка. Однако локализация и интенсивность реакций на белки и ферменты зависит от степени зрелости Г. к. Пирогова—Лангханса. Увеличение объема ядрышек в этих клетках является свидетельством медленного синтеза РНК, увеличение числа митохондрий свидетельствует о высоком уровне окислительных процессов. Известно, что Г. к. инородных тел и Г. к. типа Пирогова—Лангханса не возникают в условиях острого воспаления. Их появление связано с пролиферативными длительно текущими воспалительными процессами.
Причины образования Г. к. при вирусных инфекциях также изучены недостаточно. Частота обнаружения их в детском возрасте связана, по-видимому, с высокой пролиферативной способностью тканей именно в этом возрастном периоде. С другой стороны, на возникновение Г. к., очевидно, оказывают влияние сами вирусы, а большинство вирусных инфекций возникает именно в детском возрасте.
Механизм образования Г. к. трактуется различными авторами по-разному. Существует мнение, что они образуются синцитиальным путем, т. е. в результате слияния одноядерных клеточных форм; некоторые авторы считают, что они образуются пролиферативным путем, т. е. благодаря делению ядра с одновременным ростом цитоплазмы. Ряд авторов признает возможность образования Г. к. и тем, и другим способом.
Установлено, что оба способа образования Г. к. имеют место как в живом организме, так и в культуре тканей. Так, напр., образование Г. к. в культуре тканей из макрофагов идет путем слияния, образование Г. к. из эпителия и эндотелия сосудов — путем амитотического деления ядер и роста цитоплазмы, не сопровождающегося плазмотомией. В организме, напр., Г. к. типа Пирогова—Лангханса при туберкулезе образуются путем амитотического деления ядра с последующим ростом цитоплазмы; из печеночного эпителия при вирусном гепатите Г. к. образуются путем слияния клеточных тел.
Функциональное значение. Значение и роль Г. к. как активных фагоцитов впервые была выявлена И. И. Мечниковым. Все последующие наблюдения подтвердили фагоцитарную способность многих Г. к. Наличие Г. к. при инфекционных заболеваниях расценивается как проявление иммунной реакции клеточного типа; их образование зависит от высокой потребности организма в фагоцитарной деятельности. При этом фагоцитоз (см.) в Г. к. часто носит незавершенный характер (эндоцитобиоз), от к-рого зависит нестерильный иммунитет, напр, при туберкулезе. Очевидно, такого же характера роль Г. к. при вирусных инфекциях, особенно когда в Г. к. обнаруживаются вирусные включения (напр., при ветряной оспе, цитомегалии). Надо полагать, что здесь имеет место своеобразная форма внутриклеточного существования вируса.
Образование Г. к. в лейкемических инфильтратах при лечении цитостатическими средствами и рентгеновскими лучами, в частности при хроническом миелолейкозе, не имеет пока должного объяснения, предполагают, что здесь играет роль проявление клеточного иммунного ответа.
Г. к. могут фагоцитировать не только микробы, но всевозможные продукты распада и обмена (белкового, липоидного, минерального, пигментного) в организме. А. В. Немилов обнаружил Г. к. у животных и человека в селезенке и в лимфатических узлах, где они возникали в старческом возрасте в ответ на появление продуктов распада. Следовательно, Г. к. играют в организме роль не только активных фагоцитов при разного рода инфекционных процессах, но принимают активное участие в обменных процессах.
Образование Гигантских клеток имеет значение при регенерации тканей, о чем свидетельствует появление их при регенерации скелетной мускулатуры, печеночной ткани, при проявлениях регенераторных процессов со стороны мышечных волокон миокарда.
Библиография: Ван Фюрт Р. и др. Система мононуклеарных фагоцитов, новая классификация макрофагов, моноцитов и их клеток предшественников, Бюлл. ВОЗ, т. 46, № 6, с. 814, 1973, библиогр.; Гистология, под ред. В. Г. Елисеева и др., М., 1972; Подвысоцкий В. В. О разъедании омертвевших участков печеночной ткани гигантскими клетками гепатофагами, Врач, № 3, с. 1,1889; Струков А. И. Патологическая анатомия, М., 1971; Шройт И. Г. Экспериментальная корь, патоморфология и патогенез, Кишинев, 1961, библиогр.; Fаllis В. D. Textbook of pathology, N. Y., 1964; Langhan s T. Uber Riesenzellen mit wand-standigen Kernen in Tuberkeln und die fibrose Form des Tuberkels, Virchows Arch. path. Anat., Bd 42, S. 382, 1868.
Читайте также: