
Что такое адсорбция вируса
начальный этап вирусной инфекции, заключающийся в прикреплении вируса к поверхности клетки.
Адсорбция Ж. — 1. Поглощение, всасывание вещества из раствора или газа поверхностью твердого тела или поверхностным слоем жидкости.
Толковый словарь Ефремовой
Адсорбция — -и; ж. [от лат. ad- - к и sorbere - поглощать, всасывать]. Поглощение вещества из газа или раствора поверхностным слоем жидкого или твёрдого адсорбента (используется в химии.
Толковый словарь Кузнецова
Адсорбция — (ад- + лат. sorbeo поглощать) поглощение (концентрирование) газов или растворенных веществ на поверхности твердого тела или жидкости.
Большой медицинский словарь
Адсорбция Бактериофага — прикрепление бактериофага к поверхности бактериальной клетки, наступающее при наличии у них специфических рецепторов.
Большой медицинский словарь
Адсорбция Вируса — начальный этап вирусной инфекции, заключающийся в прикреплении вируса к поверхности клетки.
Большой медицинский словарь
Адсорбция — поглощение вещества из раствора или газа поверхностным слоем жидкости или твердого тела (адсорбентом); играет важную роль в биол. системах, широко применяется в биохимии.
Словарь микробиологии
Адсорбция Вируса — первая фаза взаимодействия вируса с клеткой. Характеризуется выраженной специфичностью, определяемой соответствием рецепторов клеточной стенки (находятся в липопротеиновом.
Словарь микробиологии
Дезинтеграция Вируса — распад вириона на составные части, наступающий в процессе вирусной инфекции клетки (см. Депротеинизация) или под действием физ. факторов, противомикробных веществ, прежде всего ПАВ.
Словарь микробиологии
Инфекционная Единица Вируса — наименьшее количество вирусных частиц (вирионов), способное вызывать у чувствительного организма проявление инфекции (заболевание или локальное поражение).
Словарь микробиологии
Криптограмма Вируса — запись структуры и свойств вируса в виде четырех пар символов–1–я пара символов – тип нуклеиновой кислоты (R – РНК, D – ДНК) и число цепей нуклеиновой кислоты (1, 2); 2–я.
Словарь микробиологии
Титр Вируса — 1) количество вирусов в ед. объема (обычно в 1 мл) суспензии. Подсчитывают в электронном микроскопе или методом бляшек (см. Бляшки) на к-ре клеток. В первом случае выявляют.
Словарь микробиологии
Граффи Штамм Вируса — (A. Graffi, род. в 1910 г., нем. патолог) штамм вируса миелоидной хлорлейкемии мышей, используемый в эксперименте; у новорожденных мышей вызывает миелоидные лейкозы и гемоцитобластозы.
Большой медицинский словарь
Адсорбция — , притяжение газа или жидкости к поверхности твердого тела или жидкости, в отличие от абсорбции, при которой подразумевается проникновение одного вещества в другое.
Научно-технический энциклопедический словарь
Адсорбция — (от лат. ad - на - при и sorbeo - поглощаю), поглощение газов,паров или жидкостей поверхностным слоем твердого тела (адсорбента) илижидкости. Адсорбенты обычно имеют большую.
Большой энциклопедический словарь
Карра-зильбера Штамм Вируса — (J. G. Carr; Л. А. Зильбер, 1894-1966, сов. микробиолог) штамм вируса саркомы Рауса, вызывающий опухоли не только у птиц, но и у млекопитающих.
Большой медицинский словарь
Оболочка Вируса — белковая структура, окружающая нуклеиновую кислоту вириона или нуклеокапсид; в состав О. в. могут входить также липиды и углеводы.
Большой медицинский словарь
Рича Штамм Вируса — (М. А. Rich) штамм вируса лейкоза мышей, вызывающий у них лимфолейкозы, иногда миелолейкозы.
Большой медицинский словарь
Субъединица Вируса — (син. структурная единица вируса) структура, состоящая из одного или нескольких полипептидов вируса.
Большой медицинский словарь
Титр Вируса — количество инфекционных, бляшкообразующих, гемагглютинирующих или других единиц активности вируса, определяемое в единице объема вируссодержащей жидкости.
Большой медицинский словарь
Криптограмма Вируса — запись структуры и свойств вируса в виде четырёх пар символов. 1-я пара символов — тип нуклеиновой к-ты (R — РНК, D — ДНК) и число цепей нуклеиновой к-ты (1,2); 2-я пара — мол.
Биологический энциклопедический словарь
Адсорбция — (от лат. ad - y, на, при и sorbeo - поглощаю * a. adsorption; н. Adsorption, Adsorbieren, Adsorbierung; ф. adsorption; и. adsorcion) - поглощение отд. компонентов из газовых (паровых) или жидких смесей на поверхности.
Горная энциклопедия
Адсорбция — - поглощение вещества из газа или раствора поверхностью раздела между ними (или поверхностью твердого тела). Иными словами, А. есть поглощение адсорбата из объема фаз.
Математическая энциклопедия
Адсорбция — поглощение вещества из раствора или газа поверхностным слоем жидкости или твердого тела. Используется в очистке сточных вод и газовых выбросов.
Экологический словарь
Цикл репродукции вирусов при продуктивной инфекции представляет собой специфическую последовательность событий, приводящих к образованию нового потомства вирионов. Его можно разделить на несколько стадий: адсорбция вирионов на поверхности клетки, проникновение вируса или вирусного нуклеопротеида внутрь клетки, депротеинизация генома, синтез вирусных компонентов, формирование и выход из клетки зрелых вирионов. Иногда не все стадии протекают последовательно и полностью даже при продуктивной инфекции. При заражении вирусной нуклеиновой кислотой отсутствует стадия депротеинизации. Цикл репродукции вируса может прерываться на какой-либо промежуточной стадии, и в этом случае инфекционные вирионы не образуются (абортивная инфекция).
Классическое изучение кривой роста вируса в одном цикле репродукции предполагает одновременное инфицирование клеток культуры с использованием высокой множественности заражения (М=10—100) и определение накопления вируса через определенные промежутки времени.
Прикрепление (адсорбция) вирионов к поверхности клетки - первая стадия вирусной инфекции. Для того, чтобы началась инфекция, вирионы должны быть способны прикрепляться к клетке. Безусловно, этот процесс в организме является более сложным, чем в культуре клеток.
Детальное изучение процесса адсорбции вирусов показало, что он состоит из двух быстро следующих друг за другом периодов — обратимого и необратимого. В период обратимой адсорбции вирус можно удалить с поверхности клетки при обработке версеном, хемотрипсином и другими химическими веществами. При необратимой адсорбции вирус удалить с поверхности клетки не удается.
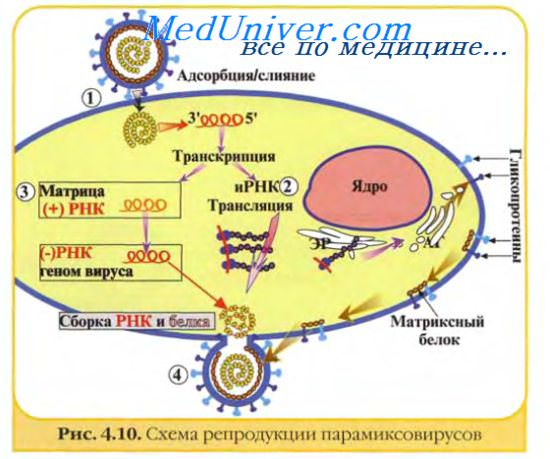
Первичный контакт с клеткой происходит в результате случайных столкновений вирионов с поверхностью клетки. Количество стабильных прикреплений вириона к клетке во много раз меньше количества случайных столкновений.
В основе прикрепления вируса к клетке лежат два механизма — неспецифический и специфический. Первый из них определяется силами электростатического взаимодействия, возникающими между разноименно заряженными группами, расположенными на поверхности клетки и вируса. В этом процессе, прежде всего, могут участвовать положительно заряженные аминные группы вирусного белка и отрицательно заряженные группы клеточной поверхности. В пользу электростатического взаимодействия между клеткой и вирусом свидетельствуют данные о зависимости присоединения вирионов к клеткам от ионной силы раствора и его рН, наличия в среде одно- и двухвалентных катионов и присутствия отрицательно заряженных групп на поверхности клетки.
Наиболее важным механизмом прикрепления вируса является специфическое взаимодействие рецепторов вируса с комплементарными рецепторами клетки.
Прикрепление происходит за счет связи между вирионными прикрепительными белками на поверхности вирионов и рецепторами на плазматической мембране клеток.
На поверхности вирионов многих семейств вирусов (все РНК-содержащие оболочечные вирусы и аденовирусы) имеются пепломеры (выступы и шипы) длиной от 10 до 30 нм, которые принимают непосредственное участие в прикреплении вируса к клетке. Отщепление пепломеров с помощью протеолитических ферментов приводит к потере вирионами способности адсорбироваться на клетках. На поверхности вирионов, не имеющих пепломеров, находятся участки связывания с клеточной поверхностью, которые отличаются сложным строением и, вероятно, состоят из нескольких полипептидов.
Спектр чувствительности клеток к вирусам в значительной мере определяется наличием соответствующих рецепторов. Рецепторный барьер может быть преодолен при заражении инфекционной нуклеиновой кислотой.
На поверхности клетки может находиться большое количество различных видов рецепторов. Полагают, что каждая клетка может адсорбировать от десятков до нескольких тысяч вирионов. На поверхности одной клетки находится около 10000 рецепторов для полиовируса. Каждый рецептор специфичен для одного или нескольких вирусов, как родственных между собой, так и относящихся к различным таксономическим группам. Например, аденовирус типа 2 и вирус Коксаки ВЗ связываются с одними и теми же клеточными рецепторами.
У герпесвирусов прикрепительными белками могут служить гликопротеины оболочки, которые соединяются с некоторыми рецепторами клетки. Клеточным рецептором для многих ортомиксовирусов являются концевые сиаловые кислоты олигосахаридов, входящих в структуру гликопротеринов или гликолипидов, экспонированных на поверхности клетки. Рецепторами для многих риновирусов являются иммуноглобулины. У вируса ВИЧ в прикреплении первоначально участвуют молекулы СД4, расположенные на поверхности клеток, особенно макрофагов и Т-хелперных лимфоцитов. Подобные взаимодействия рецептор-лиганд свойственны другим лентивирусным инфекциям.
Рецепторы для одних вирусов имеются на клетках нескольких или многих тканей, тогда как для других вирусов они обнаружены на клетках одного вида тканей или в один из периодов онтогенеза (новорожденные или взрослые животные).
Рецепторы клеток способны к регенерации после удаления их протеолитическими ферментами.
В.А. Сергеев, Е.А. Непоклонов, Т.И. Алипер, Вирусы и вирусные вакцины, 2007
Прикрепление (адсорбция) вирионов к поверхности клетки - первая стадия вирусной инфекции. Для того, чтобы началась инфекция, вирионы должны быть способны прикрепляться к клетке. Безусловно, этот процесс в организме является более сложным, чем в культуре клеток.
Детальное изучение процесса адсорбции вирусов показало, что он состоит из двух быстро следующих друг за другом периодов - обратимого и необратимого. В период обратимой адсорбции вирус можно удалить с поверхности клетки при обработке версеном, хемотрипсином и другими химическими веществами. При необратимой адсорбции вирус удалить с поверхности клетки не удается.
Первичный контакт с клеткой происходит в результате случайных столкновений вирионов с поверхностью клетки. Количество стабильных прикреплений вириона к клетке во много раз меньше количества случайных столкновений.
В основе прикрепления вируса к клетке лежат два механизма — неспецифический и специфический. Первый из них определяется силами электростатического взаимодействия, возникающими между разноименно заряженными группами, расположенными на поверхности клетки и вируса. В этом процессе, прежде всего, могут участвовать положительно заряженные аминные группы вирусного белка и отрицательно заряженные группы клеточной поверхности. В пользу электростатического взаимодействия между клеткой и вирусом свидетельствуют данные о зависимости присоединения вирионов к клеткам от ионной силы раствора и его pH, наличия в среде одно- и двухвалентных катионов и присутствия отрицательно заряженных групп на поверхности клетки [198].
Наиболее важным механизмом прикрепления вируса является специфическое взаимодействие рецепторов вируса с комплементарными рецепторами клетки.
Прикрепление происходит за счет связи между вирионными прикрепительными белками на поверхности вирионов и рецепторами на плазматической мембране клеток.
На поверхности вирионов многих семейств вирусов (все РНК-содержащие оболочечные вирусы и аденовирусы) имеются пепломеры (выступы и шипы) длиной от 10 до 30 нм, которые принимают непосредственное участие в прикреплении вируса к клетке. Отщепление пепломеров с помощью протеолитиче- ских ферментов приводит к потере вирионами способности адсорбироваться на клетках. На поверхности вирионов, не имеющих пепломеров, находятся участки связывания с клеточной поверхностью, которые отличаются сложным строением и, вероятно, состоят из нескольких полипептидов.
Спектр чувствительности клеток к вирусам в значительной мере определяется наличием соответствующих рецепторов. Рецепторный барьер может быть преодолен при заражении инфекционной нуклеиновой кислотой.
На поверхности клетки может находиться большое количество различных видов рецепторов. Полагают, что каждая клетка может адсорбировать от десятков до нескольких тысяч вирионов. На поверхности одной клетки находится около 10000 рецепторов для полиовируса [169]. Каждый рецептор специфичен для одного или нескольких вирусов, как родственных между собой, так и относящихся к различным таксономическим группам. Например, аденовирус типа 2 и вирус Кок- саки ВЗ связываются с одними и теми же клеточными рецепторами [110, 164].
У герпесвирусов прикрепительными белками могут служить гликопротеины оболочки, которые соединяются с некоторыми рецепторами клетки. Клеточным рецептором для многих ортомиксовирусов являются концевые сиаловые кислоты олигосахаридов, входящих в структуру гликопротеринов или гликолипидов, экспонированных на поверхности клетки. Рецепторами для многих риновирусов являются иммуноглобулины. У вируса ВИЧ в прикреплении первоначально участвуют молекулы СД4, расположенные на поверхности клеток, особенно макрофагов и Т-хелперных лимфоцитов. Подобные взаимодействия рецептор-лиганд свойственны другим лентивирусным инфекциям [663].
Рецепторы для одних вирусов имеются на клетках нескольких или многих тканей, тогда как для других вирусов они обнаружены на клетках одного вида тканей или в один из периодов онтогенеза (новорожденные или взрослые животные).
Рецепторы клеток способны к регенерации после удаления их протеолитиче- скими ферментами [110, 1600].
Адсобция вкл 3 стадии:
-адсорбция на клетке
-проникновение в клетку
-раздевание вируса в клетке
Далее адсорбция стан специф т.к. прикрепительные белки вирусов узнаут клеточные рецепторы.
Вирусы исп-ют рецепторы, предназначенные для прохождения в кл-ку необх для её ж\д вещ-в
Для пикорно-вирусов и аденовирусов это белки, для вир гриппа и парамиксавирусов это сиаловая кислота, входящая в состав гликолипидов и гликопротеидов.
Адсорбция на первых этапах яв-ся обратимой т.к. вир частица обр-ет единичную связь с рецептором, такая адсорбция непрочная;
На 1 вирусную частицу до 3 тыс рецепторов в липидном бислое цпм.
Кол-во специф рецепторов м.б. 10 4 -10 5 на одну клетку. Некоторые представлены в огр наборе, что будет опр-ть чувствительность клеток к данному вирусу.
Для нек вирусов хар-но наличие широкого круга хозяев.
Вирусные прикрепительные белки могут находиться в особых структурах вируса.
У простоустроенных вирусов в составе капсида; у сложноустроенных в составе суперкапсида и представляют собой множественные молекулы.
Проникновение вируса в клетку:
2.слияние вирусной и клеточной мембран.
1 – проходит в спец участках кл мембраны, где имеются спец ямки, покрытые со стороны цитоплазмы белком с большой молекул массой (клатрином) на дне этой ямки спец рецепторы.
Ямки обеспеч быструю инвагинацию и обр-е покрытых клатрином внутрикл вокуолей(скорость проникновения около 10 минут с момента адсорбции).
Покрытые вакуоли в кл слив с др вакуолями и обр-ют рецептосомы, которые могут сливаться с лизосомами, где происходит распад белков на аминокислоты.
Проникшие таким способом вирусы окружены мембраной.
2 – слияние вирусной и кл мембран происходит точечно, вз\д кл белка с липидами кл мембраны, в рез происходит интеграция мембран вируса и кл, а внутр комплекс выходит наружу.
Вирусы способствуют слиянию клеток и образованию синситиев.
Раздевание сопр-ся рядом хар-ных особенностей:
1.в рез распада вир частицы исчезает инф активность, появляется чув-ть к нуклеазам, возникает уст-ть к нейтрализир действию антител, теряется фоточувствительность при исп ряда препаратов.
Раздевание вирусов происходит в лизосомах, в яд порах, на яд мембранах, раздевание происходит равномерно.
58.Экспресия вирусного генома: особенности транскрипции вирусной НК.
Транскрипция – процесс перепис ДНК на РНК по законам генетич кода. Осущ с помощью РНК-полимеразы(спец фермент).
Продуктом транскрипции яв-ся мол ДНК.
Реализация генетич инф у вирусов:
1)у ДНК содерж: форма переноса генетич инф по схеме ДНК-транскрипт-РНК-трансл-белок
Транскрипция происходит в ядре:фермент ДНК-полимераза(герпес, аденовирусы).
У ДНК сод вирусов, у кот репрод происходит в цитоплазме есть фермент ДНК-полимераза, кот попад в клетку в составе вируса(оспа).
Нет необходимых спец белков
Реализация генетич мат: РНК-РНК-белок
Среди РНК сод вирусов имеются ретровирусы, которые осущ уник путь передачи генетич инф:
РНК перепис на ДНК, ДНК интегр с кл геномом и в его составе переписывается на РНК.
В составе этих вирусов есть вирус спец фермент – обратная транскриптаза или ривертаза.Этот же фермент синт нить ДНК на матрице. После синтеза ДНК замыкается в кольцо, интегрирует с кл геномом и далее исп РНК – полимераза кл для ф-я иРНК.
59. Экспрессия вирусного генома:трансляция вирусных белков.
Синтез белка происходит в рез трансляции ген информации.
Процесс трансляции состоит из 3-х фаз:
1)инициация(узнавание рибосомой иРНК)
2)элонгация(удлинение полипептидной цепи)
Стратегия вирусного генома, используещего кл аппарат трансляции. Например на подавление трансляции кл иРНК или на избират трансляцию вирусных иРНК, которых меньше чем клеточных.
Этот механизм реализуется на ур-е специф узнавания малой рибосомальной субъединицы, т.е на уровне инициирующего комплекса.
Специф узнавание рибосомой вирусных иРНК осущ-ся за счет вирус спец инициаторных факторов.
Способы формирования вирусных белков:
1)инф-РНК трансл в гигантский полипептид предшественника, который после синтеза посл-о нарезается на зрелые функц акт белки
2)иРНК трансл с образованием зрелых белков или белков,которые незначит модифиц после синтеза.
Модификация вир белков в осн хим:гликолизирование , ацелирование,мителирование,сульфирование,фосфолирирование, протелетич нарезание.
60.Экспрессия вирусного генома:репликация вирусного генома и сборка вирусных частиц.
Репликация – синтез мол нукл к-ты гомолог геномов.
Репл ДНК сод вирусов в осн катализ клеточными ферментами и мех сходен с мех репликации кл ДНК.Репликация генома РНК сод вирусов осущ-ся вирус спец ферментами-репликтаза.
Репликация однонитч РНК осущ-ся в 2 этапа:
1)синтез компл геному нитей
2)синтез компл генома
Репликативные комплексы:когда в зарож клетке происходит ф-е комплексов, в кот осущ процесс реплик геномов.
Сборка вирусных частиц.
Вирусы, репл которых происходит в ядрах-ядерные(в осн ДНК сод вирусы)вирусы, репл кот происх в цитоплазме(в осн РНК сод).
В основе самосборки лежит белок нуклеинов и белокбелковое узнавание.
Белокнукл узнавание огр неб уч молекулы нукл кислоты и опр-ся уник посл-ми нуклеотидов в некодир части вирусного генома, с этого участка нач процесс сборки вир частицы.
Далее присоединение особ белковых мол осущ-ся за счет спец белокбелков вз\д.
Сборка РНК сод вирусов.
Просто устр вирусы.
Сборка закл в ассоциации вир генома с виручными капсидными белками с образованием нуклеокапсида.
Сл-устроенные вирусы:нуклеокапсид мигрирует к месту сборки и выстраивается под участками мембран, с нар стороны кот уже встр вирусные суперкапсидные белки, постепенно нуклеокапсид выпячиваетсяы ч\з модифицир мембрану.
Все вирусные компоненты прибывают к месту сборки независимо др от друга. Первыми прибывают суперкапсидные белки(обычно гликопротеиды) вкл гликопр в опр уч-ке кл мембран приводит к модиф мембран
Сборка ДНК сод вирусов вкл в себя 4 этапа:
1)закл в ассоциации ДНК с внут белками и форм-е нуклеокапсида или сердцевин; - зак фор-ем неполных форм(промеж форма).
2)модиф белков незрелых вирионов нарез полипептиды(втор промежформа).
3)почкование пром форм ч\з яд мембрану в перенуклеарное пространство
4)достройка и созрев вирионов происходит в мембранах ЭПС и апп Гольджи, от куда вирус транспортируется по кл пов-ти.
Прикрепление вирусных частиц к поверхности клетки-хозяина — первая стадия инфекционного процесса. Начальный контакт вируса с клеткой происходит в результате случайного столкновения по типу броуновского движения.
В основе адсорбции лежат два механизма. Первый из них (неспецифический) определяется силами электростатического взаимодействия, возникающими между разноименно заряженными группами, расположенными на поверхности клетки и вирусами. Второй — специфический. Специфичность связи между вирусом и клеткой обусловлена комплементарными клеточными и вирусными рецепторами.
Процесс адсорбции возможен при наличии соответствующих рецепторов. Узнавание клеточных рецепторов вирусными белками (рецепторами), является высокоспецифическим процессом.
Максимальная скорость адсорбции вируса наблюдается лишь при определенном соотношении концентрации вируса и клеток, влиянии рН, температуры, ионного состава среды.
Процесс адсорбции состоит из двух быстро следующих друг за другом периодов: обратимого и необратимого. Период обратимого прикрепления может закончиться десорбцией. При длительном контакте вируса с клеткой никакие воздействия не позволяют освободить адсорбированный вирус, наступает стадия необратимой адсорбции.
Вирусные прикрепительные белки (вирусные рецепторы).
У просто организованных вирусов млекопитающих содержатся прикрепительные белки в составе капсида. У сложно организованных вирусов эти белки входят в состав суперкапсида и представлены разнообразными молекулами.
Проникновение вирусов в клетку
Исторически сложилось представление о двух альтернативных механизмах проникновения в клетку вирусов животных — путем эндоцитоза и путем слияния вирусной и клеточной мембран. Однако оба эти механизма не исключают, а дополняют друг друга.
Эндоцитоз означает, что вирусная частица попадает в цитоплазму в результате инвагинации участка плазматической мембраны и образования вакуоли, которая содержит вирусную частицу.
Рецепторный эндоцитоз. представляет собой частный случай рецепторного или адсорбционного эндоцитоза. Этот процесс является обычным механизмом, благодаря которому в клетку поступают питательные и регуляторные белки, гормоны, липопротеины и другие вещества из внеклеточной жидкости, Рецепторный эндоцитоз происходит в специализированных участках плазматической мембраны
Большинство оболочечных и безоболочечных вирусов животных проникает в клетку по механизму рецепторного эндоцитоза. Эндоцитоз обеспечивает внутриклеточный транспорт вирусной частицы в составе эндоцитарной вакуоли. Таким путем, например, ядерные вирусы попадают в ядро, а реовирусы в лизосомы.
Слияние вирусной и клеточной мембран. Для того чтобы внутренний компонент вируса мог пройти через клеточную мембрану, ряд оболочечных вирусов эволюционно приобрел механизм индукции слияния мембран. У оболочечных вирусов слияние обусловлено точечным взаимодействием вирусного белка слияния с липидами клеточной мембраны, в результате которого вирусная липопротеидная оболочка интегрирует с клеточной мембраной, а внутренний компонент вируса оказывается по другую ее сторону.
Вирусы вызывают два типа слияния клеток: 1) слияние снаружи и 2) слияние изнутри. Слияние снаружи происходит при высокой множественности инфекции. Напротив, слияние изнутри происходит при низкой множественности инфекции.
Раздевание вируса в клетке
Проникшие в клетку вирусные частицы должны раздеться для того, чтобы вызвать инфекционный процесс. Смысл раздевания заключается в удалении вирусных защитных оболочек, которые препятствуют экспрессии вирусного генома. В результате раздевания освобождается внутренний компонент вируса, который способен вызвать инфекционный процесс.
В ряде случаев способность вирусов вызвать инфекционный процесс определяется возможностью их раздевания в клетке данной системы. Тем самым эта стадия является одной из ограничивающих инфекцию.
Раздевание ряда вирусов происходит в специализированных участках внутри клетки (лизосомах, структурах аппарата Гольджи, околоядерном пространстве, ядерных порах на ядерной мембране).
Вторая фаза репродукции
Транскрипция
Это переписывание информации с ДНК на РНК по законам генетического кода. Транскрипция осуществляется специального фермента ДНК-зависимой РНК-полимеразы.
Продуктами транскрипции в клетке являются иРНК. Сама клеточная ДНК, являющаяся носителем генетической информации, не может непосредственно программировать синтез белка. Передачу генетической информации от ДНК к рибосомам осуществляет РНК-посредник (иРНК). На этом основана центральная догма молекулярной биологии, которая выражается следующей формулой:
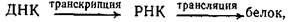
где стрелки показывают направление переноса генетической информации.
Транскрипция в зараженной клетке. Синтез комплементарных РНК на родительских матрицах с помощью родительской транскриптазы носит название первичной транскрипции в отличие от вторичной транскрипции, происходящей на более поздних стадиях инфекционного цикла на вновь синтезированных, дочерних матрицах с помощью вновь синтезированной транскриптазы. Большая часть иРНК в зараженной клетке является продуктом вторичной транскрипции.
Поперечные профили набережных и береговой полосы: На городских территориях берегоукрепление проектируют с учетом технических и экономических требований, но особое значение придают эстетическим.
Механическое удерживание земляных масс: Механическое удерживание земляных масс на склоне обеспечивают контрфорсными сооружениями различных конструкций.


Организация стока поверхностных вод: Наибольшее количество влаги на земном шаре испаряется с поверхности морей и океанов (88‰).
Для вирусов характерен дизъюнктивный (от disjuncus — разобщенный) способ репродукции-размножения. Потомство вируса возникает в результате сборки нуклеиновых кислот и белковых субъединиц, которые синтезируются раздельно клеткой хозяина.
Проникновение вируса в клетку и воспроизведение себе подобных проходит в несколько фаз:
1.проникновение в клетку хозяина,
2.синтез ферментов, необходимых для репликации вирусных нуклеиновых кислот,
3.синтез вирусных частей,
4.сборка и композиция зрелых вирионов,
5.выход зрелых вирионов из клетки.
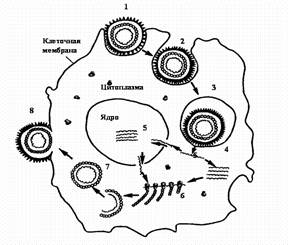
Стадии репродукции вирусов.
1 - адсорбция вириона на клетке; 2 - проникновение вириона в клетку путем виропексиса;
3 - вирус внутри вакуоли клетки; 4 - `раздевание вириона вируса; 5 - репликация вирусной нуклеиновой кислоты; 6 - синтез вирусных белков на рибосомах клетки; 7 - формирование вириона; 8 - выход вириона из клетки путем почкования.
Фаза I — адсорбция вириона на поверхности клетки.
Протекает в две стадии: первая — неспецифическая, когда вирус удерживается на поверхности клетки при помощи электростатических сил, т. е. благодаря возникновению противоположных зарядов между отдельными участками мембраны клеток и вируса. Эта фаза взаимодействия вируса с клеткой обратима, на нее оказывают влияние такие факторы, как рН и солевой состав среды.
Вторая стадия — специфическая, когда взаимодействуют специфические рецепторы вируса и рецепторы клетки, комплементарные друг другу. По химической природе рецепторы клетки могут быть мукопротеидами (или мукополисахаридами) и липопротеидами. Разные вирусы фиксируются на разных рецепторах: вирусы гриппа, парагриппа, аденовирусы — на мукопротеидах, а вирусы клещевого энцефалита, полиомиелита — на липопротеидах.
Фаза II — проникновение вируса в клетку. Электроноскопические наблюдения за процессом проникновения вирусов в чувствительные к ним клетки показали, что оно осуществляется посредством механизма, напоминающего пиноцитоз, или, как чаще называют, виропексис. В месте адсорбции вируса клеточная стенка втягивается внутрь клетки, образуется вакуоль, в которой оказывается вирион. Параллельно клеточные ферменты (липазы и протеазы) вызывают депротеинизацию вириона — растворение белковой оболочки и освобождение нуклеиновой кислоты.
Фаза III — скрытый период (период эклипса — исчезновения). В этот период в клетке невозможно определить наличие инфекционного вируса ни химическими, ни электронно-микроскопическими, ни серологическими методами. О сущности этого явления и его механизмов пока известно мало. Предполагается, что в скрытой фазе нуклеиновая кислота вируса проникает в хромосомы клетки и вступает с ними в сложные генетические взаимоотношения.
Фаза IV — синтез компонентов вириона. В этой фазе вирус и клетка представляют единое целое, вирусная нуклеиновая кислота выполняет генетическую функцию, индуцирует образование ранних белков и изменяет функцию рибосом. Ранние белки подразделяются на:
а) белки-ингибиторы (репрессоры), подавляющие метаболизм клеток
б) белки-ферменты (полимеразы), обеспечивающие синтез вирусных нуклеиновых кислот.
Синтез нуклеиновых кислот и белков протекает неодновременно и в разных структурных частях клетки. У вирусов, содержащих ДНК или РНК, эти процессы имеют некоторые различия и особенности.
Процесс формирования вирионов начинается спустя определенное время после того, как начал осуществляться синтез составляющих их компонентов. Продолжительность этого периода довольно вариабельна и предопределяется природой вируса — для РНК-содержащих обычно короче, чем для ДНК-вирусов. Например, продукция полных вирусных частиц осповакцины начинается приблизительно спустя 5—6 ч после инфицирования клеток и продолжается в течение последующих 7—8 ч, т. е. после того как синтез вирусной ДНК уже завершен.
Между нуклеиновой кислотой и соответствующим белковыми субъединицами образуются очень прочные связи, о чем свидетельствуют трудности отделения белка от вирусной нуклеиновой кислоты. Большую прочность вирусной частице придают входящие в ее состав углеводы и особенно липиды.
Формирование вирионов, так же как и синтез компонентов вируса, происходит в разных местах клетки, при участии различных клеточных структур. После завершения процесса формирования образуется зрелая дочерняя вирусная частица, обладающая всеми свойствами родительского вириона. Но иногда наблюдается образование так называемых неполных вирусов, которые состоят или только из нуклеиновой кислоты, или из белка, или из вирусных частиц, формирование которых остановилось в какой-то промежуточной стадии.
Фаза VI — выход зрелых вирионов из клетки. Существуют два основных механизма выхода зрелых вирионов из клетки:
1) выход вириона с помощью почкования. В этом случае наружная оболочка вириона происходит из клеточной мембраны, она содержит как материал клетки хозяина, так и вирусный материал;
2) выход зрелых вирионов из клетки через бреши в мембране. Эти вирусы не имеют наружной оболочки. При таком механизме выхода вирусов клетка, как правило, погибает и в среде появляется большое количество вирусных частиц.
Причиной гибели зараженной клетки могут быть три механизма:
2.защитная реакция клетки, запускающая генетическую программу ее гибели (апоптоз);
3. иммунная система организма, уничтожающая зараженную клетку.
Кроме продуктивного типа взаимодействия вируса и клетки возможно интегративное сосуществование или вирогения. Вирогения характеризуется интеграцией (встраиванием) нуклеиновой кислоты вируса в геном клетки, а также репликацией и функционированием вирусного генома как составной части генома клетки. Для интеграции с клеточным геномом необходимо возникновение кольцевой формы двунитевой ДНК вируса. Встроенная в состав хромосомы клетки вирусная ДНК называется провирусом. Провирус реплицируется в составе хромосомы и переходит в геном дочерних клеток, т.е. состояние вирогении наследуется. Под влиянием некоторых физических или химических факторов провирус может переходить в автономное состояние с развитием продуктивного типа взаимодействия с клеткой. Дополнительная генетическая информация провируса при вирогении сообщает клетке новые свойства, что может быть причиной развития опухолей, аутоиммунных и хронических заболеваний. На способности вирусов к интеграции с геномом клетки основаны персистенция (от лат. persisto - постоянно пребывать, оставаться) вирусов в организме и развитие персистентных вирусных инфекций. Например, вирус гепатита В способен вызывать персистирующие поражения с развитием хронического гепатита и часто опухолей печени.
Читайте также: