
Что такое наследственность бактерий и вирусов
Генетика микробов
Изменчивость и наследственность микроорганизмов является частью общебиологической проблемы изменчивости и наследственности. Наследственность и изменчивость в сущности две стороны одного явления. В природе постоянно наблюдается процесс передачи наследственных свойств организма из поколения в поколение и в то же время идет процесс изменчивости. Наследственность и изменчивость - это два противоречивых и вместе с тем неразрывно связанных между собой процесса. Они основа развития живого мира. По сравнению с другими организмами изменчивость микробов наблюдается чаще и осуществляется легче и быстрее, это объясняется большой быстротой их размножения и пластичностью.
Изменяться могут самые разнообразные свойства микробов - морфологические, ферментативные, антигенные, патогенные и др. Различают ненаследственную и наследственную изменчивость. Ненаследственная изменчивость (модификация) очень часто наблюдается под воздействием различных факторов внешней среды. Она заключается в количественном изменении некоторых свойств микроба, т. е. в ослаблении и утрате или усилении этих свойств. Когда воздействие факторов, вызвавших эти изменения, прекращается, то возникшие измененные признаки также утрачиваются. Наследственная изменчивость необратима, она развивается вследствие перестройки наследственного аппарата микроорганизма в результате непосредственного внешнего или внутреннего воздействия на него или внедрения чужеродного генетического материала (трансформация, конъюгация, рекомбинация и др.).
В микробиологии основоположником учения об изменчивости является Луи Пастер. Он искусственным путем получил необратимое ослабление вирулентности возбудителей сибирской язвы (1881) и бешенства (1885), а также получил способ приготовления живых вакцин для борьбы с этими заболеваниями. Это открытие Пастера имело поворотное значение для борьбы с инфекционными заболеваниями человека и животных. Применение живых вакцин привело к ликвидации многих эпидемий, сведя их до единичных заболеваний. Противосибиреязвенная вакцина получена Пастером путем длительного (12-24 дня) выращивания при повышенной температуре (42°) вирулентной культуры сибиреязвенной палочки.
Путем такой направленной изменчивости микробов в настоящее время получено 20 живых вакцин, из них 14 против вирусных и 6 против бактериальных инфекций. Все время идут работы по созданию новых вакцин и улучшению некоторых из существующих.
Благодаря простоте своего строения вирусы обладают большой способностью к изменчивости. Вирусы растений способны приспособляться к условиям жизни в организмах филогенетически очень отдаленных видов. Так, к вирусу табачной мозаики восприимчивы 236 видов растений 33 различных семейств. Из животных вирусов наибольшей изменчивостью обладает вирус гриппа. Вирусы гриппа, выделенные во время отдельных эпидемий в различных странах, отличаются от основных типов А и В по антигенному строению, морфологическим признакам, способности к адаптации и пр. В 1947-1952 гг. в Китае появилась новая разновидность - тип А2, которая вызвала эпидемию гриппа во всем мире. Об изменчивости вирусов говорит тот факт, что наибольшее количество живых вакцин получено из вирусов.
Изменчивость грибов имеет большое практическое значение, так как многие из них широко используются в различных производствах. Были получены расы дрожжей, хорошо развивающиеся при пониженных или повышенных температурах, требуемых в тех или иных производствах. Для интенсификации бродильных процессов получены расы дрожжей, способные использовать углеводы в больших концентрациях, а также приспосабливаться к высокому осмотическому давлению, к продуктам собственной жизнедеятельности, к различным физическим и химическим факторам. Применение этих рас дает большую продукцию. Ярким примером этого является производство антибиотиков.
За последние годы в изучении наследственности микроорганизмов наблюдаются большие достижения, имеющие значение не только для микробиологии, но и для общей генетики, биохимии и других наук. Проведенные генетические исследования приближают нас к реальной возможности влияния на процессы наследственности и изменчивости микробов.
Исходными для понимания явлений наследственности служат данные о химическом составе хромосом. Хромосомы высших организмов являются нуклеопротеидами, генетическим материалом в которых служит ДНК. У бактерий и фагов хромосомами является непосредственно ДНК, на нити которой в линейном порядке на определенных местах расположены определенные наследственные признаки. Количество хромосом в клетках различных организмов постоянно: у человека 46 хромосом, у кукурузы 10 и т. д. У бактерий весь генетический материал представляет одну хромосому. Это было найдено при помощи генетического метода.
Наследственность неразрывно связана с размножением, а размножение - с делением клеток, при котором происходит саморепродукция хромосом и ДНК. Соматические клетки имеют диплоидный, двойной набор хромосом. Дочерние клетки имеют такое же число хромосом, как и материнская (митоз). В половых клетках содержится вдвое меньше хромосом, так как при образовании половых клеток (мейоз) происходит особое редукционное деление с уменьшением нормального числа хромосом наполовину, например в половых клетках человека 23 хромосомы. Такой набор хромосом называется гаплоидным.
Из микроорганизмов только некоторые грибы и водоросли имеют типичный половой процесс с диплоидным и гаплоидным набором хромосом. Бактерии и фаги генетически ведут себя как организмы с одной гаплоидной хромосомой.
Характерно, что обычные морфологические методы смогли дать только общее описание ядерного вещества, применение же генетического метода исследования раскрыло тонкую структуру бактериальной хромосомы, хотя понятие хромосомы у бактерий носит несколько условный характер. Ген в настоящее время не является чисто умозрительной категорией, понятие гена получило вполне материальное содержание. Ген представляет собой участок (локус) хромосомы, слагающийся из цепи нуклеотидов молекулы ДНК, обладающий специфической функцией. Хромосома - это цепь линейно расположенных генов. Участок молекулы ДНК, составляющий ген, включает от 500 до нескольких тысяч нуклеотидов. Термину "ген" равнозначны: наследственный фактор, генетическая единица, детерминанта и др. Совокупность всех наследственных факторов (генов) в хромосомах организма называется генотипом. Фенотипом является внешний вид организма со всеми внешними и внутренними признаками. Фенотип бактерии есть результат взаимодействия ее генотипа и среды.
Появление в каком-либо организме новых наследственно передаваемых свойств может быть вызвано разными типами изменений в генетическом аппарате. Источником наследственной изменчивости могут быть мутации генов, т. е. изменения в их химическом строении, и структурные мутации хромосом. При этом могут происходить в двойной спирали ДНК следующие изменения: 1) замещение пары оснований, имевшихся в исходной молекуле ДНК, другой парой; 2) выпадение пары оснований из молекулы ДНК (деления); 3) внедрение новой пары оснований в молекулу ДНК; 4) инверсия - поворот нескольких пар оснований на 180°. Таким образом, в основе мутаций лежат молекулярные изменения в хромосоме.
Мутации у бактерий выявляются при наследственных изменениях любого признака микроба. Наиболее легко выявляются и количественно точно учитываются такие признаки: ауксотрофность к аминокислотам, пуринам и пиримидинам, витаминам; чувствительность или устойчивость к антибиотикам или фагам; лизогенность; ферментация углеводов и др. Предложены определенные обозначения мутантов. Так, ауксотрофность аминокислот обозначают начальными буквами или слогами их, например: гистидина - his, триптофана - try и т. д., двойные, тройные ауксотрофы обозначаются his try. Способность ферментировать углеводы, например лактозу - 1ас + , неспособность ферментировать - 1ас - . Чувствительность к стрептомицину - Str-s, резистентность - Str-r, зависимость - Str-d и т. д. Под влиянием мутагенных факторов были получены ауксотрофные мутанты, которые потеряли способность к синтезу важных для микроорганизмов веществ. Так, одни мутанты потеряли способность синтеза тех или иных аминокислот, другие мутанты - синтеза витаминов и т. д. Эти ауксотрофы (минус-варианты) могут расти только на средах, содержащих вещества, синтезировать которые они уже не могут. У них нарушена активность соответствующих ферментов. С помощью таких ауксотрофов удалось проследить нормальный биосинтез многих аминокислот, витаминов, углеводов, азотистых оснований.
Мутации у микробов так же, как и у высших организмов, делятся на спонтанные и индуцированные. Спонтанные мутации возникают при воздействии факторов, являющихся нормальными условиями среды. Индуцированные мутации возникают в результате обработки микробов мутагенными агентами. К последним относятся различные виды радиации (ультрафиолетовые, рентгеновские лучи, быстрые нейтроны, протоны и др.), действие температуры и пр. Спонтанные мутации наблюдаются очень редко. Частота их колеблется от 10 10 до 10 4 , т. е. одна мутантная клетка на 10 тыс. млрд. клеток. Мутагены увеличивают частоту мутаций в 10-100 тысяч раз.
Представление о строении генетического аппарата бактерийной клетки основано на изучении у них механизмов генетического обмена. У бактерий нет такого полового процесса, какой имеется у высших организмов. У бактерий обычно происходит только односторонняя передача части наследственного материала (фрагмента ДНК) от одной бактерийной клетки - донора к другой - реципиенту. При этом в клетке реципиента происходит взаимодействие между генетическим материалом донора и реципиента, которое приводит к образованию дочерних рекомбинантных клеток. Последние клетки сохраняют некоторые признаки реципиента и приобретают новые признаки, полученные от клетки донора. Механизмами такой рекомбинации, перегруппировки признаков в потомстве, т. е. изменчивости, являются трансформация, трансдукция и конъюгация.
Трансформация (превращение, перестройка) заключается в том, что некоторые бактерии при выращивании их в присутствии веществ, извлеченных из клеток родственных им видов или разновидностей, приобретают некоторые свойства последних. Пневмококки - возбудители воспаления легких - имеют несколько разновидностей. Когда к неболезнетворной бескапсульной разновидности прибавили убитые микробные тела болезнетворной разновидности, имеющей капсулу, и ввели эту смесь белым мышам, то белые мыши, вопреки ожиданию, погибли и из крови их была выделена болезнетворная капсульная разновидность (Ф. Гриффитс, 1928). Контрольные мыши, получившие только убитые микробные тела, не заболели. Таким образом, в организме мышей неболезнетворный пневмококк приобрел свойства болезнетворного, хотя и убитого, пневмококка. Это свойство стало наследственно закрепленным. После больших исследований было найдено, что эта наследственная изменчивость была вызвана ДНК убитых микробных тел болезнетворного пневмококка (О. Т. Эвери, 1944). В дальнейшем такие трансформации были получены при помощи одной только ДНК, полученной из микробных тел. Так же хорошо изучена трансформация устойчивости к антибиотикам - пенициллину, стрептомицину. Если ДНК, выделенную из антибиотиков устойчивых бактерий, добавить в культуру чувствительных к антибиотикам бактерий, то в культуре под влиянием ДНК некоторое количество клеток приобретает наследственную устойчивость к данному антибиотику. Активность ДНК, измеряемая в гаммах на 1 мл среды (v=10 -6 г), оказалась чрезвычайно высокой. Показана также возможность и межвидовой трансформации. Установление генетической роли ДНК имеет общебиологическое значение, так как позволило перейти к изучению различных биологических процессов на молекулярном уровне (синтез белка, нуклеиновых кислот, кодирование и др.).
Трансдукция - наследственно закрепленная передача признаков от одной бактерийной клетки (донора) другой клетке (реципиенту), не имеющей этих признаков, при помощи умеренного фага. Так наблюдалась передача свойств разлагать некоторые сахара, образовывать жгутики, споры, устойчивость к пенициллину. Это явление оказалось довольно распространенным среди бактерий. Некоторые созревающие частицы фага захватывают небольшие фрагменты ДНК бактерий, содержащие тот или иной признак, и передают их новой бактерийной клетке - хозяину. Предполагают, что генетический материал донорской клетки частично замещается генетическим материалом умеренного фага во время его пребывания в донорской клетке и этот материал затем фагом переносится в клетку-реципиент. Одновременно трансдуцируются один, реже два тесно сцепленных признака от одной бактериальной клетки к другой.
Конъюгация бактерий. Наличие конъюгации у бактерий наблюдалось некоторыми учеными давно. Но подробно она была изучена в последнее время. Существование конъюгации у бактерий впервые было доказано генетическим методом Ледербергом и Татум (1947). Конъюгация двух бактериальных клеток была обнаружена в электронном микроскопе. Клетки, из которых одна удлиненная - донор, а другая круглая и более крупная - реципиент, сближаются, между ними образуется мостик, по которому генетический материал донора переходит в клетку-реципиент. Был установлен односторонний перенос генетического материала донора, обозначаемого как F + (мужского типа), в клетку-реципиент, обозначаемую знаком F - (женского типа). Клетка-донор содержит особый фактор плодовитости, обозначаемый F-фактор. При скрещивании штаммов F - и F - рекомбинаций не образуется, они образуются при скрещивании штаммов F + и F - с частотой около 10 6 , т. е. очень незначительной.
В дальнейшем были найдены штаммы доноры F + , которые при скрещивании с F - образовывали рекомбинанты с очень большой частотой 10 1 -10 3 . Такие штаммы обозначаются High, frecuency of recombination Hfr. - высокая частота рекомбинации АШ, ЭФ, ЭР. Эти штаммы очень облегчили работу по изучению конъюгации.
При конъюгации происходит медленный перенос генетических факторов бактериальной хромосомы от донорской клетки в реципиентную в строгой последовательности их расположения в хромосоме донора. Количество перенесенных факторов зависит от продолжительности контакта клеток. Конъюгацию можно прекращать искусственно через различные промежутки времени путем встряхивания в особом смесителе, и таким образом можно определять, какие наследственные факторы и в какой последовательности расположены в хромосоме клетки. При помощи конъюгации оказалось возможным легко и более точно составить генетическую карту хромосомы бактерийной клетки.
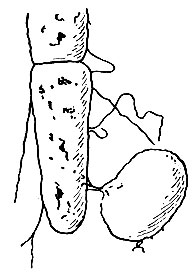
Рис. 48. Электронная микрограмма конъюгации кишечной палочки
Генетика вирусов изучалась главным образом на фагах. Хромосому вируса представляет одна молекула ДНК или РНК, находящаяся в белковой оболочке вируса. Основными признаками фага, наиболее часто используемыми в генетических исследованиях, являются: морфология бляшек на бактерийной культуре, способность фага реагировать с определенными штаммами бактерий хозяев, чувствительность фага к физическим и химическим факторам, лизогения и др. Генетический обмен, скрещивание фагов осуществляется путем заражения одной клетки бактерии двумя генетически различными фагами. В потомстве фага, возникшем в зараженной клетке, обнаруживаются особи с признаками каждого родительского фага и особи с объединенными признаками обоих родителей. Генетический материал фага представляет собой линейно расположенные локусы отдельных признаков. Так были построены генетические карты фагов группы Т и кишечной палочки К-12.
В настоящее время уже встал вопрос о дробимости гена, о его конечных размерах. Так, американец Бензер в опытах по детализации гена r11 фага Т4 разделил этот ген на два цистрона, которые состоят из более мелких единиц: реконов и мутонов. Реконом он считает минимальный участок хромосомы, способный к генетической рекомбинации. Мутон - минимальный участок хромосомы, изменение которого вызывает мутацию. Цистрон - функциональная единица. Он становится неактивным при мутационном повреждении входящего в его состав мутона или рекона. Бензер полагает, что если ген r11 состоит из 3000 нуклеотидов, то мутон его состоит всего из 7 нуклеотидов.
Практическая задача генетики - направленное получение микроорганизмов с полезными для человека свойствами. Значение ее особенно наглядно на примере создания микробов высокопродуктивных культур, образующих антибиотические, физиологически активные вещества, дефицитные аминокислоты и пр. Лучшим методом для этого является селекция мутантов, полученных при воздействии различных мутагенных факторов на исходные (дикие) штаммы. Процесс выведения высокопродуктивных штаммов состоит из многих ступенчатых этапов, состоящих из воздействия на культуру мутагенных факторов с последующим отбором наиболее продуктивного штамма. Этот мутантный штамм может еще подвергаться воздействию мутагенов с последующим отбором наиболее продуктивных мутантов. Так был получен советский штамм пенициллина "Новый гибрид 369", при выведении которого применялись гибридизация, ультрафиолетовые лучи, этиленимин и отбор. Этот штамм применяется на пенициллиновых заводах многих стран.
Во Всесоюзном институте антибиотиков в очень короткий срок была увеличена во много раз активность исходных штаммов при помощи воздействия на них ультрафиолетовых, рентгеновских лучей, химического соединения этиленимина и др. воздействий.
Сайт СТУДОПЕДИЯ проводит ОПРОС! Прими участие :) - нам важно ваше мнение.
ГЕНЕТИКА БАКТЕРИЙ
Противомикробный режим
Противомикробный режим — совокупность строго регламентированных техническими нормативными правовыми актами (Санитарными правилами, приказами Министерства здравоохранения) и обязательных для выполнения противомикробных мероприятий в лечебных, детских или иных учреждениях и производствах.
Ответственность за организацию и проведение санитарно-гигиенических мероприятий возлагается на руководителя учреждения (главного врача ЛПУ, заведующего ДДУ, директора школы, директора производства). Контролирующая функция по его соблюдению осуществляется врачом-эпидемиологом или врачом по коммунальной гигиене территориального центра гигиены и эпидемиологии. В штатной структуре крупных ЛПУ есть врач-эпидемиолог, осуществляющий вспомогательно-контролирующие функции.
В приемном отделении ЛПУ противомикробный режим предусматривает:
– исключение встречных потоков больных;
– осмотр кожных покровов и зева, термометрию поступающих больных для своевременного выявления подозрительных на инфекционные заболевания; в случае обнаружения больного с подозрением на инфекционное заболевание мебель, предметы и помещение дезинфицируют;
– полную санитарную обработку поступающего пациента (душ или ванна с продезинфицированной мочалкой, подстригание ногтей);
– маркировку инвентаря, кушетка должна быть покрыта клеенкой;
– уборку не реже 2 раз в день влажным способом с применением дезинфектантов.
В отделениях ЛПУ противомикробный режим предусматривает:
– наличие индивидуальных предметов ухода у каждого пациента;
– гигиеническую ванну (душ) не реже 1 раза в 7–10 дней;
– смену постельного и нательного белья не реже 1 раза в 7–10 дней, для послеоперационных больных, для пациенток послеродовых палат — ежедневно, для новорожденных — несколько раз в сутки по мере необходимости;
– проветривание палат не реже 4 раз в день;
– камерную дезинфекцию одежды перед передачей от одного больного другому; с разрешения главного врача пациент может пользоваться собственным бельем со сменой его по графику;
– больничную обувь протирают тампоном, смоченным 25 % формалином или 40 % уксусной кислотой, помещают в полиэтиленовый пакет на 3 часа, вынимают и проветривают 10–12 часов до исчезновения запаха препарата;
– при появлении случая ВБИ проводится внеочередной медосмотр всего персонала.
Генетика бактерий— наука о наследственности и изменчивости бактерий.
Наследственность бактерий — способность бактерий сохранять и передавать дочерним клеткам родительские признаки.
Бактерии обладают рядом важных преимуществ перед другими организмами и являются удобным материалом для генетики, так как:
– имеют короткий период генерации: многие бактерии делятся каждые 20–40 мин и быстро накапливают биомассу;
– многие бактерии легко культивируются в лабораторных условиях на сравнительно простых питательных средах;
– малые размеры микроорганизмов позволяют получать в одной пробирке или чашке и исследовать массовые популяции, состоящие из 10 8 –10 10 отдельных клеток, и благодаря этому выявлять такие редкие события, как мутация или передача приобретенного признака, не нуждаясь в сложных вспомогательных средствах и довольствуясь малым пространством;
– гаплоидны (имеют один набор генов), что исключает доминантность признаков;
– у бактерий высокая частота рекомбинаций и мутаций; так как большинство мутаций являются рецессивными, у диплоидных организмов их невозможно обнаружить в присутствии генов дикого (нормального) типа;
– генетический обмен выходит за пределы вида;
– некоторые имеют половую дифференциацию в виде донорских и реципиентных клеток;
– имеют различные интегрированные в хромосомы и обособленные фрагменты ДНК.
Нам важно ваше мнение! Был ли полезен опубликованный материал? Да | Нет
Молекулярная биология, изучающая фундаментальные основы жизни, является в значительной степени детищем микробиологии. В качестве основных объектов изучения в ней используют вирусы и бактерии, а основное направление- молекулярная генетика основана на генетике бактерий и фагов.
Бактерии- удобный материал для генетики. Их отличает:
- относительная простота генома (сопокупности нуклеотидов хромосом);
- гаплоидность (один набор генов), исключающая доминантность признаков;
- различные интегрированные в хромосомы и обособленные фрагменты ДНК;
- половая дифференциация в виде донорских и реципиентных клеток;
- легкость культивирования, быстрота накопления биомасс.
Общие представления о генетике.
Ген- уникальная структурная единица наследственности, носитель и хранитель жизни. Он имеет три фундаментальные функции.
1.Непрерывность наследственности- обеспечивается механизмом репликации ДНК.
2.Управление структурами и функциями организма - обеспечивается с помощью единого генетического кода из четырех оснований (А- аденин, Т- тимин, Г- гуанин, Ц- цитозин). Код триплетный, поскольку кодон- функциональная единица, кодирующая аминокислоту, состоит из трех оснований (букв).
3.Эволюция организмов- благодаря мутациям и генетическим рекомбинациям.
В узкоспециальном плане ген чаще всего представляет структурную единицу ДНК, расположение кодонов в которой детерминирует первичную структуру соответствующей полипептидной цепи (белка). Хромосома состоит из особых функциональных единиц- оперонов.
Основные этапы развития (усложнения) генетической системы можно представить в виде следующей схемы:
кодон à ген à оперон à геном вирусов и плазмид à хромосома прокариот (нуклеоид) à хромосомы эукариот (ядро).
Генетический материал бактерий.
1.Ядерные структуры бактерий- хроматиновые тельца или нуклеоиды (хромосомная ДНК). У бактерий одна замкнутая кольцевидная хромосома (до 4 тысяч отдельных генов). Бактериальная клетка гаплоидна, а удвоение хромосомы (репликация ДНК) сопровождается делением клетки. Вегетативная репликация хромосомной (и плазмидной) ДНК обусловливает передачу генетической информации по вертикали- от родительской клетки- к дочерней. Передача генетической информации по горизонтали осуществляется различными механизмами- в результате конъюгации, трансдукции, трансформации, сексдукции.
2.Внехромосомные молекулы ДНК представлены плазмидами, мигрирующими генетическими элементами- транспозонами и инсервационными (вставочными) или IS- последовательностями.
Плазмиды- экстрахромосомный генетический материал (ДНК), более просто устроенные по сравнению с вирусами организмы, наделяющие бактерии дополнительными полезными свойствами. По молекулярной массе плазмиды значительно меньше хромосомной ДНК, содержат от 40 до 50 генов.
Их объединение в одно царство жизни с вирусами связано с наличием ряда общих свойств- отсутствием собственных систем мобилизации энергии и синтеза белка, саморепликацией генома, абсолютным внутриклеточным паразитизмом.
Их выделение в отдельный класс определяется существенными отличиями от вирусов.
1.Среда их обитания- только бактерии (среди вирусов , кроме вирусов бактерий- бактериофагов имеются вирусы растений и животных).
2.Плазмиды сосуществуют с бактериями, наделяя их дополнительными свойствами. У вирусов эти свойства могут быть только у умеренных фагов при лизогении бактерий, чаще же всего вирусы вызывают отрицательный последствия, лизис клеток.
3.Геном представлен двунитевой ДНК.
4.Плазмиды представляют собой “голые” геномы, не имеющие никакой оболочки, их репликация не требует синтеза структурных белков и процессов самосборки.
Плазмиды могут распространяться по вертикали (при клеточном делении) и по горизонтали, прежде всего путем конъюгационного переноса. В зависимости от наличия или отсутствия механизма самопереноса (его контролируют гены tra- оперона) выделяют конъюгативные и неконъюгативные плазмиды. Плазмиды могут встраиваться в хромосому бактерий- интегративные плазмиды или находиться в виде отдельной структуры- автономные плазмиды ( эписомы).
Классификация и биологическая роль плазмид.
Функциональная классификация плазмид основана на свойствах, которыми они наделяют бактерии. Среди них- способность продуцировать экзотоксины и ферменты, устойчивость к лекарственным препаратам, синтез бактериоцинов.
Основные категории плазмид.
1.F- плазмиды - донорские функции, индуцируют деление (от fertility - плодовитость). Интегрированные F - плазмиды- Hfr- плазмиды (высокой частоты рекомбинаций).
2.R- плазмиды (resistance) - устойчивость к лекарственным препаратам.
3.Col- плазмиды- синтез колицинов (бактериоцинов)- факторов конкуренции близкородственных бактерий (антогонизм). На этом свойстве основано колицинотипирование штаммов.
4.Hly- плазмиды- синтез гемолизинов.
5.Ent- плазмиды- синтез энтеротоксинов.
6.Tox- плазмиды- токсинообразование.
Близкородственные плазмиды не способны стабильно сосуществовать, что позволило объединить их по степени родства в Inc- группы (incompatibility- несовместимость).
Биологическая роль плазмид многообразна, в том числе:
- контроль генетического обмена бактерий;
- контроль синтеза факторов патогенности;
- совершенствование защиты бактерий.
Бактерии для плазмид- среда обитания, плазмиды для них- переносимые между ними дополнительные геномы с наборами генов, благоприятствующих сохранению бактерий в природе.
Мигрирующие генетические элементы - отдельные участки ДНК, способные определять свой перенос между хромосомами или хромосомой и плазмидой с помощью фермента рекомбинации транспозазы. Простейшим их типом являются инсерционные последовательности (IS- элементы) или вставочные элементы, несущие только один ген транспозазы, с помощью которой IS- элементы могут встраиваться в различные участки хромосомы. Их функции- координация взаимодействия плазмид, умеренных фагов, транспозонов и генофора для обеспечения репродукции, регуляция активности генов, индукция мутаций. Величина IS- элементов не превышает 1500 пар оснований.
Транспозоны (Tn- элементы) включают до 25 тысяч пар нуклеотидов, содержат фрагмент ДНК, несущий специфические гены, и два Is- элемента. Каждый транспозон содержит гены, привносящие важные для бактерии характеристики, как и плазмиды (множественная устойчивость к антибиотикам, токсинообразование и т.д.). Транспозоны- самоинтегрирующиеся фрагменты ДНК, могут встраиваться и перемещаться среди хромосом, плазмид, умеренных фагов, т.е. обладают потенциальной способностью распространяться среди различных видов бактерий.
Понятие о генотипе и фенотипе.
Генотип- вся совокупность имеющихся у организма генов.
Фенотип- совокупность реализованных (т.е. внешних) генетически детерминированных признаков, т.е. индивидуальное (в определенных условиях внешней среды) проявление генотипа. При изменении условий существования фенотип бактерий изменяется при сохранении генотипа.
Изменчивость у бактерий может быть ненаследуемой (модификационной) и генотипической (мутации, рекомбинации).
Временные, наследственно не закрепленные изменения, возникающие как адаптивные реакции бактерий на изменения окружающей среды, называются модификациями (чаще - морфологические и биохимические модификации). После устранения причины бактерии реверсируют к исходному фенотипу.
Стандартное проявление модификации- распределение однородной популяции на две или более двух типов- диссоциация. Пример- характер роста на питательных средах: S- (гладкие) колонии, R- (шероховатые) колонии, M- (мукоидные, слизистые) колонии, D- (карликовые) колонии. Диссоциация протекает обычно в направлении Sà R. Диссоциация сопровождается изменениями биохимических, морфологических, антигенных и вирулентных свойств возбудителей.
Мутации- скачкообразные изменения наследственного признака. Могут быть спонтанные и индуцированные, генные (изменения одного гена) и хромосомные (изменения двух или более двух участков хромосомы).
Одновременно у бактерий имеются различные механизмы репарации мутаций, в том числе с использованием ферментов- эндонуклеаз, лигаз, ДНК- полимеразы.
Генетические рекомбинации- изменчивость, связанная с обменом генетической информации. Генетические рекомбинации могут осуществляться путем трансформации, трансдукции, конъюгации, слияния протопластов.
1.Трансформация- захват и поглощение фрагментов чужой ДНК и образование на этой основе рекомбинанта.
2.Трансдукция- перенос генетического материала фагами (умеренными фагами- специфическая трансдукция).
3.Конъюгация- при непосредственном контакте клеток. Контролируется tra (transfer) опероном. Главную роль играют конъюгативные F- плазмиды.
Геном вирусов содержит или РНК, или ДНК (РНК- и ДНК- вирусы соответственно). Выделяют позитивную (+) РНК, обладающую матричной активностью и соответственно- инфекционными свойствами, и негативную ( - ) РНК, не проявляющую инфекционные свойства, которая для воспроизводства толжна транскрибироваться (превращаться) в +РНК. Механизмы репродукции различных вирусов очень сложные и существенно отличаются. Основные их схематические варианты представлены ниже.
1. вирионная (матричная) +РНК à комплементарная -РНК (в рибосомах) à вирионная +РНК.
2. - РНК à вирусная (информационная) +РНК à - РНК (формируется на геноме зараженной клетки).
3. однонитевая ДНК: +ДНК à +ДНК -ДНК à +ДНК -ДНК +ДНК à +ДНК.
4. ретровирусная однонитевая РНК: РНК à ДНК (провирус) à РНК.
5. двунитевая ДНК: разделение нитей ДНК и формирование на каждой комплементарной нити ДНК.
Генофонд вирусов создается и пополняется из четырех основных источников:
двух внутренних (мутации, рекомбинации) и двух внешних (включение в геном генетического материала клетки хозяина, поток генов из других вирусных популяций).
Комплементация- функциональное взаимодействие двух дефектных вирусов, способствующее их репликации и горизонтальной передаче.
Фенотипическое смешивание- при заражении клетки близкородственными вирусами с образованием вирионов с гибридными капсидами, кодируемыми геномами двух вирусов.
Популяционная изменчивость вирусов связана с двумя разнонаправленными процессами - мутациями и селекцией, связанными с внешней средой как индуктором мутаций и фактором стабилизирующего отбора. Гетерогенность вирусных популяций- адаптационный генетический механизм, способствующий пластичности (устойчивости, приспособляемости) популяций, фактор эволюции и сохранения видов во внешней среде.
Генофонд вирусных популяций сохраняется за счет нескольких механизмов:
- восстановления изменчивости за счет мутаций;
- резервирующих механизмов (возможность перехода любых, даже негативных мутаций в следующую генерацию)- комплементация, рекомбинация;
- буферных механизмов (образование дефектных вирусных частиц, иммунных комплексов и др.), способствующие сохранению вируса в изменяющихся внешних условиях.
Читайте также: