
Что такое онкогенные ретровирусы
Ретровирусы - оболочечные, сферические вирусы, которые выходят почкованием через клеточную мембрану хозяина. Они имеют приблизительно 100 нм в диаметре. Геном состоит из двух идентичных линейных односпиральных молекул РНК. Икосаэдральный нуклеокапсид содержит спиральный рибонуклеопротеид и окружен оболочкой состоящей из гликопротеидов и липидов.
Характерная особенность ретровирусов - присутствие в вирионе необычного фермента - РНК зависимой ДНК полимеразы или обратной транскриптазы(отсюда имя retro, означающее обратно). В отличие от классической транскрипции генетической информации от ДНК на РНК, фермент обратная транскриптаза готовит ДНК-копию РНК-генома ретровируса - первоначально РНК-ДНК гибрид, а затем его двухспиральную ДНК-форму. Двухспиральная ДНК-форма ретровирусного генома, называемая провирусом, интегрирует в ДНК инфицированной клетки-хозяина. Именно от провируса транслируются все ретровирусные белки. Заражение онкогенным ретровирусом не ведет к цитолизу или гибели инфицированных клеток, но провирус остается интегрированным в ДНК клетки-хозяина до конца жизни клетки и воспроизводится вместе с клеточным геномом при размножении клеток.
В то время как все онкогенные РНК-содержащие вирусы принадлежат семейству Retroviridae, не все ретровирусы онкогенны. Семейство Retroviridae классифицируется на три подсемейства.
1. Oncovirinaeвключает все онкогенные РНК содержащие вирусы (прежде называемое онкорнавирус).
Ретровирусы широко распространены; их находят почти у всех позвоночных, включая животных, птиц и рептилий. Основываясь на круге хозяев и типах вызываемых болезней, онкогенные Ретровирусы можно разделить на следующие группы:
1. Вирусы лейкозно-саркоматозного комплекса птиц. Группа антигенно родственных вирусов, которые вызывают Avian лейкозы (вирусы лимфоматоза, миелобластоза и эритробластоза) или саркому у домашних птиц (вирус саркомы Рауса, ВСР).
2. Вирусы мышиных лейкозов. Эта группа состоит из нескольких штаммов вирусов мышиной лейкемии и вирусов саркомы, названных по имени исследователи впервые описавших их (например Гросс, Френд, Молони, Раушер).
4. Вирусы лейкозов и сарком других животных.Большое количество вирусов было выделено из лейкозов и сарком различных видов животных - кошек, хомяков, крыс, морских свинок и обезьян.
Видовая специфичность. Ретровирусы обычно поражают только один вид хозяина, специфика обусловлена главным образом присутствием вирусных рецепторов на поверхности клетки-хозяина. В зависимости от их способности расти в клетках другого вида, ретровирусы делятся на 1) экотропные (размножаются только в клетках естественного хозяина); 2) амфитропные (размножаются в клетках естественного и чужих видов); и 3) ксенотропные (размножаются только в клетках чужих видов, но не в клетках естественных хозяев).
Резистентность. Ретровирусы неустойчивы, инактивируются при 56 о С в течение 30 минут, слабыми кислотами, эфиром и формалином. Они устойчивы при – 30 о С
Морфология.Ретровирусы существуют в виде четырех морфологических типов. Частицы типа А существуют только внутри клеток. Они имеют 60-90 нм в диаметре и содержат кольцевидный нуклеоид, окруженный мембраной. Они могут являться формой предшественника других типов. Типы B, C и D являются внеклеточными. Диаметр В частицы – 100-130 nm, с эксцентрическим нуклеоидом и несут поверхностные шипики. Частицы С типа имеют центральный нуклеоид и гладкую поверхностную мембрану. Частицы D типа еще не охарактеризованы. Они имеют эксцентрический нуклеоид и несут короткие поверхностные шипики.
Большинство ретровирусов – частицы С типа. Вирус рака молочных железы мыши - частица типа B, а вирус рака молочной железы обезьян Мэзон-Пфайзера - частица типа D.
Антигены.Имеется два типа антигенов – типоспецифические гликопротеидные антигены, расположенные на оболочке, и группо-специфические нуклеопротеидные антигены, расположенные в ядре вириона. Перекрестные реакции между поверхностными антигенами ретровирусов от различных видов хозяев не наблюдаются.
Геномная структура.Ретровирусы имеют относительно простую геномную структуру.
Провирус стандартного ретровируса (такого как недефектный вирус лейкоза птиц или мышей) состоит из трех генов, требуемых для вирусной репликации - gag, pol, и env. Ген gag кодирует белки нуклеокапсида, которые являются группоспецифическими антигенами, ген polкодирует РНК-зависимую ДНК-полимеразу, ген envкодирует гликопротеиды оболочки. С обоих концов провируса имеется длинный концевой повтор (LTR), непосредственно связывающийся с ДНК клетки-хозяина. LTR-участки обеспечивают контроль регуляции функции генов провируса.
Некоторые ретровирусы (трансрегулирующие вирусы) типа HTLV или HIV несут четвертый ген tat после envгена. Это – трансактивирующий ген, который регулирует функцию вирусных генов.

Инфекция, вызванная онкогенными вирусами, является причиной 15,4% заболеваний раком по всей земле. Первый онкогенный вирус, который был идентифицирован — это куриный вирус саркомы Рауса в 1911 году. Десятилетиями позже были обнаружены серии других онкогенных вирусов, в том числе вирус папилломы Шопа (вирус папилломы кроличьего хвоста), вирус опухоли молочной железы мыши, аденовирус, вирус SV40 (обезьяний вирус sv40). Теория опосредованного вирусом онкогенеза в итоге была экспериментально продемонстрирована в 1976 году Гарольдом Вармусом и Майклом Бишопом путем превращения инфицированных клеток в опухолевые вирусом саркомы (ген v-src вызывает образование опухоли после захвата протоонкогена человеческой клетки в результате трансдукции).
Несмотря на то, что онкогенные вирусы нашли в исследованиях онкогенной активности животных клеток, они были не в состоянии преобразовать человеческие клетки. Теория вирусного онкогенеза в человеческой клетке оставалась спорной до 1965 года, когда вирус Эпштейн-Барр (EBV) был обнаружен в клетках лимфомы Беркитта. В ходе последующих исследований, в том числе выделении Т-лимфотропного вируса человека (HTLV) из клеток Т-клеточной лимфомы и выявлении роли папилломовирусов человека с высокой онкогенной активностью в развитии рака шейки матки, был определен путь для понятия человеческих онкогенных вирусов. Обнаружение связи между герпес-вирусом и саркомой Капоши (KSHV) и лимфомой и между полиомавирусом клеток Меркеля (MCV) и карциномой клеток Меркеля (MCC) подчеркнуло возможность открытия еще большего количества онкогенных вирусов с помощью современных технологий. Далее каждый вирус будет рассмотрен отдельно.
Вирус Эпштейн-Барр
Вирус Эпштейн-Барр (ВЭБ, EBV) — ДНК-содержащий вирус, который преимущественно поражает B-лимфоциты, также является митогеном для них; вызывает лимфопролиферативные болезни (лимфома Беркитта, носоглоточная карцинома, NK-клеточная лейкемия, лимфома Ходжкина и т.д.), а также инфекционный мононуклеоз. Вирионы вируса были обнаружены при электронной микроскопии биоптата лимфомы Беркитта в начале 1964 года Энтони Эпштейном и Ивонной Барр.
Строение. ВЭБ имеет вирусный капсидный антиген (VCA), ядерные антигены-EBNAs 1, 2, 3, 3a, 3b, 3c; латентные мембранные протеины (LMPs) 1, 2 и две маленькие Эпштейна-Барр-кодируемые РНК (EBER) молекулы — EBER1 и EBER2. EBNAs и LMPs являются ДНК-связывающими белками, считающимися необходимыми для развития инфекции и фиксации.
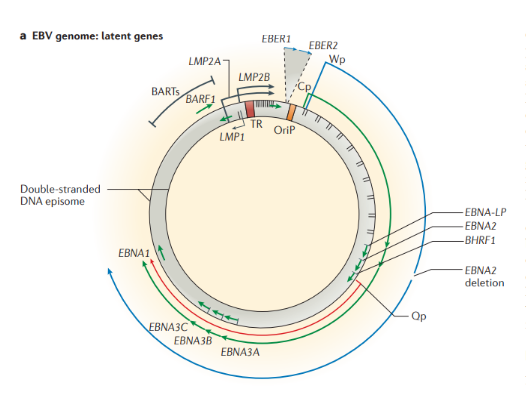
- необходим для трансформации В-лимфоцитов;
- функционирует как конститутивный активатор рецептора ФНО и имеет сходство с CD-40 рецептором;
- активирует сигнальные пути, которые делают вклад в онкогенный эффект.
В зависимости от набора экспрессируемых вирусных генов в пораженных клетках выделяют три типа латентности (I, II и III тип), характерные для различных видов лимфом: I тип подразумевает избирательную экспрессию EBNA-1 при ВЭБ-положительной лимфоме Беркитта; II тип заключается в экспрессии EBNA-1, LMP-1 и LMP-2 и является отличительным признаком ВЭБ-положительной ходжкинской лимфомы, периферических T/NK-клеточных лимфом; латентность III типа, характеризуемая экспрессией всего набора из девяти латентных белков ВЭБ, наблюдается при лимфопролиферативных поражениях, возникающих у пациентов с тяжелым иммунодефицитом (после трансплантации паренхиматозных органов или стволовых клеток, ВИЧ-инфекции). Тип латентности определяет восприимчивость инфицированных клеток к различным иммунотерапевтическим тактикам. Индуцированные ВЭБ злокачественные новообразования связаны с экспрессией латентных генов. У большинства иммунокомпетентных носителей ВЭБ вирус не индуцирует опухолевый процесс. Новообразования развиваются при комплексном взаимодействии ВЭБ, иммуногенетических факторов, факторов внешней среды (паразитарных инфекций — чаще, малярии, недоедании, потреблении пищи, содержащей канцерогены) и иммунодефицита (ВИЧ-инфекция, трансплантация).
Папилломавирусы человека
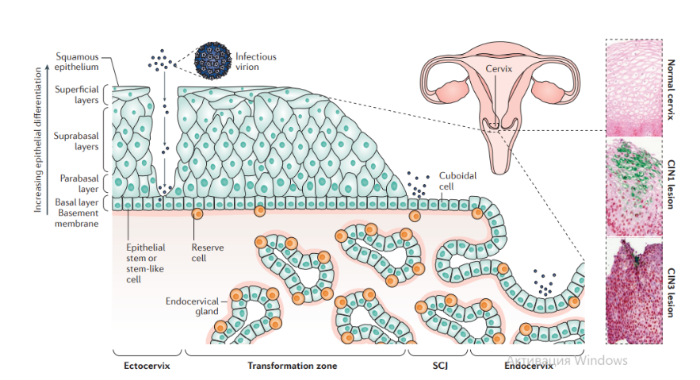
Описано около 200 папилломавирусов. Вирусы передаются при контактах через микротравмы кожи и слизистых оболочек, а также половым путем. Папилломавирусы человека вызывают доброкачественные кожные, генитальные, оральные и конъюнктивальные папилломы; индуцируют пролиферацию эпителия, обладают онкогенным потенциалом. С папилломавирусом, особенно ПВЧ-5 и ПВЧ-8, тесно связано развитие немеланомных разновидностей рака кожи. Различают низкоонкогенные папилломавирусы, вызывающие бородавки, которые не малигнизируются, и высокоонкогенные, способные вызвать рак шейки матки, — ПВЧ-16, 18, 31, 33, 35, 45, 51, 52, 58.
Строение. Капсид икосаэдрический, состоит из двух капсидных белков (L1-L2), формирующих 72 пентамера. Геном — двунитевая циркулярная ДНК; имеет в зависимости от вируса восемь ранних генов (E1-E8) и два поздних гена (L1-L2). Гены E6-E7 обладают онкогенной активностью.
Патогенез. В целом, для развития продуктивного поражения ВПЧ (высокого или низкого риска) требуется, чтобы вирус имел доступ к эпителию базального слоя как только эпителиальный барьер был подвержен риску. Папилломавирусы должны инфицировать делящуюся клетку, чтобы стать признанными как малое количество копий ядерной эписомы, и для развития стойкого поражения первичная инфицированная клетка, вероятно, должна быть похожа на долгоживующую эпителиальную стволовую клетку или похожую на нее. Кроме того, связанные с заживлением ран изменения в местном микроокружении, включая увеличение количества факторов роста, могут играть ключевую роль в создании резервуара инфекции в базальном слое. Способность ВПЧ инфицировать, и реплицироваться, и управлять прогрессированием рака зависит от функции вирусного белка, положении в эпителии, где эти белки экспрессируются. Как только инфицированные базальные клетки поделились и в конечном итоге вошли в парабазальный слой, они начинают проходить программу экспрессии генов, которая связана с дифференциацией. Амплификация вирусного генома обычно происходит в срединно-эпителиальных слоях, тогда как сбор и выход вируса происходит вблизи наружной поверхности эпителия.
Несмотря на то, что эффективная амплификация генома требует комбинированного действия множественных продуктов вирусного гена, включая E6, E7, E2 и кодированную вирусом геликазы E1, поддержание репликации в базальном слое эпителия может зависеть от факторов репликации клетки хозяина. Программа, зависящая от дифференциации, которая происходит в надбазальных слоях эпителиальной клетки, включает как временные изменения в активности промотора, так и изменения в шаблонах сплайсинга мРНК, которые облегчают производство различных продуктов вирусных генов на разных стадиях во время программы дифференциации. Функция вирусного белка далее регулируется посттрансляционной модификацией, включая фосфорилирование и протеолитическое деление клетки.
Герпесвирус, ассоциированный с саркомой Капоши
KSHV, или человеческий герпесвирус 8, был обнаружен в 1994 году как член семейства гамма-герпеса человека, присоединившись к ВЭБ. Инфекция иммунокомпрометированных индивидуумов с KSHV была связана с развитием саркомы Капоши, полученной из эндотелиальных клеток, и по меньшей мере двумя B-лимфоцит лимфопролиферативными заболеваниями: первичная выпотная лимфома и многоцентричная болезнь Кастлмана. Однако изучение патогенеза и онкогенеза KSHV затруднено отсутствием значимой модели животных и восприимчивой клеточной культуры.
Онкогенное действие. Две иммортализованные клеточные линии, KS Y-1 и SLK, когда-то использовались для исследований KS и KSHV, но в KS Y-1 были также обнаружены клетки линии рака мочевого пузыря T 24, что затруднило дальнейшую работу. Первичные эмбриональные метанефрические мезенхимальные клетки-предшественники почек крысы восприимчивы к инфекции и трансформации KSHV, но из них распространяются ограниченное количество инфекционных вирионов. Линии B-клеток, полученные из первичной выпотной лимфомы, обычно заражаются KSHV на латентной стадии и могут быть индуцированы для получения малого количества вирионов KSHV, но первичные B-лимфоциты из периферической крови или тонзиллярной ткани являются устойчивыми к инфекции KSHV, и для их инфицирования может потребоваться совместный рост с KSHV-позитивными клетками.
KSHV кодирует несколько важных белков, которые обладают некоторой онкогенной активностью для индуцирования пролиферации клеток, иммортализации, трансформации и передачи сигналов; производства цитокинов; иммунного контроля; антиапоптозной активности; и ангиогенеза. К ним относятся вирусные латентные белки, латентный ассоциированный ядерный антиген (LANA), vFLIP (FADD (Fas-ассоциированный белок с доменом смерти), подобный ферменту, ингибирующему синтез интерферона или ингибирующему белок каспазы 8 (FLICE)), vCyclin и вирусные литические белки G-белкового рецептора (vGPCR), интерферон-регуляторный фактор 1 (vIRF-1) и K1. Хотя истинную онкогенную природу каждого белка еще предстоит определить, накопление доказательств указывает на то, что каждый из них вносит свой вклад в онкогенез KSHV. Таким образом, полный спектр вызванной KSHV злокачественности может потребовать, чтобы несколько онкогенных продуктов работали вместе в присутствии клеток хозяев и экологических факторов. Например, как LANA, так и vIRF-1 нацелены на клеточный опухолевый супрессор p53. LANA также ингибирует pRB и PP2A.vCyclin, активатор CDK4 6,198, подавляет p27kip1, ингибитор CDK199 и вызывает реакцию на задержку старения G1, результаты от гиперактивации NF-κB. vFLIP и K1 активируют путь сигнала NF-κB для предотвращения апоптоза B-клеток. vGPCR и K1 влияют на сигнальные пути AKT и NF-κB и способствуют ангиопролиферативному и воспалительному поражению саркомой Капоши.
Т-лимфотропный вирус человека (HTLV)
T-лимфотропный вирус человека (HTLV) относится к семейству ретровирусов, является возбудителем T-клеточного лимфолейкоза взрослых. Вирус впервые был изолирован в 1980 году от больного Т-лимфомой.
Геном. Gag, Pol и Env являются вирусными структурными белками, другие — вирусными регуляторными/вспомогательными белками. За исключением гена hbz, который кодируется минус-цепью провирусного генома HTLV из 3'-LTR, все остальные гены кодируются плюсовой нитью по направлению 5'-LTR. Следует отметить, что 5'-LTR часто удаляется или метилируется по мере прогрессирования заболевания. Кроме того, на поздних стадиях лейкемиогенеза в гене tax часто возникают нонсенс- или миссенс-мутации. Хотя белок tax и ген HBZ индуцируют опухоли у трансгенных мышей, а p12 проявляет слабую онкогенную активность, ни один из вирусных белков/генов, кроме tax, не требуется для опосредованного HTLV-1 опухолеобразования. Продукт tax гена действует на терминальные повторы LTR, стимулируя синтез вирусной иРНК, а также образование рецепторов IL-2 на поверхности зараженной клетки.
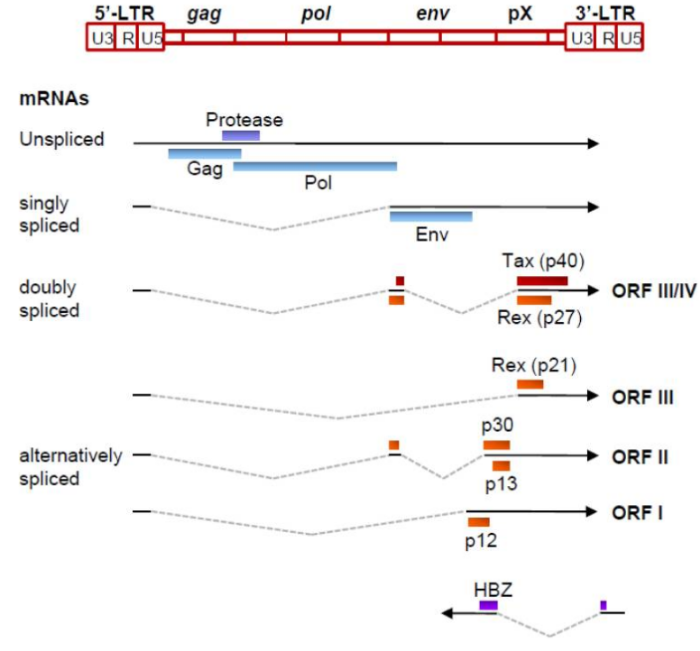
Рисунок 3. Gag, Pol и Env вирусные регуляторные/вспомогательные белки HTLV
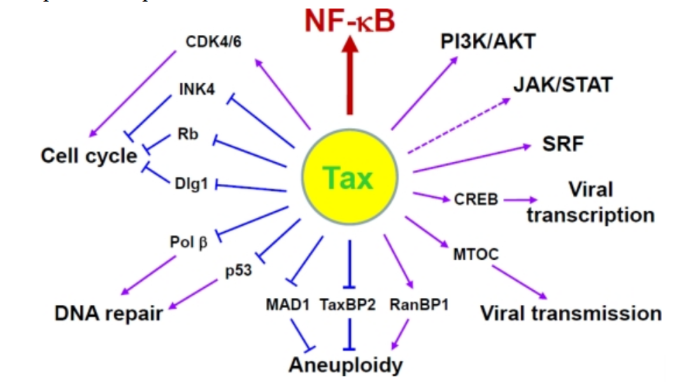
Рисунок 4. К леточные пути вирусного онкобелка tax
Патогенез. Проникновение HTLV-1 в клетку человека охарактеризовывает начало выживания и репликации вируса. HTLV-1 связывается с рецептором клеток, имеющих необходимый антиген, что инициирует процесс инвазии. Рецепторный комплекс состоит из транспортера глюкозы (GLUT1), гепаринсульфат протеогликана (HSPG) и рецептора VEGF-165 нейропилина-1 (NRP-1). HTLV-1 взаимодействует с HSPG. Процесс слияния происходит через комбинацию GLUT1 с HSPG/NRP-1, после чего ядро, содержащее вирусную РНК, доставляется в цитоплазму клеток-мишеней. После обратной транскрипции HTLV-1 встраивает свой геном в генома хозяина, чтобы сформировать провирус, окруженный двумя LTR на 5' и 3' концах. Затем провирус транскрибируется и кодирует структурные, регуляторные и вспомогательные белки. Впоследствии вирусные геномные РНК и Gag, Env и Gag-Pol белки переносятся в плазматическую мембрану, чтобы начать собираться в зрелую вирусную частицу.
Вирусы гепатита
Вирус гепатита В (HBV) и вирус гепатита С (HCV) представляют собой несвязанные вирусы, имеющие тропность к гепатоцитам и реплицирующиеся в них. Около 2 миллиардов человек инфицированы HBV, и более 350 миллионов человек становятся хроническими носителями. Только 5-10% взрослых, которые приобретают инфекцию, становятся хроническими носителями, у 30% развиваются прогрессирующие хронические заболевания печени: гепатит, фиброз, цирроз и, наконец, гепатоцеллюлярная карцинома (HCC).
Патогенез. HCC, индуцированная посредством HBV и HCV, развивается в области воспаления и регенерации, что является результатом хронического повреждения печени, а это в свою очередь указывает на то, что патогенез HCC иммунно-опосредован. Эти вирусы постоянно реплицируются в культуре клеток без явных повреждений и смерти клеток, подразумевая, что они являются не цитопатическими. Постоянная репликация вируса является фактором риска для образования HCC, потому что воспаление часто приводит к длительным хроническим заболеваниям печени CLD без выведения вируса. В этом контексте HBV и HCV генерируют белки, подавляющие иммунитет, который должен контролировать хронические инфекции. HBV- и HCV-кодированные белки изменяют экспрессию гена хозяина и клеточный фенотип, что является признаком рака. Эти изменения способствуют независимой от фактора роста пролиферации, сопротивлению к ингибированию роста, инвазии тканей и метастазированию, ангиогенезу, перепрограммированию энергетического метаболизма и устойчивости к апоптозу. Хроническое воспаление также способствует генетической нестабильности в клетках опухоли. Вклад HBV в развитие HCC включает в себя экспрессию гена гепатита Bx (HBx) и, возможно, усеченные пре-S или S-полипептиды карбоксимицина; основной белок и неструктурные (NS) белки NS3 и NS5A HCV способствуют онкогенной трансформации. Изменения в экспрессии гена хозяина, которые способствуют возникновению опухолевого генеза, также, похоже, поддерживают репликацию вируса и/или защиты вирусных гепатоцитов от иммунного опосредованного повреждения и разрушения.
Онкогенные Вирусы
Впервые вирусы связали со злокачественными новообразованиями в своих наблюдениях Эллерман и Банг (1908), которые отметили, что способ передачи лейкемии у домашней птицы напоминает таковой при инфекционной болезни. Раус (1911) показал, что солидная злокачественная опухоль, куриная саркома вызывается вирусом; за это открытие он был запоздало удостоен Нобелевской премии в 1966. Вирусы, вызывающие опухоли у млекопитающих были впервые обнаружены Шоупом, который выделил вирус фибромы кролика в 1932 и вирус папилломы в 1933. Хотя папиллома является доброкачественной опухолью, она может малигнизироваться. Биттнер (1936) предположил, что рак молочной железы у мышей может быть вызван вирусом, передаваемым от матери потомству через грудное молоко. В течение 1950-ых годов было выявлено большое количество вирусов, вызывающих лейкемию у грызунов. Большой интерес был вызван открытием Стюарт и Эдди (1957) вируса полиомы, который при введении новорожденным грызунам мог вызывать развитие большого количество разнообразных опухолей. Трентином (1962) было показано, что введение некоторых типов аденовирусов человека новорожденным хомякам вызывало развитие сарком. Бёркит (1963) выявил специфическое географическое распространение лимфом у африканских детей и предположил, что они могут быть вызваны вирусом, передающимся насекомыми. Выделенный из лимфомы Бёркита вирус Эпштейна-Барр было предложено считать этиологическим агентом лимфомы Бёркита. Много вирусов было выделено из человеческих опухолей или обнаруживалось электромикроскопически в пораженных клетках и тканях, но большинство из них были только "вирусами-пассажирами". присутствующими в поражениях, а не вызывающими их агентами.
Вирусы, которые вызывают опухоли у своих естественных хозяев или у подопытных животных, или же стимулируют злокачественное преобразование в культуре клеток, известны как онкогенныевирусы. Трансформация представляет собой различные изменения, которые сопровождают преобразование нормальной клетки в злокачественную. Трансформация из нормальных в злокачественные клетки является многостадийным процессом, и может быть частичной или полной. Например, некоторые вирусные агенты могут превращать инфицированные клетки в "неумирающие", так что они становятся способными к непрерывному размножению в культуре, без приобретения других особенностей злокачественного развития.
Таблица 3. Онкогенные вирусы
Вирусы лейкозов птиц
Вирусы лекцозов мышей
Вирусы рака молочных желез мышей
Лейкозно-саркоматозные вирусы различных животных
Вирусы Т-клеточной лейкемии человека
Папилломавирусы человека, кроликов и других животных
Обезьяний вирус 40
Вирус контагиозного моллюска
Вирус фибромы Шоупа
Многие типы аденовирусов человека и животных
- 1. Вирус болезни Марека
- 2. Вирус опухоли лягушек Люке
- 3. Epstem-Barrвирус
- 4. Вирусы простого герпеса типов 1 и 2
- 5. Вирус цитомегалии
V. Вирус гепатита B
Преобразованные клетки изменены по форме и теряют способность к "контактному торможению" так что вместо роста одним слоем, они растут скоплениями, одна над другой, формируя "микроопухоли". Участки трансформации могут быть легко выявлены и используются при исследовании онкогенных вирусов, таких как вирус саркомы Рауса.
Около четверти из приблизительно 600 вирусов животных обладают онкогеным потенциалом (Таблица 1). Вирусы, связанные с раковыми образованиями людей описаны в Таблице 2. Среди онкогенных вирусов есть и РНК - и ДНК-содержащие. В то время как все онкогенные РНК-содержащие вирусы (которые раньше назывались онкорнавирусы) принадлежат к единственному семейству (Ретровирусы), онкогенные вирусы встречаются среди всех основных групп ДНК-содержащих вирусов, кроме парвовирусов. Ретровирусы ответственны за естественно возникающую лейкемию и саркому у нескольких видов животных. Среди ДНК-содержащих вирусов, некоторые герпесвирусы вызывают злокачественные опухоли у их естественных хозяев.
Медицинский эксперт статьи


Для объяснения природы рака были предложены две господствующие теории - мутационная и вирусная. В соответствии с первой рак есть результат последовательных мутаций ряда генов в одной клетке, т. е. в его основе лежат изменения, возникающие на генном уровне. Эта теория в законченном виде была сформулирована в 1974 г. Ф. Бернетом: раковая опухоль моноклональна она происходит от одной исходной соматической клетки, мутации в которой вызываются химическими, физическими агентами и вирусами, повреждающими ДНК. В популяции таких мутантных клеток происходит накопление дополнительных мутаций, увеличивающих способность клеток к неограниченному размножению. Однако накопление мутаций требует определенного времени, поэтому рак развивается постепенно, и вероятность появления болезни зависит от возраста.
Вирусно-генетическая теория рака наиболее четко была сформулирована русским ученым Л. А. Зильбером: рак вызывают онкогенные вирусы, они интегрируются в хромосому клетки и создают раковый фенотип. Полному признанию вирусно-генетической теории некоторое время препятствовало то обстоятельство, что многие онкогенные вирусы имеют РНК-геном, поэтому непонятно было, как он интегрируется в хромосому клетки. После того как у таких вирусов была обнаружена обратная транскриптаза, способная воспроизводить из вирионной РНК ДНК-провирус, это препятствие отпало и вирусно-генетическая теория получила признание наряду с мутационной.
Решающий вклад в понимание природы рака внесло открытие в составе онкогенных вирусов гена злокачественности - онкогена и его предшественника, имеющегося в клетках человека, млекопитающих животных и птиц, - протоонкогена.
Протоонкогены - семейство генов, выполняющих в нормальной клетке жизненно важные функции. Они необходимы для регуляции ее роста и размножения. Продуктами протоонкогенов являются различные протеинкиназы, которые осуществляют фосфорилирование клеточных сигнальных белков, а также факторы транскрипции. Последние представляют собой белки - продукты протоонкогенов c-myc, c-fos, c-jun, c-myh и генов-супрессоров клетки.
Существуют два типа онковирусов:
- Вирусы, содержащие онкоген (вирусы one+).
- Вирусы, не содержащие онкогена (вирусы one").
- Вирусы one+ могут утрачивать онкоген, но это не нарушает их нормальной жизнедеятельности. Иначе говоря, сам по себе онкоген вирусу не нужен.
Основное различие между вирусами one+ и one" состоит в следующем: вирус one+, проникнув в клетку, не вызывает ее трансформации в раковую или вызывает крайне редко. Вирусы one", попадая в ядро клетки, трансформируют ее в раковую.
Стало быть, превращение нормальной клетки в опухолевую происходит вследствие того, что онкоген, будучи привнесенным в хромосому клетки, наделяет ее новым качеством, которое позволяет ей размножаться в организме бесконтрольно, образуя клон раковых клеток. Этот механизм превращения нормальной клетки в раковую напоминает транедукцию бактерий, при которой умеренный фаг, интегрируясь в хромосому бактерий, наделяет их новыми свойствами. Это тем более правдоподобно, что онкогенные вирусы ведут себя как транспозоны: они могут интегрироваться в хромосому, перемещаться в ней из одного участка в другой или переходить из одной хромосомы в другую. Суть вопроса заключается в следующем: каким образом протоонкоген превращается в онкоген, когда он взаимодействует с вирусом? Прежде всего необходимо отметить то важное обстоятельство, что у вирусов в связи с высокой скоростью их размножения промоторы работают с гораздо большей активностью, чем промоторы в эукариотных клетках. Поэтому, когда one"-вирус интегрируется в хромосому клетки по соседству с одним из протоонкогенов, он подчиняет работу этого гена своему промотору. Выходя из хромосомы, вирусный геном выхватывает из нее протоонкоген, последний становится составной частью вирусного генома и превращается в онкоген, а вирус из one - в one+-вирус. Интегрируясь в хромосому другой клетки, такой уже onc"-вирус одновременно трансдуцирует в нее и онкоген со всеми последствиями. Таков наиболее частый механизм образования онкогенных (one+)-вирусов и начала превращения нормальной клетки в опухолевую. Возможны и другие механизмы превращения протоонкогена в онкоген:
- транслокация протоонкогена, в результате которой протоонкоген оказывается по соседству с сильным вирусным промотором, который берет его под свой контроль;
- амплификация протоонкогена, в результате которой количество копий его возрастает, как и количество синтезируемого продукта;
- превращение протоонкогена в онкоген происходит вследствие мутаций, вызываемых физическими и химическими мутагенами.
Таким образом, основные причины превращения протоонкогена в онкоген следующие:
- Включение протоонкогена в геном вируса и превращение последнего в one+-вирус.
- Попадание протоонкогена под контроль сильного промотора либо в результате интеграции вируса, либо вследствие транслокации блока генов в хромосоме.
- Точечные мутации в протоонкогене.
Амплификация протоонкогенов. Последствиями всех этих событий могут быть:
- изменение специфичности или активности белкового продукта онкогена, тем более, что очень часто включение в геном вируса протоонкогена сопровождается мутациями протоонкогена;
- утрата клеточно-специфической и временной регуляции этого продукта;
- увеличение количества синтезируемого белкового продукта онкогена.
Продукты онкогенов - также протеинкиназы и факторы транскрипции, поэтому нарушения активности и специфичности протеинкиназ и рассматриваются как начальные пусковые механизмы превращения нормальной клетки в опухолевую. Поскольку семейство протоонкогенов состоит из 20-30 генов, то и семейство онкогенов включает в себя, очевидно, не более трех десятков вариантов.
Однако злокачественность таких клеток зависит не только от мутаций протоонкогенов, но и от изменений во влиянии на работу генов со стороны генетической среды в целом, характерной для нормальной клетки. Такова современная генная теория рака.
Таким образом, первичная причина превращения нормальной клетки в злокачественную - мутации протоонкогена или попадание его под контроль мощного вирусного промотора. Различные внешние факторы, индуцирующие образование опухолей (химические вещества, ионизирующая радиация, УФ-облучение, вирусы и др.). действуют на одну и ту же мишень - протоонкогены. Они имеются в хромосомах клеток каждого индивидуума. Под влиянием указанных факторов включается тот или иной генетический механизм, который приводит к изменению функции протоонкогена, а это, в свою очередь, дает начало перерождению нормальной клетки в злокачественную.
Раковая клетка несет на себе чужеродные вирусные белки или собственные измененные белки. Она распознается Т-цитотоксическими лимфоцитами и при участии остальных механизмов иммунной системы уничтожается. Кроме Т-цитотоксических лимфоцитов раковые клетки распознаются и уничтожаются другими киллерными клетками: NK, Pit-клетками, В-киллерами, а также К-клетками, чья цитотоксическая активность зависит от антител. В качестве К-клеток могут функционировать полиморфно-ядерные лейкоциты; макрофаги; моноциты; тромбоциты; мононуклеарные клетки лимфоидной ткани, лишенные маркеров Т- и В-лимфоцитов; Т-лимфоциты, имеющие Fc-рецепторы для IgM.
Противоопухолевым действием обладают интерфероны и некоторые другие биологически активные соединения, образуемые иммунокомпетентными клетками. В частности, раковые клетки распознаются и разрушаются рядом цитокинов, в особенности такими, как фактор некроза опухоли и лимфотоксин. Они представляют собой родственные белки с широким спектром биологической активности. Фактор некроза опухоли (ФНО) - один из главных медиаторов воспалительных и иммунных реакций организма. Он синтезируется различными клетками иммунной системы, главным образом макрофагами, Т-лимфоцитами и купферовскими клетками печени. ФНОа был обнаружен в 1975 г. Е. Карсвеллом с сотрудниками; это полипептид с м. м. 17 кД. Он обладает сложным плейотропным действием: индуцирует экспрессию молекул МНС класса II у иммунокомпетентных клеток; стимулирует продукцию интерлейкинов IL-1 и IL-6, простагландина ПГЕ2 (он служит негативным регулятором механизма секреции ФНО); оказывает хемотаксическое действие в отношении зрелых Т-лимфоцитов и т. п. Важнейшая физиологическая роль ФНО - модуляция роста клеток в организме (рострегулирующая и цитодифференцирующая функции). Кроме того, он селективно подавляет рост злокачественных клеток и вызывает их лизис. Предполагается, что ростмодулирующая активность ФНО может быть использована в противоположном направлении, а именно, для стимуляции роста нормальных и подавления роста злокачественных клеток.
Лимфотоксин, или ФНО-бета, - белок с м. м. около 80 кД, синтезируется некоторыми субпопуляциями Т-лимфоцитов, также обладает способностью лизировать клетки-мишени, несущие чужеродные антигены. Способностью активировать функции NK-клеток, К-клеток, макрофагов, нейтрофильных лейкоцитов обладают и другие пептиды, в частности пептиды, являющиеся фрагментами молекул IgG, например тафтеин (цитофильный полипептид, выделенный из СН2-домена), фрагменты Fab, Fc и др. Только благодаря постоянному взаимодействию всех иммунокомпетентных систем обеспечивается противоопухолевый иммунитет.
Большинство людей не болеет раком не потому, что у них не возникают мутантные раковые клетки, а потому, что последние, возникнув, своевременно распознаются и уничтожаются Т-цитотоксическими лимфоцитами и другими звеньями иммунной системы раньше, чем успевают дать злокачественное потомство. У таких людей противоопухолевый иммунитет работает надежно. Напротив, у больных раком мутантные клетки вовремя не распознаются или не уничтожаются иммунной системой, а беспрепятственно и бесконтрольно размножаются. Следовательно, рак - это следствие иммунодефицита. Какое звено иммунитета при этом страдает, - необходимо выяснить, чтобы наметить более эффективные пути борьбы с заболеванием. В этом плане большое внимание уделяется разработке способов биотерапии рака, основанных на комплексном и последовательном использовании модуляторов биологической и иммунологической реактивности, т. е. химических веществ, синтезируемых иммунокомпетентными клетками, которые способны модифицировать реакции взаимодействия организма с опухолевыми клетками и обеспечивать противоопухолевый иммунитет. С помощью таких модификаторов иммунологической реактивности открывается возможность воздействия как в целом на иммунную систему, так и избирательно на ее отдельные механизмы, в том числе контролирующие образование факторов активации, пролиферации, дифференцировки, синтез интерлейкинов, факторов некроза опухолей, лимфотоксинов, интерферонов и т. п., чтобы устранить состояние иммунодефицита при раке и повысить эффективность его лечения. Уже описаны случаи излечения миеломы человека с помощью лимфокинактивированных киллеров и интерлейкина-2. В экспериментальной и клинической иммунотерапии рака наметились следующие направления.
- Введение в опухолевые ткани активированных клеток иммунной системы.
- Использование лимфо- или (и) монокинов.
- Применение иммуномодуляторов бактериального происхождения (наиболее эффективны ЛПС и производные пептидогликана) и индуцированных ими продуктов, в частности ФНО.
- Использование противоопухолевых антител, в том числе моноклональных.
- Комбинированное использование различных направлений, например первого и второго.
Перспективы использования модуляторов иммунологической реактивности для биотерапии рака необычайно широки.

[1], [2], [3], [4], [5], [6], [7], [8], [9], [10]
Читайте также:
- Вирус цитадель что это
- Вирус рвота температура красное горло
- Почему ребенок вялый после ротавируса
- Вирус понос температура что делать
- Что такое калькулятор гриппа