
Что такое стратегия вирусного генома
Лекция 5
ТРАНСКРИПЦИЯ
Транскрипция — это переписывание ДНК на РНК по законам генетического кода. Это означает, что РНК состоит из нуклеотидных последовательностей, комплементарных ДНК. Нити ДНК в участке транскрипции разделяются и функционируют как матрицы, к которым присоединяются комплементарные нуклеотиды благодаря спариванию комплементарных оснований (аденин связывается с тимином, урацил — с аденином, гуанин — с цитозином и цитозин - с гуанином). Транскрипция осуществляется с помощью специального фермента — РНК-полимеразы, который связывает нуклеотиды путем образования 3'-5'-фосфодиэфирных мостиков. Такое связывание происходит лишь в присутствии ДНК-матрицы.
Продуктами транскрипции в клетке являются иРНК. Сама клеточная ДНК, являющаяся носителем генетической информации, не может непосредственно программировать синтез белка. Передачу генетической информации от ДНК к рибосомам осуществляет РНК-посредник. На этом основана центральная догма молекулярной биологии, которая выражается следующей схемой:
ДНК - (транскрипция) –и РНК – (трансляция) - белок
где стрелки показывают направление переноса генетической информации.
Реализация генетической информации у вирусов. Стратегия вирусного генома в отношении синтеза иРНК у разных вирусов различна. У ДНК-содержащих вирусов иРНК синтезируется на матрице одной из нитей ДНК. Формула переноса генетической информации у них такая же, как и в клетке.
ДНК-содержащие вирусы, репродукция которых происходит в ядре, используют для транскрипции клеточную полимеразу. К этим вирусам относятся паповавирусы, аденовирусы, вирусы герпеса. ДНК-содержащие вирусы, репродукция которых происходит в цитоплазме, не могут использовать клеточный фермент, находящийся в ядре. Транскрипция их генома осуществляется вирусспецифическим ферментом — ДНК-полимеразой, которая проникает в клетку в составе вируса. К этим вирусам относятся вирусы оспы и иридовирусы.
К этой группе вирусов относятся пикорнавирусы, тогавирусы, коронавирусы. У них нет необходимости в акте транскрипции для синтеза вируспецифических белков. Поэтому транскрипцию как самостоятельный процесс у этих вирусов не выделяют. Иначе обстоит дело у вирусов, геном которых не может выполнять функцию иРНК. В клетке синтезируется комплементарная геному РНК, которая и является информационной. Передача генетической информации у этих вирусов осуществляется по схеме:
РНК– иPHK - белок
У этих вирусов транскрипция выделена как самостоятельный процесс в инфекционном цикле. К ним относятся две группы вирусов животных.
2. Вирусы, геном которых представлен двунитчатой РНК (диплорнавирусы). Среди вирусов животных к ним относятся реовирусы.
В клетке нет фермента, который может полимеризовать нуклеотиды на матрице РНК. Эту функцию выполняет вирусспецифический фермент — РНК-полимераза, или транскриптаза, которая находится в составе вирусов и вместе с ними проникает в клетку.
Среди РНК-содержащих вирусов животных есть семейство ретровирусов, которые имеют уникальный путь передачи генетической информации. РНК этих вирусов переписывается на ДНК, ДНК интегрирует с клеточным геномом и в его составе переписывается на РНК, которая обладает информационными функциями. Путь передачи генетической информации в-этом случае осуществляется по более сложной схеме:
РНК - ДНК - иРНК - белок
Ферменты, транскрибирующие вирусный геном. Транскрипция ряда ДНК-содержащих вирусов — паповавирусов, аденовирусов, вирусов герпеса, парвовирусов, гепадна-вирусов осуществляется в ядре клетки, и в этом процессе широко используются механизмы клеточной транскрипции — ферменты транскрипции и дальнейшей модификации транскриптов. Транскрипция этих вирусов осуществляется клеточной РНК-полимеразой II — ферментом, который осуществляет транскрипцию клеточного генома. Однако особая группа транскриптов аденовируса синтезируется с помощью другого клеточного фермента — РНК-полимеразы III. У двух других семейств ДНК-содержащих вирусов животных - вирусов оспы и иридовирусов — транскрипция происходит в цитоплазме. Поскольку в цитоплазме нет клеточных полимераз, транскрипция этих вирусов нуждается в специальном вирусном ферменте — вирусной РНК-полимеразе. Этот фермент является структурным вирусным белком.
У РНК-содержащих вирусов транскрипция осуществляется вирусспецифическими транскриптазами, т. е. ферментами, закодированными в вирусном геноме. Вирусспецифические транскриптазы могут быть как структурными белками, входящими в состав вириона (эндогенная транскриптаза), так и неструктурными белками, которые синтезируются в зараженной клетке, но не включаются в вирион.
Транскрипция в зараженной клетке. Синтез комплементарных РНК на родительских матрицах с помощью родительской транскриптазы носит название первичной транскрипции в отличие от вторичной транскрипции, происходящей на более поздних стадиях инфекционного цикла на вновь синтезированных, дочерних матрицах, с помощью вновь синтезированной транскриптазы. Большая часть иРНК в зараженной клетке является продуктом вторичной транскрипции.
Транскриптивные комплексы. У сложно устроенных РНК-содержащих вирусов животных транскрипция происходит не на матрице голой РНК, а в составе вирусных нуклеокапсидов или сердцевин (транскриптивные комплексы). Связанные с геномом капсидные белки не только не препятствуют транскрипции, но и необходимы для нее, обеспечивая правильную конформацию тяжа РНК, защиту его от клеточных протеаз, связь отдельных фрагментов генома друг с другом, а также регуляцию транскрипции.
Регуляция транскрипции. Транскрипция вирусного генома строго регулируется на протяжении инфекционного цикла. Регуляция осуществляется как клеточными, так и вирусспецифическими механизмами. У некоторых вирусов, в основном ДНК-содержащих, существует три периода транскрипций — сверхранняя, ранняя и поздняя. К этим вирусам относятся вирусы оспы, герпеса, паповавирусы, аденовирусы. В результате сверхранней и ранней транскрипции избирательно считываются сверхранние и ранние гены с образованием сверхранних или ранних иРНК. При поздней транскрипции считывается другая часть вирусного генома — поздние гены, с образованием поздних иРНК. Количество поздних генов обычно превышает количество ранних генов. Многие сверхранние гены являются генами для неструктурных белков — ферментов и регуляторов транскрипции и репликации вирусного генома. Напротив, поздние гены обычно являются генами для структурных белков. Обычно при поздней транскрипции считывается весь геном, но с преобладанием транскрипции поздних генов.
Фактором регуляции транскрипции у ядерных вирусов является транспорт транскриптов из ядра в цитоплазму, к месту функционирования иРНК — полисомам.
У РНК-содержащих вирусов синтез транскриптов также строго контролируется в отношении как количества каждого класса транскриптов, так и периода инфекции, когда определенные транскрипты синтезируются с максимальной скоростью. На ранней стадии инфекции преимущественно синтезируются транскрипты двух генов вируса гриппа — NP и NS, на поздней стадии инфекции — транскрипты генов М, НА и NA. Остальные три гена для Р-белков синтезируются примерно с одинаковой скоростью на протяжении всего периода инфекции. У реовирусов на ранней стадии инфекции преимущественно транскрибируется 4 из 10 фрагментов генома и лишь на поздней стадии транскрибируется весь геном. Однако если поместить геном вируса в бесклеточную РНК-синтезирующую систему, будет происходить равномерная транскрипция всех 10 фрагментов генома. Эти факты говорят о жестком контроле транскрипции со стороны клетки-хозяина и возможном наличии специфических клеточных регуляторов.
ТРАНСЛЯЦИЯ
Синтез белка в клетке происходит в результате трансляции иРНК. Трансляцией называется процесс перевода генетической информации, содержащейся в иРНК, на специфическую последовательность аминокислот. Иными словами, в процессе трансляции осуществляется перевод 4-буквенного языка азотистых оснований на 20-буквенный язык аминокислот.
Рибосомы. Синтез белка в клетке осуществляется на рибосоме. Рибосома состоит из двух субъединиц, большой и малой, малая субъединица примерно в два раза меньше большой. Обе субъединицы содержат по одной молекуле рибосомальной РНК и ряд белков. Рибосомальные РНК синтезируются в ядре на матрице ДНК с помощью РНК-полимеразы. В малой рибосомальной субъединице есть канал, в котором находится информационная РНК. В большой рибосомальной субъединице есть две полости, захватывающие также малую рибосомальную субъединицу. Одна из них содержит аминоацильный центр (А-центр), другая — пептидильный центр (П-центр).
Фазы трансляции. Процесс трансляции состоит из трех фаз: 1) инициации, 2) элонгации и 3) терминации.
Вначале с иРНК связывается малая рибосомальная субъединица. К комплексу иРНК с малой рибосомальной субъединицей присоединяются другие компоненты, необходимые для начала трансляции. Их по крайней мере три в прокариотической клетке и более девяти в эукариотической клетке. Инициаторные факторы определяют узнавание рибосомой специфических иРНК и, таким образом, являются определяющим фактором в дискриминации между различными иРНК, присутствующими в клетке, как правило, в избыточном количестве.
В результате формируется комплекс, необходимый для инициации трансляции, который называется инициаторным комплексом. В инициаторный комплекс входят: 1) иРНК; 2) малая рибосомальная субъединица; 3) аминоацил-тРНК, несущая инициаторную аминокислоту; 4) инициаторные факторы; 5) несколько молекул ГТФ.
В рибосоме осуществляется слияние потока информации с потоком аминокислот. Аминоацил-тРНК входит в А-центр большой рибосомальной субъединицы, и ее антикодон взаимодействует с кодоном иРНК, находящейся в малой рибосомальной субъединице. При продвижении иРНК на один кодон тРНК перебрасывается в пептидильный центр, и ее аминокислота присоединяется к инициаторной аминокислоте с образованием первой пептидной связи. Свободная от аминокислоты тРНК выходит из рибосомы и может опять функционировать в транспорте специфических аминокислот.
Терминация трансляции. Терминация трансляции происходит в тот момент, когда рибосома доходит до терминирующего кодона в составе иРНК. Трансляция прекращается, и полипептидная цепь освобождается из полирибосомы. После окончания трансляции полирибосомы распадаются на субьединицы, которые могут войти в состав новых полирибосом.
В основу современной классификации положены следующие основные критерии:
1) тип нуклеиновой кислоты (РНК или ДНК), ее структура (количество нитей);
2) наличие липопротеидной оболочки;
3) стратегия вирусного генома;
4) размер и морфология вириона, тип симметрии, число капсомеров;
5) феномены генетических взаимодействий;
6) круг восприимчивых хозяев;
7) патогенность, в том числе патологические изменения в клетках и образование внутриклеточных включений;
8) географическое распространение;
9) способ передачи;
10) антигенные свойства.
На основании перечисленных признаков вирусы делятся на семейства, подсемейства, роды и типы. Деление на семейства произведено по критериям, изложенным в пунктах 1 и 2, деление на роды и типы – на основании нижеперечисленных признаков. Схематически строение семейств вирионов, поражающих позвоночных, приведено на рисунке 5 Дополнительно выделены еще 2 семейства: Gepadnaviridae и Flaviviridae (выделенные из семейства Togaviridae).
Современная классификация вирусов человека и животных охватывает более 4/5 всех известных вирусов, которые распределены в 19 семейств, из них 7 – ДНК-содержащих и 12 – РНК-содержащих вирусов. Некоторые допускаются привычные латинизированные обозначения, цифры при обозначении типов, сокращения, буквы и их сочетания.
В начале 1940-х годов стало более или менее ясно, что вирусы, как и другие живые организмы, содержат нуклеиновые кислоты. Однако, особенность строения вирусного генома заключается в том, что наследственная информация может быть записана как на ДНК, так и РНК в зависимости от типа вируса. Уникальное свойство вирусной РНК хранить наследственную информацию впервые было продемонстрировано Гирером, Шраммом и Френкель-Конратом при изучении инфекционности РНК вируса табачной мозаики (ВТМ). Было показано, что очищенные препараты РНК ВТМ сохраняют инфекционность при полном отсутствии белка. Это открытие противоречило всеобщему убеждению, что единственная роль РНК заключается в передаче информации от ДНК к белку. В настоящее время способность РНК служить хранилищем генетической информации уже ни у кого не вызывает сомнений.
Следует учитывать, что наличие одного вида нуклеиновой кислоты является характеристикой вириона, но не вируса. В жизненном цикле ДНК-содержащих вирусов геномная нуклеиновая кислота транскрибируется, образуя РНК. Присутствие у ДНК-содержащих вирусов (вирус осповакцины) ДНК-зависимой РНК-полимеразы было показано в 1967 г. Катес и МакАусланом. РНК-содержащие вирусы транскрибируют свой геном с использованием РНК-зависимой РНК-полимеразы, впервые обнаруженной у реовирусов в 1968 г. Целый ряд РНК-содержащих вирусов имеют в жизненном цикле стадию обратной транскрипции и синтезируют ДНК на матрице РНК с помощью фермента обратной транскриптазы (РНК-зависимая ДНКполимераза или ревертаза). Открытие этого фермента в составе онкогенных РНК-содержащих вирусов сделано Балтимором в 1970 г.
Примерно 20 % вирусов имеют ДНК-геном, 80 % – РНК-геном.
По своей химической природе нуклеиновые кислоты вирусов не отличаются от нуклеиновых кислот клеток (организмов) и представляют собой полинуклеотидные цепи, образованные чередованием четырех дезоксирибонуклеотидов в случае ДНК или рибонуклеотидов в случае РНК, соединенных фосфодиэфирными связями. Нуклеотид представляет собой азотистое основание (аденозин (А), гуанозин (G), цитидин (C), тимидин (T) или уридин (U) в случае РНК), связанное с дезоксирибозой (в ДНК), или рибозой (в РНК) гликозидной связью. Фосфорная кислота в нуклеиновых кислотах присоединяется сложноэфирной связью к 3’– и 5’-ОН группам сахаров смежных нуклеотидов. Отличительной особенностью ДНК целого ряда вирусов бактерий является наличие метилированных оснований (5’-метилцитозина, 6’метиламинопурина), которые могут входить в состав ДНК в качестве минорных или мажорных оснований. Так, ДНК фагов fd и ?Х174 (колифаги) содержит 1-2 метилированных основания, а в ДНК фага Х12, лизирующего морскую бактерию Xantomonas oryza, вообще нет обычного цитозина, который полностью замещен 5метилцитозином. Источником происхождения таких оснований является энзиматическое метилирование уже синтезированной цепи ДНК. Данный процесс осуществляют вирусспецифические метилазы, которые используют в качестве донора метильных групп S-аденозилметионин клетки хозяина.
Вирусные нуклеиновые кислоты характеризуются поразительным разнообразием форм. Вирусный геном может быть представлен как однонитчатыми, так и двунитчатыми молекулами РНК и ДНК. ДНК может быть как линейной, так и кольцевой молекулой (таблица 1), РНК – как непрерывной, так и фрагментированной и кольцевой молекулой (таблица 2, рисунок 4).
Таблиц 1 – Виды ДНК-геномов вирусов

Таблица 2 – Виды РНК-геномов вирусов

При обсуждении вопроса организации генома вирусов необходимо выделить ряд его характерных отличий от геномов организмов.
Размеры . Если ориентировочно учесть, что геном эукариот имеет размер 4?10 9 п.н. и длину, достигающую 1,5-2 метра, а геном прокариот – 6?10 6 п.н., то размеры геномов вирусов значительно меньше. Так, размер генома крупных ДНК-содержащих вирусов составляет только 2-4?10 5 п. н. (200-450 т.п.н. у поксвирусов и вирусов герпеса), минимальные вирусы имеют геном длиной 1 мкм и состоящий из 1,2 т.п.н. Самый маленький геном среди вирусов, поражающих человека, имеет вирус гепатита В (3,2 т.п.н.).
Экономичность . Размеры геномов вирусов определяются емкостью капсида вириона. В связи с этим, вирусы очень экономно хранят генетическую информацию, что проявляется отсутствием многократно повторяющихся генов и, часто, наличием перекрывающихся открытых рамок считывания.
Наличие двух типов геномов . Носителем генетической информации у вирусов может быть как ДНК, так и РНК.
Многообразие структурных форм ДНК и РНК . Природа как бы испробовала на вирусах все возможные варианты структурной организации нуклеиновых кислот, которые представлены на рисунок 4.

Рисунок 4 – Схема, иллюстрирующая виды нуклеиновых кислот вирусов
Разнообразие стратегий репликации, основанных на ДНК-зависимом синтезе ДНК, РНК-зависимом синтезе РНК и РНК-зависимом синтезе ДНК.
Изменчивость РНК. Полимеразы, катализирующие репликацию РНК, и обратная транскриптаза имеют минимальные возможности для исправления ошибок синтеза. В результате, частота возникновения ошибок при синтезе РНК приблизительно в 10 тысяч раз выше, чем при репликации ДНК, и она зависит от числа нуклеотидов, составляющих вирусный геном. Это означает, что геном любой индивидуальной частицы РНК– содержащего вируса будет содержать одну или несколько мутаций, отличающих его от последовательности дикого типа данной вирусной разновидности. Этот простой факт имеет далеко идущие последствия для биологии и эволюции РНК-содержащих вирусов, потому что потомство РНК-вируса (природное или лабораторное) представляет собой не совокупность однородных двойников, а скорее молекулярный рой родственных нуклеотидных последовательностей, сгруппированных в месте синтеза последовательностей. Этот молекулярный рой или “квази-разновидность” обеспечивает источник фенотипических вариантов, которые могут быстро ответить на изменяющееся давление естественного отбора. Как следствие, РНК-содержащие вирусы могут эволюционировать в миллион раз быстрее чем, ДНК-организмы. В тоже время высокая изменчивость РНК не обеспечивает быструю эволюцию РНК-содержащих вирусов, так, как размеры генома вирусов налагают верхние пределы на высокую норму ошибок полимеразы. Комбинация уровня репликационных ошибок и размера генома определяют “порог ошибки”, выше которого вирус не может поддерживать целостность последовательности квазиразновидностей. В результате, немногие РНК вирусы имеют размер генома более 30 килобаз (kb), чаще всего он колеблется в пределах 5-15 kb.
Принимая во внимание, что генетически разнообразное потомство может нести летальные мутации, что снижает потенциал для быстрого эволюционного ответа, РНК-геномы этого размера сбалансированы ниже их порогов ошибки.
Детально рассмотрим особенности организации вирусных ДНК и РНК.
Молекулярная масса вирусных ДНК варьирует в широких пределах от 1?10 6 до 250?10 6 . Самые большие вирусные геномы содержат несколько сотен генов, а самые маленькие содержат информацию, достаточную для синтеза лишь нескольких белков.
В геномах, представленных двунитчатыми ДНК, информация обычно закодирована на обеих нитях ДНК. Это свидетельствует о максимальной экономии генетического материала у вирусов, что является неотъемлемым свойством их как генетических паразитов. В связи с этим оценка генетической информации не может быть проведена по молекулярной массе молекул.
Хотя в основном структура ДНК уникальна, т. е. большинство нуклеотидных последовательностей встречаются лишь по одному разу, однако на концах молекул имеются повторы, когда в концевом фрагменте линейной ДНК повторяется ее начальный участок. Повторы могут быть прямыми и инвертированными.
Способность к приобретению кольцевой формы, которая потенциальнозаложена в концевых прямых и, инвертированных повторах, имеет большое значение для вирусов. Кольцевая форма обеспечивает устойчивость ДНК к экзонуклеазам. Стадия образования кольцевой формы обязательна для процесса интеграции ДНК с клеточным геномом. Наконец, кольцевые формы представляют собой удобный и эффективный способ регуляции транскрипции и репликации ДНК.
Инфекционный процесс при заражении этими вирусами возникает лишь при проникновении в клетку частиц обоих типов.
Из нескольких сотен известных в настоящее время вирусов человека и животных РНК-геном содержит около 80 % вирусов. Способность РНК хранить наследственную информацию является уникальной особенностью вируса.
У просто организованных и некоторых сложно организованных вирусов вирусная РНК в отсутствие белка может вызвать, инфекционный процесс. Впервые инфекционная активность РНК вируса табачной мозаики была продемонстрирована X. Френкель-Конратом и соавт. в 1957 г. и А. Гирером и Г. Шраммом в 1958 г. Впоследствии положение об инфекционной активности РНК было перенесено на все РНК-содержащие вирусы, однако долголетние усилия доказать это для таких вирусов, как вирусы гриппа, парамиксовирусы, рабдовирусы (так называемые минуснитевые вирусы), оказались бесплодными: у этих вирусов инфекционной структурой являются не РНК, а комплекс РНК с внутренними белками. Таким образом, геномная РНК может обладать инфекционной активностью в зависимости от своей структуры.
Структура вирусных РНК чрезвычайно разнообразна. У вирусов обнаружены однонитчатые и двунитчатые, линейные, фрагментированные и кольцевые РНК. РНК-геном в основном является гаплоидным, до геном ретровирусов – диплоидный, т.е. состоит из двух идентичных молекул РНК (таблица 2).
Многообразие видов РНК-геномов расширяется за счет существования последовательностей, отличающихся направлением связей сахаро-фосфатного остова. Однонитевые РНК могут иметь позитивную полярность – (+)РНК, негативную полярность – (-)РНК или могут быть представлены обоюдозначащей цепью – (+/-)РНК (амбисенс стратегия кодирования). В свою очередь, РНК позитивной полярности могут иметь разную структурную организацию. Являясь матричной РНК, могут иметь на 5’– конце кэп (7-метилгуанозин), а на 3’-конце – поли-А последовательность; могут не иметь кэпа или поли-А; могут иметь на 5’-конце геномный белок; могут иметь на 3’-конце тРНК-подобную или шпильковую структуру.

Рисунок 5 – Вторичная структура вирусных РНК
В основном однонитчатые РНК являются линейными молекулами, однако РНК-фрагменты буньявирусов обнаружены в виде кольцевой формы. Кольцевая форма возникает за счет образования водородных связей между концами молекул.
Двунитчатые РНК. Этот необычный для клетки тип нуклеиновой кислоты, впервые обнаруженный у реовирусов, широко распространен среди вирусов животных, растений и бактерий. Вирусы, содержащие подобный геном, называют диплорнавирусы.
Общей особенностью диплорнавирусов является фрагментированное состояние генома. Так, геном реовирусов состоит из 10 фрагментов, ротавирусов – из 11 фрагментов.
Как и в других живых системах, у вирусов соответствие между аминокислотной последовательностью белка и нуклеотидной последовательностью геномной нуклеиновой кислоты устанавливается с помощью универсального вырожденного генетического кода, где кодирующей единицей является триплет нуклеотидов (кодон). В вирусных системах используются те же наборы кодоновых значений, что и в бактериальных, архейных и эукариотических системах. Однако у вирусов генетическая информация может храниться, как в смысловой полинуклеотидной цепи, так и в последовательности матричной цепи.
Для сохранения генетической информации в окружающей среде и передачи ее новому поколению вирусы упаковывают геномные нуклеиновые кислоты в белковый капсид и часто в суперкапсид (липидсодержащая оболочка), формируя внеклеточную форму вируса – вирион. Как правило, вирионы, попадая в клетку, обеспечивают продуктивный инфекционный цикл, давая вирусное потомство. Однако целый ряд так называемых интегративных вирусов встраивают свой геном в хромосомы хозяина, в том числе клеток зародышевой линии, обеспечивая длительное сохранение генетической информации вируса в ряду поколений хозяина.
Несмотря на микроскопические размеры, нуклеиновые кислоты вирусов несут информацию не только о капсидных белках, но и о ферментах, необходимых для синтеза ДНК, РНК и их модификации, для синтеза РНК транскриптов и их процессинга, для обеспечения синтеза белков и их посттрансляционной модификации и воздействия на биосинтетические процессы клетки-хозяина. Данный факт объясняется наличием разнообразнейших механизмов увеличения генетической информации. Так, к способам увеличения информационной емкости вирусного генома являются: 1) двукратное считывание одной и той же иРНК, но с другого инициирующего кодона; 2) сдвиг рамки трансляции; 3) сплайсинг; 4) транскрипция с перекрывающихся областей ДНК и др.
Так, генетический материал мелкого бактериофага ?Х174 представлен одноцепочечной ДНК и состоит всего из 9 генов, продукты которых хорошо изучены. ДНК, необходимая для кодирования этих продуктов, должна состоять минимум из 6078 нуклеотидов. На самом же деле хромосома фага состоит из 5374 нуклеотидов. Этот парадокс был разрешен после проведения в 1978 г. группой Ф.Сенгера полного секвенирования ДНК этого фага. Оказалось, что кодирующие последовательности двух генов (В и Е) локализованы внутри кодирующих последовательностей двух других генов (А и D). При этом рамка считывания в каждом случае оказывалась сдвинутой на одну пару нуклеотидов. Например, в определенном участке внутри гена D находится последовательность, которая в полипептиде D кодирует последовательность Валин-тирозин-глицин-треонин. Рамка считывания гена Е смещена вправо на один нуклеотид от рамки считывания гена D. Поэтому триплет ATG распознается РНК-полимеразой как стартовый и в полипептиде Е появляется формилметионин, за которым последует Валин, кодируемый триплетом GTA, и т.д.

Сходным образом кодирующая последовательность гена В оказывается внутри кодирующей последовательности гена А. В результате сдвига рамки считывания кодируемые перекрывающимися генами полипептиды полностью отличаются друг от друга по последовательностям аминокислот. Вместе с тем в случае замены или делеции одного нуклеотида инактивируются сразу два гена.
В ходе эволюции вирусов сложилось несколько стратегий, обеспечивающих: а) организацию вирусных геномов и их кодирующую функцию; б) экспрессию вирусных генов; в) репликацию вирусных геномов и г) сборку и созревание вирусного потомства. Прежде чем рассмотреть данные положения, следует напомнить, что фундаментальные биологические особенности вирусов делают реализацию их генетической информации процессом, не имеющим себе аналогов среди других форм жизни, включая микоплазмы, риккетсии и хламидии. Во-первых, это обусловлено тем, что вирусы содержат один тип нуклеиновой кислоты — РНК или ДНК. Во-вторых, у вирусов отсутствует собственный аппарат для реализации своей генетической информации. Вирусы не имеют ферментов, участвующих в производстве энергии, и белоксинтезирующих систем. Поэтому вирус должен представить информационную РНК, которую клетка должна распознавать и транслировать. Пути образования мРНК в процессе репликации РНК-вирусов представлены на рисунке. В клетке нет ферментов для транскрипции мРНК с вирусного РНК-генома, а в цитоплазме нет ферментов для транскрипции вирусной ДНК.
В связи с этим клеточную транскриптазу для синтеза вирусных мРНК могут использовать только вирусы, содержащие ДНК и способные проникать в ядро. Все другие вирусы вынуждены создавать собственные ферменты для синтеза мРНК.
Почти все РНК-геномы относят к линейным молекулам. Некоторые из них, например, геномы пикорнавирусов, содержат на 5'-конце РНК ковалентно связанные полипептиды. Геномы одноцепочечных РНК-вирусов могут быть либо монолитными (пикорна-, тога-, парамиксо-, рабдо-, корона-, ретровирусы), либо сегментированными (ортомиксо-, арена- и буньявирусы). Среди РНК вирусов с двухцепочечным сегментированным геномом наиболее изучены реовирусы.
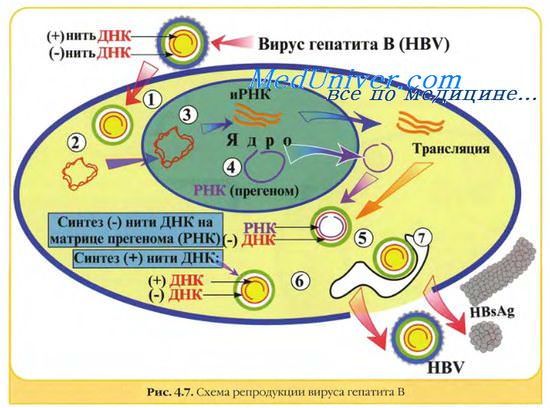
Все известные ДНК-вирусы позвоночных имеют монолитный геном. Все они, за исключением парвовирусов, имеют полностью или частично двухспи-ральный геном. Отдельные представители парвовирусов содержат линейную оцДНК; у других (например, у аденоассоциированных вирусов) комплементарные цепи ДНК находятся в разных вирионах. ДНК папиллома- и полиомавиру-сов — кольцевая и сверхспиральная, в то время как ДНК герпесвирусов, аденовирусов и осповирусов — линейная.
ДНК вируса гепатита В представляет собой кольцевую двуспиральную молекулу, на обеих цепях которой в разных областях обнаружены большой и малый одноцепочечные участки.
Репликация большинства ДНК-вирусов включает механизмы, которые хорошо известны в клеточной биологии: транскрипция мРНК на двуцепочечной ДНК и репликация ДНК. Для РНК-вирусов ситуация является уникальной и связана с генетической информацией, кодируемой РНК. РНК-вирусы с различным типом генома связаны с необходимостью использовать различные пути синтеза мРНК. Кроме (+) оцРНК вирусов, у которых геномная РНК функционирует как мРНК, все другие типы вирусной РНК, прежде всего, должны быть транскрибированы в мРНК. Так как клетки эукариотов не содержат РНК-зависимой РНК-полимеразы, (-) оцРНК вирусы и дцРНК вирусы должны содержать вирионную РНК-зависимую РНК-полимеразу
Эукариотические клетки не могут транслировать полицистронные мРНК частями в виде нескольких индивидуальных белков. ДНК-вирусы расщепляют свои полицисторные РНК-транскрипты на моноцисторные мРНК, используя клеточный механизм.
Большинство РНК-вирусов, размножающихся в цитоплазме, не имеют доступа к ферментам, осуществляющим РНК-процессинг и сплайсинг в ядре, и вынуждены по-разному решать эту проблему. У вирусов с сегментированным геномом каждый фрагмент является отдельным геном. Другие вирусы имеют полицистронный геном, но образуют моноцистронные РНК-транскрипты, разделяя окончание и реинициацию транскрипции.
Некоторые вирусы имеют полицистронную РНК, которая транслируется в полипротеин, который затем расщепляется протеолитически с образованием конечных продуктов.
Анализ данных о молекулярных механизмах репликации вирусов позвоночных позволил установить сходство и различия в способах реализации генетической информации вирусов, относящихся к различным таксономическим группам. Благодаря этому оказалось возможным выделить основные типы стратегии экспресии и репликации вирусных геномов. Семейства вирусов, реплицирующиеся по сходному пути, объединены в один тип. Репликация вирусов каждого типа начинается с инициирующих единиц, которыми являются проникшие в клетку вирусные геномы: вирионные РНК или ДНК или вирионные нуклеопротеины.
Вирусы, геном которых может служить в качестве мРНК, принято называть вирусами с позитивным (+) геномом. Вирусы с негативным (-) РНК геномом неспособность функционировать в качестве мРНК компенсируют наличием вирионной транскриптазы.
Читайте также: