
Факторы влияющие на чувствительность животных к вирусам
Вирусы являются внутриклеточными инфекционными агентами. Весь репликативный цикл вируса осуществляется с исполь зованием метаболических и генетических ресурсов клеток. Поэтому патогенез вирусных инфекций, в первую очередь, следует рассматривать на молекулярном и клеточном уровнях (В.И. Покровский, О.И. Киселев, 2002). Вместе с тем, инфекционный процесс, вызванный вирусами, развивается в пределах того или иного органа или ткани, так как большинство вирусов обладают достаточно высокой органной или тканевой тропностью. Поэтому характер развития внутритканевых процес сов при вирусных инфекциях, с одной стороны, определяется, как правило, цитопатическим действием вируса на клетки данной ткани и органа, а с другой стороны, реакцией внутритканевых и органных систем защиты от вирусной инфекции. Последние не редко носят деструктивный характер и усугубляют течение забо левания и его последствия. Поэтому понимание патогенеза вирусных инфекций представляется исключительно важным для создания рациональных схем лечения, основанных на гармоничном сочетании симптоматического лечения и противовирусной химиотерапии.
Стадии репликативного цикла вируса гриппа представлены на рис. 1. Такой же репликативный цикл свойствен большинству РНК- и ДНК-содержащих вирусов, кроме тех, кото рые обладают особыми механизмами хронизации процесса, латенции и способностью к интеграции в клеточный геном (например, ВИЧ, гепатит С, вирус папилломы и.т.). Ключевыми стадиями репликативного цикла вируса являются проникновение в клетки, декапсидация, активация процессов транскрипции и трансляции, собственно репликация вирусного генома и созревание зрелых вирусных частиц с выходом нового потомства инфекционного вируса.
Проникновение вируса в клетки - одна из ключевых стадий патогенеза инфекционного процесса. Для большинства вирусов степень цитопатического действия вируса прямо зависит от множественности заражения. В естественных условиях инфицирова ние клеток вирусами не происходит обычно с высокой множе ственностью. Исключение составляют респираторные вирусы, которые передаются воздушно-капельным путем с высокой инфицирующей дозой. ВИЧ и гепатиты, наоборот, эффективно передаются с кровью в самых незначительных трудно определяе мых титрах. Такие вирусы имеют особые свойства, определяющие их контагиозность.
Для проникновения вируса в клетку, необходимо: высокое сродство к вирусоспецифическому рецептору; множественность рецепторов; эффективность слияния вируса с клетками при инфицировании; способность к образованию синцития, к образованию гигантских синпластов, состоящих из множества слившихся клеток, в которых вирусный нуклеопротеид без эндоцитоза беспрепятственно переходит от одной зараженной клетки к множеству других через цитоплазму.
Такими свойствами из респираторных вирусов обладает респираторно-синцитиальный вирус, а среди ретровирусов – вирус иммунодефицита человека.
Стадии жизненного цикла вируса гриппа и соответствующие ингибиторы репродукции в известной степени могут быть экстраполированы на многие другие вирусы, за исключением тех, жизненный цикл которых включает интеграцию в клеточный геном. В процессе репликации вирусов обычно происходит селективная блокада трансляции клеточных мРНК и активация экспресси клеточных генов, кодирующих провоспапительные лимфокины. Многократные раунды репликации вирусного генома приводят к истощению энер гетических ресурсов клеток и пула предшественников азотистых оснований. Повышенное содержание провоспалительных лимфокинов приводит к развитию воспалительной реакции и локально му генерированию свободных радикалов, оказывающих, в свою очередь, повреждающее действие на клетки и ткани через перекисное окисление липидов клеточных мембран, инактивацию жизненно важных ферментов. Поэтому одним из важнейших факторов патогенности вирусов является их репликативный потенциал: чем активнее происходит репликация, тем сильнее цитопатическое действие вирусов на клетки и пораженную ткань (орган).
Значительный вклад в цитопатическое действие вирусов вносит репродукция вирусов в митохондриях, что сопровождается нарушением функций этих органоидов, падением продукции АТФ и индукцией опосредованного митохондриями пути апоптоза.
Впервые возможность репродукции вирусов в митохондриях была доказана выдающимся отечественным вирусологом В.М. Ждановым. Узкий тропизм вирусов, в известной степени, спасителен для организма, так как инфекционный процесс ограничен одним органом.
Это обстоятельство позволяет, проводит рациональную симптоматическую терапию, направленную на защиту данной ткани или органа и ориентировать противовирусные средства в данный орган, например, легкие.
Возбудители гриппа относятся к семейству ортомиксовирусов (Orthomyxo-viridae) и являются пневмотропными вирусами. Схематическое строение вируса гриппа представлено на рис. 2.1. Геном вируса складывается из 8-ми фрагментов однонитчатой РНК, которые кодируют
10 вирусных белков. Фрагменты РНК имеют общую белковую оболочку, соединяющую их, образуя антигенно-стабильный рибонуклеопротеид (S-антиген), который определяет принадлежность вируса к серотипу А, В или С. Снаружи вирус покрыт двойным липидным слоем с внутренней стороны которого находится слой мембранного белка.
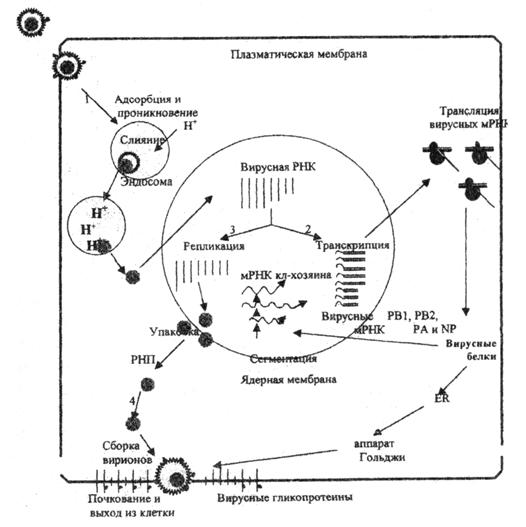
Рис. 1. Репликативный цикл вируса гриппа (В.И.Покровский, О.И.Киселев, 2002)
Представлены 4 основные фазы репликативного цикла вируса гриппа: 1 – адсорбция и проникновение вируса в клетку хозяина; 2 – транскрипция вирусной РНК и трансляция вирусных белков; 3 – репликация вирусной РНК и 4 – самосборка вирионов и последующий выход их из клетки.
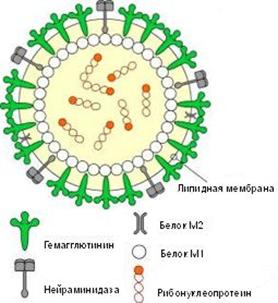
Рис. 2. Схематическое строение вируса гриппа
Гемаглютинин и нейраминидаза являются факторами агрессии вируса гриппа. Интенсивность интоксикации при гриппе определяется свойствами гемаглютинина, а нейраминидаза обладает выраженным иммунодепресивным действием. Оба поверхностных антигена характеризуются выраженной способностью к изменчивости, в результате чего появляются новые антигенные варианты вируса. Гемаглютинины 1, 2, 3 типов и нейраминидазы 1, 2 типов содержат вирусы, которые поражают человека. Другие антигены характерные для вирусов гриппа животных (свиней, собак, лошадей, многих видов птиц и др.).
Вирусы гриппа А. Эти возбудители более вирулентны и контагиозны, чем вирусы гриппа В и С. Это обусловлено тем, что вирус А содержит 2 типа нейраминидазы (N1, N2) и 4 –гемаглютинина (H0, H1, H2, H3); вирус В – 1 и 1 соответственно, поэтому он имеет лишь антигенные варианты внутри одного серотипа; вирус С содержит только гемаглютинин и не содержитнейраминидазу, но он имеет рецептор, который разрушает энзим. Вирусу С не свойственная изменчивость.
Одновременно циркулируют и имеют эпидемическое распространение 2 подтипа вируса гриппа А H3N2 и H1N1. На сегодняшний день во всем мире доминирует вариант А/Сидней/05/97, А/Берн/07/95 и А/Пекин/262/95. Характерной чертой современных вирусов гриппа А (H3N2) является изменение ряда их биологических свойств: тяжело размножаются в куриных эмбрионах, не агглютинируют эритроциты кур, взаимодействуют только с эритроцитами человека или морской свинки, имеют низкую иммуногенную активность.
Вирусы гриппа В выделяются только от людей. По вирулентности и эпидемиологической значимости они уступают вирусам гриппа А. По антигенным свойствам гемаглютинин инейраминидазу вируса В можно разделить на 5 подтипов. Их изменчивость носит более постепенный и медленный характер, что объясняет эпидемиологические особенности вируса гриппа В (эпидемии возникали 1 раз в 3-4 года до 1988 г.). В мире сейчас циркулируют 2 варианта вируса гриппа В: В/Пекин/184/93 (Европа, Америка, Африка, Австралия) и В/Виктория/2/87 (Юго-Восточная Азия). Характерной чертой современных вирусов гриппа А и В является их высокая чувствительность к ингибиторам гемаглютинирующей и инфекционной активности, которые содержатся в сыворотке крови нормальных животных.
Вирусы гриппа С . Вирусы гриппа С, в отличие от вирусов А и В, не вызывают эпидемий, но они могут быть причиной вспышек в организованных детских коллективах. Подъемы заболеваемости гриппом С нередко предшествуют или сопровождают эпидемии гриппа А и В. У детей раннего возраста и более старших возрастных группах он приводит к заболеваниям в легкой ибессимптомной форме.
Для вирусов гриппа С характерна значительно большая стабильность антигенных и биологических свойств. По ряду биологических характеристик эти возбудители отличаются от других представителей семейства ортомиксовирусов. Им присуща низкая репродуктивная активность в разных клеточных системах и наличие других, чем у вирусов гриппа А и В, рецепторов на поверхности эритроцитов. Рецептор-деструктирующая активность связана не с нейраминидазой, как у вирусов гриппа А и В, а с ферментом нейраминат-0-ацетилестеразою.
Сложность этиологии усиливают факты выделения от людей вирусов, которые не свойственные человеку. Гены вируса гриппа (H5N1), выделенные в 1997 году от человека в Гонконге, оказались подобными генам птичьего вируса, который преодолел межвидовой барьер. Оказалось, что человеческие и птичьи вирусы гриппа были реасортантами (рекомбинантные штаммы), которые получили внутренние гены РВ1 и РВ2 от перепелиного вируса гриппа А (H9N2) – штаммы А/перепел/Гонконг/61/97.
Вирусы гриппа малоустойчивы во внешней среде, в течение нескольких часов при комнатной температуре разрушаются. Под действием дезинфицирующих растворов (спирта, формалина, сулемы, кислот, щелочей) они быстро погибают. Нагревание до 50-60°С инактивирует вирусы в течение нескольких минут, в замороженном состоянии при температуре -70° С они сохраняются годами, не теряя инфекционных свойств, быстро погибают под воздействием ультрафиолетового излучения. Оптимум размножения вируса происходит при +37°С в слабощелочной среде. Как и другие вирусы, возбудитель гриппа отнюдь не чувствительный к антибиотикам и сульфаниламидам.
Вирус гриппа не растет на обычных питательных средах. Его можно выделить из материала, который получен от больного в первые дни болезни (смывы из носоглотки, мокроты), путем заражения культур или клеток куриных эмбрионов.
Традиционно считалось, что человеческий организм освобождается от возбудителя в ближайшие дни после выздоровления. Однако постепенно накапливались косвенные свидетельства, которые опровергают эту точку зрения. И в 1985 г. были получены прямые доказательства формирования длительной персистенции вируса гриппа. У одного человека удалось многократно выделять вирус гриппа в течение 9-ти месяцев и дольше. Возможность пожизненной персистенции вируса в организме человека продолжает изучаться.
1. Грипп и другие респираторные вирусные инфекции / под ред. О.И. Киселева, И.Г. Мариничева, А.А. Сомининой. – СПб. – 2003.
2. Дриневский В.П., Осидак Л.В., Цыбалова Л.М. Острые респираторные инфекции у детей и подростков // Практическое руководство под редакцией О.И. Киселева. – Санкт-Петербург. – 2003.
3. Железникова Г.Ф., Иванова В.В., Монахова Н.Е. Варианты иммунопатогенеза острых инфекций у детей. – Санкт-Петербург. – 2007. – 254 с.
4. Ершов Ф.И. Грипп и другие ОРВИ // Антивирусные препараты. Справочник. – М., 2006. –С. 226-247.
5. Ершов Ф.И., Романцов М.Г. Антивирусные средства в педиатрии. – М., 2005. – С.159-175.
6. Ершов Ф.И., Киселев О.И. Интерфероны и их индукторы (от молекул до лекарств). – М., 2005. – С. 287-292.
7. Иванова В.В. Острые респираторно-вирусные заболевания // Инфекционные болезни у детей. – М., 2002.
8. Онищенко Г.Г., Киселев О.И., Соминина А.А. Усиление надзора и контроля за гриппом как важнейший элемент подготовки к сезонным эпидемиям и очередной пандемии. – М., – 2004. – С.5-9.
9. Об утверждении стандарта медицинской помощи больным гриппом, вызванным идентифицированным вирусом гриппа (грипп птиц) // Приказ Минздравсоцразвития №460 от 07.06.2006 г.
10. Романцов М.Г., Ершов Ф.И. Часто болеющие дети: Современная фармакотерапия. – М., 2006. – 192 с.
11. Стандартизированные принципы диагностики, лечения и экстренной профилактики гриппа и других острых респираторных инфекций у детей / под ред. О.И. Киселева. – СПб. – 2004. – С. 82-95.
12. Лекарственные средства в фармакотерапии патологии клетки / под редакцией Т.Г. Кожока.-М., 2007.
Лекция 8
Патогенез вирусных инфекций
Модуль 3
Комплексная цель модуля
Комплексная цель модуля состоит в необходимости объединить лекционный материал, касающийся всех возможных проявлений вируса на клеточном и организменном уровне, продемонстрировать единую систему протекания вирусной инфекции при заражении организма-хозяина. В задачу лекционного материала, объединенного в данный модуль входит необходимость обобщить информацию о классификации вирусных инфекций, о вирусном патогенезе на клеточном и организменном уровне, о цитопатическом эффекте вирусного поражения, а также путях проникновения и распространения вирусов в организме хозяина.
Модуль состоит из двух лекций, материал которых позволяет решить поставленную цель.
Под инфекцией понимают комплекс процессов, происходящих при взаимодействии инфекционного агента с организмом хозяина. Однако в связи с тем, что вирусы являются внутриклеточными паразитами, а точнее, генетическими паразитами, в основе их взаимодействия с организмом всегда лежит инфекционный процесс на уровне клетки, который реализуется путем взаимодействия вирусного и клеточного геномов. Поэтому возможно классифицировать инфекции как на клеточном уровне, так и на уровне организма
КЛАССИФИКАЦИЯ ВИРУСНЫХ ИНФЕКЦИЙ НА КЛЕТОЧНОМ УРОВНЕ
Автономные и интеграционные инфекции. Если вирусный геном реплицируется независимо от клеточного генома, такая инфекция называется автономной. Понятие автономии относительно, оно ограничивается лишь отсутствием физической связи между вирусным и клеточным геномами, хотя взаимодействие их постоянно происходит в течение инфекции. Автономная форма вирусной инфекции характерна для большинства вирусов животных.
Если вирусный геном включается в состав клеточного генома, или, как принято называть этот процесс, интегрирует с клеточным геномом и реплицируется вместе с ним, такая инфекция называется интеграционной. Интеграционная инфекция возникает в результате физического объединения генома вируса и клетки. При этой форме инфекции вирусный геном реплицируется и функционирует как составная часть клеточного генома. Интегрировать могут как полный геном, так и часть генома. При гепатите В
возможна интеграция полного генома, при аденовирусных и герпесвирусных инфекциях обычно интегрирует часть генома, при инфекции онковирусами может интегрировать как полный геном, так и часть его. Вирусные последовательности в составе клеточного генома называются провирусом, или провирусной ДНК.
При интеграционных инфекциях нет ни сборки вирусной частицы, ни выхода вируса из клетки. Клетка может сохранить нормальные функции и при ее делении вирусные последовательности могут переходить в геном дочерних клеток. Такая ситуация наблюдается в случае инфекции, вызванной онкогенными вирусами. Интеграция может привести к неопластической трансформации клетки. Трансформированная клетка приобретает способность к неограниченному делению в результате нарушения регуляторных механизмов, контролирующих деление. Интеграционный тип инфекции возможен для нескольких семейств ДНК-содержащих вирусов: аденовирусов, паповавирусов, вирусов герпеса, а также для вируса гепатита В и обязателен для одного семейства РНК-содержащих вирусов — ретровирусов. В соответствии с данными В. М. Жданова, интеграционная форма инфекции может возникнуть при заражении и другими РНК-содержащими вирусами, такими, как вирус клещевого энцефалита (семейство тогавирусов), вирусы кори и SV5 (семейство парамиксовирусов) и др. Обязательным условием в этом случае является присутствие в клетках фермента — обратной транскриптазы, необходимого для процесса интеграции. Возникающая интеграционная инфекция может явиться причиной ряда хронических и аутоиммунных заболеваний.
Механизм интеграции вирусного генома с клеточным геномом. Из многих моделей, объясняющих процесс интеграции, наиболее признанной является модель Кемпбелла. В соответствии с этой моделью для интеграции с клеточным геномом необходима кольцевая форма двунитчатой вирусной ДНК. Эта молекула ДНК прикрепляется к клеточной ДНК, в месте прикрепления обе молекулы разрезаются и образовавшиеся концы сшиваются таким образом, что вирусная ДНК становится частью клеточного генома. Существенную роль в интеграции играют длинные концевые повторы двунитчатой ДНК, которые определяют специфичность интеграции в результате узнавания ими определенных участков клеточного генома. ДНК паповавирусов является циркулярной и двунитчатой и полностью отвечает требованиям модели Кемпбелла.
Продуктивная и абортивная инфекции. Инфекция может быть продуктивной и абортивной. Продуктивная инфекция завершается образованием инфекционного потомства. Абортивной называется инфекция, которая не завершается образованием инфекционных вирусных частиц, или они образуются в гораздо меньшем количестве, чем при продуктивной инфекции. Абортивная инфекция может возникнуть при следующих трех обстоятельствах: 1) заражение чувствительных клеток дефектным вирусом; 2) заражение чувствительных клеток в неразрешающих условиях; 3) заражение нечувствительных клеток стандартным вирусом.
Заражение чувствительных клеток дефектным вирусом. Дефектным называется такой вирус, который не способен проявить все генетические функции, необходимые для образования инфекционного потомства.
Существуют дефектные вирусы и дефектные вирусные частицы. Дефектными называются такие вирусы, которые репродуцируются лишь в присутствии вируса-помощника, например аденоассоциированный вирус (семейство парвовирусов), дающий потомство только в присутствии аденовируса-помощника. Дефектные вирусные частицы накапливаются в популяции многих вирусов, особенно при пассировании их с высокой множественностью инфекции. Дефектные частицы интерферируют при репродукции вируса с инфекционными вирусными частицами и потому называются дефектными интерферирующими частицами (ДИ-частицами). Этот тип вирусных частиц наиболее хорошо изучен на модели вирусов везикулярного стоматита и гриппа. Получение дефектных частиц вируса гриппа при заражении куриных эмбрионов с высокой множественностью инфекции получило название феномена фон Магнуса по имени исследователя, впервые его описавшего. Дефектные вирусные частицы вызывают абортивную инфекцию в связи с тем, что они лишены части генетического материала. Например, дефектные частицы вируса гриппа содержат неполные последовательности Р-генов, кодирующих три высокомолекулярных вирусных белка.
Заражение чувствительных клеток в неразрешающих условиях. Абортивная Инфекция может возникать при изменении условий, в которых происходит инфекционный процесс. Эти условия возникают в организме и могут моделироваться в эксперименте; в организме — повышение температуры, изменение рН в очаге воспаления и концентрации ионов, наличие антиметаболитов, ингибиторов и т. д.; в эксперименте — изменение температуры инкубации, состава питательной среды, внесение антиметаболитов и ингибиторов и т. д. В результате клетка либо погибнет без продукции инфекционного вируса, либо инфекция прерывается на определенном этапе. При устранении неразрешающих условий абортивная инфекция превращается в продуктивную. Смена абортивной инфекции на продуктивную может осуществиться и с помощью вируса-помощника.
Заражение нечувствительных клеток стандартным вирусом приводит к наиболее распространенной форме абортивной инфекции.
Непермиссивность клетки к определенному вирусному агенту может проявиться на любом этапе инфекции. Чувствительность клетки к ряду вирусов определяется наличием на клеточной поверхности специфических рецепторов, обусловливающих адсорбцию и проникновение вируса в клетку. Такой генетически обусловленный механизм клеточной резистентности наиболее четко установлен для пикорнавирусов, а также онковирусов птиц. Для большинства вирусов можно подобрать две клеточные системы, в одной из которых будет развиваться продуктивная, а в другой — абортивная инфекция. Механизм генетически обусловленной резистентности клеток к вирусам широко варьирует, но в основе его лежит либо отсутствие клеточных факторов, необходимых для репродукции вируса, либо наличие факторов, нарушающих процесс репродукции.
У сложно устроенных вирусов клеточная непермиссивность часто проявляется на стадии сборки вирусных частиц; нарушение сборки в некоторых непермиссивных системах для вирусов гриппа и парамиксовирусов обусловлено уменьшением количества молекул матриксного белка вируса.
Острая и хроническая инфекция.Как продуктивная, так и абортивная инфекция может протекать в виде острой или хронической инфекции.
Острой называется такая форма инфекции, при которой после образования вирусного потомства клетка либо погибает, либо выздоравливает и не содержит вирусных компонентов. Хроническая инфекция — это такая форма инфекции, при которой клетка продолжает продуцировать вирусные частицы или вирусные компоненты в течение длительного времени и передает эту способность дочерним клеткам.
Чаще хроническую форму приобретает абортивная инфекция, так как вирусный генетический материал обычно не входит в состав вирусного потомства, а накапливается в клетках и передается в дочерние клетки. Одним из факторов, вызывающих хроническую инфекцию, являются ДИ-частицы. Такие частицы, попадая в клетки вместе с инфекционными вирусными частицами, конкурируют с ними за факторы репродукции и препятствуют образованию инфекционного потомства. В результате гибель клеток предотвращается. При появлении в системе новых чувствительных клеток в них вновь возникает продуктивная инфекция с образованием ДИ-частиц, и такой цикл инфекции возобновляется снова и снова.
Цитолитическая и нецитолитическая инфекции. Острая инфекция на клеточном уровне может быть цитолитической и нецитолитической в зависимости от судьбы зараженной клетки. Инфекция, завершающаяся гибелью (лизисом) клетки называется цитолитической. Инфекция, которая непосредственно не приводит к лизису клетки, и клетка еще может функционировать в течение некоторого периода времени, продуцируя вирусные частицы, называется нецитолитической.
Смешанная инфекция. Вестественных условиях распространен феномен смешанной инфекции, при котором клетка заражается двумя или несколькими разными вирусами. Два и больше инфекционных процесса, происходящих одновременно в одной клетке, могут оказывать различное влияние друг на друга. Возможны несколько вариантов взаимодействия вирусов в процессе смешанной инфекции.
1. Один из вирусов подавляет репродукцию второго вируса, или подавляется репродукция обоих вирусов. Этот феномен называется интерференцией вирусов.
2. Вирус усиливает репродукцию второго вируса в результате комплементации или экзальтации. Комплементация может происходить между двумя родственными или неродственными вирусами, например между аденовирусом и аденоассоциированным вирусом человека или SV40, при этом вирус-помощник предоставляет другому вирусу неструктурный белок. Экзальтация может быть связана с подавлением процесса образования интерферона первым вирусом.
3. Оба вируса не оказывают существенного влияния на процесс репродукции каждого из них, однако может происходить нарушение морфогенеза обоих вирусов.
Смешанная инфекция широко используется вирусологами для изучения генетических функций вирусов и дефектности геномов.
ЦИТОПАТОЛОГИЯ ЗАРАЖЕННОЙ ВИРУСОМ КЛЕТКИ
Цитопатический эффект является следствием нескольких причин: 1) нарушение нормальной жизнедеятельности клетки в результате механического повреждающего действия вирусных компонентов на клеточные структуры; 2) повреждение лизосом, в результате чего освобождаются высокоактивные лизосомальные ферменты, вызывающие аутолиз клетки; 3) интенсивное истощение белковых и энергетических ресурсов клетки за счет переключения клеточных ферментов и белок-синтезирующего аппарата на синтез вирусспецифических макромолекул; 4) специфическое повреждающее действие вирусов на клеточные молекулы. Эти причины повреждения клетки различным образом проявляются и сочетаются при разных вирусных инфекциях.
Симпласты. Некоторые вирусы вызывают характерный цитопатический эффект, проявляющийся в слиянии клеток и образовании многоядерных клеток, называемых симпластами или синтицием. Образование симпластов обусловлено действием на клеточные мембраны прилежащих друг к другу клеток вирусных белков слияния и определяется тем же механизмом, который обеспечивает слияние вирусной и клеточной мембраны и проникновение вирусов в клетку. Слияние может происходить как за счет белков родительского вируса при заражении клеток большими концентрациями вируса (слияние снаружи), так и за счет внутриклеточного накопления вновь синтезированных вирусных белков слияния (слияние изнутри). Образование симпластов вызывают многие вирусы: парамиксовирусы, некоторые ретровирусы, вирусы герпеса. В определенных условиях (при низких значениях рН) слияние вызывают вирусы гриппа, буньявирусы и др.
Особенности вирусной инфекции в клеточной популяции. Основной особенностью вирусной инфекции в клеточной популяции является гетерогенность системы в связи с гетерогенностью вирусных частиц и клеток, входящих в состав популяции. В любом вирусном препарате наряду с инфекционными вирионами находятся ДИ-частицы. Клетки в каждой клеточной популяции широко варьируют по чувствительности к вирусу, и инфекция может протекать не так, как на клеточном уровне. Например, при заражении вирусом, вызывающим в клетках продуктивную инфекцию, чувствительные клетки популяции могут погибнуть, и в популяции за счет некоторого количества нечувствительных клеток может установиться хроническая инфекция.
Тропизм – склонность вируса к тому или иному вороту инфекции. При респираторных инфекциях – вирус локализуется в носоглотке, трахее и легких; при энтеровирусных – в кале; при нейротропных – в ГМ или СМ; при дермотропных – в коже.
Патогенез вирусных инфекций.
Под патогенезом понимают совокупность процессов, вызывающих заболевание, его развитие и исход.
2.Количеством инфекционных частиц
3.Реакцией клетки на инфекцию.
4.Реакция организма на изменение клеток и тканей.
В основе тропизма вирусов лежит чувствительность к вирусу определенных клеток.
Патогенез обусловлен основными механизмами взаимодействия вирусов с клетками:
- атрофия или дистрофия (ЦПД)
- образование телец включений
- образование симпластов и синцитиев
- латентная (хроническая) инфекция.
Под инфекцией понимают состояние зараженности макроорганизма. Это комплекс процессов, происходящих при взаимодействии инфекционного агента с организмом хозяина. В связи с тем, что вирусы являются внутриклеточными паразитами в основе их взаимодействия с организмом всегда лежит инфекционный процесс на уровне клетки, который реализуется путем взаимодействия вирусного и клеточного генома, поэтому возможно классифицировать инфекцию как на клеточном, так и на организменном уровне.
Патогенез на клеточном уровне – сюда входит ЦПД (видимые морфологические изменения клеток под воздействием того или иного вирусного агента). Характер ЦПД различен и зависит от:
2.Биохимических свойств вируса
Характер ЦПД оценивается по 4-х бальной системе крестовой и учитываются изменения, когда используются культуры клеток для титрования (т.е.).
Патогенез на организменном уровне.
Состояние инфекции как всякого биологического процесса динамично, динамку взаимодействия обычно называют инфекционным процессом. С одной стороны инфекционный процесс включает: внедрение, размножение и распространение возбудителя в организме, а также патогенное действие, а с другой стороны реакцию организма на это действие.
Патогенное действие возбудителя может быть неодинаковым. Оно проявляется в форме инфекционной болезни различной тяжести, в другом без ярко выраженных клинических признаков в третьих проявляется лишь изменениями, выявленными вирусологическими, биохимическими, иммунологическими методами. Это зависит от:
- количества и качества возбудителя, проникшего в восприимчивый организм, условий внутренней и внешней среды, определяющих резистентность животного и характеризуются взаимодействием микро и макроорганизмов. По характеру взаимодействия возбудителя болезни и организма выделяют 3 формы:
1.инфекционная болезнь – это инфекционный процесс, характеризуется определенными клиническими признаками, а также нарушениями, функциональными расстройствами и морфологическими повреждениями тканей.
2.Микробоносительство – иммунологическая субинфекция. Дифференцированный подход к различным формам инфекции дает возможность правильно вести диагностику инфекции выявлять зараженных животных в неблагополучном стаде. Патогенез любой инфекционной болезни определяется специальным действием возбудителя и ответными реакциями организма, зависящими от условий, в которых происходит взаимодействие микро и макроорганизма. При этом немаловажное значение имеют пути проникновения и распределения возбудителя. Ворота возбудителя: кожа, слизистые, мочеполовая система, плацента.
Каждый вид возбудителя эволюционно приспособился к таким путям внедрения, которое обеспечивает благоприятные условия для размножения и распространения – входные ворота для каждой инфекции характеризуется специфичностью. Чтобы проводить профилактику необходимо учитывать специфичность ворот инфекции. Например, при ИНАН возбудитель проникает через кожу при укусе насекомых. При ящуре основной путь алиментарный, при бешенстве – через покус.
Классификация вирусных инфекций.
Различают автономные и интегрированные инфекции. Автономные - при этом вирусный геном реплицируется независимо от клеточного генома. Автономная инфекция характерна для большинства вирусов.
Интегрированные инфекции – вирусный геном включается в состав клеточного генома, т.е. интегрируются в клеточный геном и реплицируются вместе с ним. При этом вирусный геном реплицируется и функционирует как составная часть клеточного генома. Интегрировать может как полный геном так и часть. При интегрированных инфекциях нет ни сборки вирусных частиц ни выхода.
Автономная инфекция – клетка иногда приобретает способность к неограниченному делению в результате нарушения регулирующих механизмов, контролирующих деление. Это чаще наблюдается при онкогенных инфекциях.
Продуктивная и абортивная инфекции:
1.Продуктивная – завершается выходом инфекционного потомства.
2.Абортивная - инфекционного потомства не образуется или его мало.
Формы течения – как и продуктивная, так и абортивная могут протекать в острой и хронической форме. Острая инфекция – это инфекция, в результате которой клетка либо выздоравливает либо погибает. Острая инфекция на клеточном уровне может быть цитолитической (когда происходит гибель клетки).
Хроническая инфекция – это инфекция, при которой клетка продолжает продуцировать вирусные частицы в течение длительного времени и предает эту способность дочерним клеткам. Чаще хроническую форму приобретает абортивная инфекция т.к. вирусный материал накапливается и передается дочерней клетке.
Смешенная инфекция – клетка заражается двумя или несколькими разными вирусами, в результате чего в клетке могут совмещаться два и более инфекционных процесса. Возможно несколько вариантов взаимодействия вируса в процессе смешанной инфекции:
1.Интерференция – один вирус подавляет действие другого.
2.Комплементация (экзальтация) – один вирус усиляет действие другого.
Классификация вирусных инфекций на организменном уровне.
В основу классификации положено:
3.Проявление клинических симптомов
4.Выделение вирусов в окружающую среду
Одна из форм может переходить в другую (например, очаговая в генерализованную, острая в хроническую).
Вирус действует вблизи входных ворот инфекции, в связи с локальной репродукцией. Они имеют более короткий скрытый период по сравнению с генерализованными.
После ограниченного периода репродукции в первичных очагах происходит генерализация инфекций – вирусы проникают в другие системы, например при ящуре, полиомиелите, оспе.
Длится непродолжительный период и протекает с выделением в окружающую среду. Заканчивается гибелью или выздоровлением.
При продолжительном взаимодействии вируса с организмом. Она может быть латентная, хроническая, медленная.
Латентная инфекция – не сопровождается выделением вируса в окружающую среду, при определенных условиях может переходить в острую и хроническую.
При гриппе, сепсисе, СПИДе и др.
Это длительно текущий процесс. Характеризуется периодами ремиссии (аденовирус, герпес).
Медленные инфекции – своеобразное взаимодействие вируса с фагом и характеризуется длительными инкубационными периодами.
При изучении любого инфекционного заболевания важно знать источник, место постоянного обитания и размножения, пути распространения, место и сроки сохранения, возникновения во внешней среде, способы передачи от больных к здоровым.
Естественная среда – живой организм, здесь он находит все условия существования. Длительность пребывания вирусов колеблется в значительных пределах и зависит от биологических свойств, реактивности организма. От условий патогенеза. Источники инфекции – только зараженные организмы. Они играют роль лишь в процессе передачи. Большинство животных выделяют вирусы с экскретами, секретами, кровью, истечениями, мокротой. При большинстве вирусных инфекций в основе патогенеза лежит вирусемия (ящур, чума и др). При этих заболеваниях вирус выделяется всеми возможными путями. При хроническом течении вирусовыделение менее интенсивно, но может быть длительным. При вирусных заболеваниях локализация ограничивается одним путем: пневмонии – с каплями мокроты. Самое интенсивное выделение вируса во внешнюю среду наблюдается в острый период заболевания, однако при ряде заболеваний и в инкубационный период. Бессимптомные инфекции протекают при вакцинировании живыми вакцинами.

Организация стока поверхностных вод: Наибольшее количество влаги на земном шаре испаряется с поверхности морей и океанов (88‰).

Общие условия выбора системы дренажа: Система дренажа выбирается в зависимости от характера защищаемого.

Опора деревянной одностоечной и способы укрепление угловых опор: Опоры ВЛ - конструкции, предназначенные для поддерживания проводов на необходимой высоте над землей, водой.

Папиллярные узоры пальцев рук - маркер спортивных способностей: дерматоглифические признаки формируются на 3-5 месяце беременности, не изменяются в течение жизни.
Читайте также: