
Клетки с вирусными изменениями что такое
86. 1. Определение, химический состав вирусов, функции компонентов.
Функция нуклеиновой кислоты вируса: …
Функция белков вирусных частиц: …
Размножение вирусов, синтез их нуклеиновых кислот.
Где размножается вирус?
О видах вирусов
86. 2. (Молекулярные механизмы действия вирусов.)
Реакция иммунной системы на обнаружение вируса.
Как вирусная частица попадает в клетки.
Что происходит с вирусной частицей после её попадания в клетку.
86. 3. Онкогенные вирусы. См. п.87.
86. 4. ВИЧ. См. п.77 и 80.
86. 5. Прионы.
86. 1. Определение, химический состав вирусов, функции компонентов. –
Опеределение вируса.
Вирусами называют КОМПЛЕКСЫ НУКЛЕИНОВЫХ КИСЛОТ С БЕЛКАМИ,
которые могут находиться во внешней среде вне клеток и
могут при попадании в организм проникать в клетки и размножаться внутри клеток.
Некоторые вирусы вдобавок к НК и белкам содержат липиды и углеводы.
Таким образом, вирусы не являются клетками, у вирусов нет органелл.
Отдельный комплекс нуклеиновых кислот с белками называется вирусной частицей или вирионом.
Функция нуклеиновой кислоты вируса:
нуклеиновые кислоты используются для синтеза белков вирусных частиц (кодируют первичную структуру белка).
Функция белков вирусных частиц:
1 – защищают НК от повреждений (НК сами по себе очень неустойчивы), образуя оболочку вокруг НК,
2 – участвуют в размножении вирусов в качестве ферментов и т.д. см. далее.
Углеводы вирусных частиц часто находятся на поверхности вирусных частиц и могут помогать вирусным частицам проникать в клетки.
Отдельная вирусная частица и вирусные частицы данного вида вирусов содержат только один вид НК – или ДНК, или РНК.
Вирусы, в вирионе которых находится ДНК, называются ДНК-овые вирусами,
а вирусы, в вирионе которых находится РНК, называются, РНК-овыми вирусами.
ДНК вируса может быть не только двухцепочечной, но и ОДНОЦЕПОЧЕЧНОЙ.
РНК вируса может быть как одноцепочечной (как в клетках), так и ДВУХЦЕПОЧЕЧНОЙ.
Размножение вирусов, синтез их нуклеиновых кислот.
Синтез нуклеиновых кислот вирусов происходит разными способами. –
1) Наряду с репликацией ДНК (см. п. 78),
2) у вирусов есть репликация РНК
3) и обратная транскрипция (см. п.80).
Какой бы ни была нуклеиновая кислота в самом вирионе,
при размножении вируса происходит синтез вирусных матричных РНК для синтеза вирусных белков (для трансляции).
Где размножается вирус?
Синтез вирусных нуклеиновых кислот и синтез вирусных белков может происходить только внутри клеток,
так как для синтезов нужны среда, рибосомы (осуществляются синтез белка), нуклеотиды и аминокислоты и т.д.
Поэтому размножаться вне клеток вирусы не могут. –
Пока вирус не проник в клетку, он не может размножаться, не может как-то влиять на обмен веществ в организме.
Поэтому наличие вируса (вирионов) в крови
не означает, что человек будет болеть болезнью, которая вызывается этим вирусом.
Он может заболеть, только если вирусные частицы проникнут в клетки,
а если они не проникнут в клетки, то человек не заболеет, хоть и будет носителем вируса.
О видах вирусов
Вирусы, содержащие разные нуклеиновые кислоты (то есть с разными последовательностями нуклеотидов) и разные белки, относят к разным видам вирусов.
Вирусные частицы с одинаковыми (или очень похожими) последовательностями нуклеотидов в НК, относятся к вирусу одного и того же вида.
Кроме того, последовательность нуклеотидов нуклеиной кислоты вируса данного вида
может меняться, то есть вирус может мутировать.
В итоге получается другой ШТАММ данного вида вируса, но сам вирус относится к тому же виду вирусов, что и до мутации.
Вирусы, которые поражают бактерии, называются БАКТЕРИОФАГАМИ.
Бактериофаги могут использоваться для внесения в клетку бактерии чужих для неё генов для синтеза клеткой белков, кодируемых этими генами,
То есть бактериофаги могут играть роль векторов – это используется генной инженерией.
В эукариотические клетки вирусы попадают не так, как в бактериальные (далее).
86. 2. (Молекулярные механизмы действия вирусов.)
Реакция иммунной системы на обнаружение вируса.
При попадании вирусной частицы в организм её должны заметить и уничтожить клетки иммунной системы
(с помощью АНТИТЕЛ – специальных белков, задача которых – распознавание антигенов)
для того чтобы вирус не успел проникнуть в клетку, размножиться в ней, убить её и заражать новые клетки.
Как вирусная частица попадает в клетки.
Проникновение вирусной частицы в клетку начинается с того, что
вирус связывается с определённым белком на поверхности клетки,
который таким образом выполняет функцию РЕЦЕПТОРА клетки для этого вируса.
Роль рецептора вируса обычно играют белки, предназначенные для других целей –
например, ВИЧ использует рецепторы хемокинов.
Если рецепторы вируса отличаются от обычных так, что вирус не может с их помощью проникнуть в клетку, то вирус не приведёт к заболеванию.
Причиной отличия рецептора может быть мутация гена, который кодирует этот рецептор.
Что происходит с вирусной частицей после её попадания в клетку.
После попадания вирусной частицы внутрь клетки
вирусная нуклеиновая кислота высвобождается, после чего
могут синтезироваться новые молекулы вирусной нуклеиновой кислоты (см. п.77)
и новые молекулы вирусных белков (п.82).
После этого из молекул вирусных нуклеиновых кислот и молекул вирусных белков
могут образовываться новые вирусные частицы –
так происходит РАЗМНОЖЕНИЕ ВИРУСОВ.
Новые вирусные частицы могут покидать клетку, в которой образовались,
и поступать в новые клетки, заражая всё новые и новые клетки.
Если заражённые вирусом клетки подвергнутся АПОПТОЗУ (см. п. 79),
то распространение вирусных частиц может затормозиться.
Поэтому замедленный апоптоз способствует распространению вирусной инфекции.
Накопление вирусных частиц в клетке может приводить к РАЗРУШЕНИЮ КЛЕТКИ;
кроме того, оно нарушает нормальную жизнедеятельность клетки и организма –
всё это приводит к появлению симптомов вирусной инфекции.
Нуклеиновые кислоты некоторых вирусов (см. ВИЧ далее) могут использоваться
в качестве матриц для образования двухцепочечных ДНК (вирусных ДНК),
которые способны встраиваться в ДНК человека –
это встраивание называется интегрированием в геном,
оно относится к ПЕРЕСТРОЙКАМ генома – см. п.79.
86. 3. Онкогенные вирусы. См. п.87.
Некоторые вирусы способствуют появлению онкологических заболеваний.
Такие вирусы называются онкогенными вирусами.
Примеры онкогенных вирусов:
вирус папилломы человека способен приводит к раку матки,
вирусы гепатитов В и С приводят к раку печени.
Онкогенные вирусы относят к группе биологических мутагенов – см. п.79.
Почему некоторые вирусы могут приводить к онкологическим заболеваниям (см. п.87 и п.92, 98):
потому что онкогенные вирусы приводят к таким МУТАЦИЯМ,
которые способствуют появлению онкологических заболеваний.
Например, считается, что онкогенными являются те вирусы,
которые приводят к встраиванию вирусных ДНК в ДНК (геном) человека (интегрируется).
Потому что это встраивание может изменить тот участок ДНК,
в который встраивается вирусная ДНК (то есть привести к мутации).
В норме, до мутации, протоонкоген кодирует белки, которые стимулируют деление клеток и относятся к онкобелкам.
В норме активность онкобелков регулируется,
поэтому они стимулируют деление клетки только тогда, когда нужно –
и поэтому деление клетки (и образованных при её делении клеток) не приводит к образование опухоли.
Но после мутации протоонкогена он может измениться так, что
кодируемые им онкобелки не смогут регулироваться
и станут стимулировать деление клетки постоянно, без остановки,
что может привести к появлению опухоли,
если мутантные клетки не подвергнутся апоптозу
или не уничтожатся клетками иммунной системы.
86. 4. ВИЧ. См. п.77 и 80.
ВИЧ – это вирус иммунодефицита человека.
Размножение ВИЧ в организме человека может привести к развитию СПИДа –
Синдрома Приобретённого Иммунодефицита Человека.
ВИЧ повреждает клетки иммунной системы (Т-хелперы, макрофаги),
что приводит к дефициту иммунных клеток и снижению иммунитета (иммунодефициту).
1. После попадания ВИЧ в организм человека
ВИЧ, как и все вирусы, сначала связывается с рецепторами на поверхности клеток:
ВИЧ связывается с рецепторами хемокинов
(хемокиновыми рецепторами;
хемокины – это гормоны, относятся к цитокинам)
и СД4 рецепторами на поверхности клеток иммунной системы.
Связыванию ВИЧ с рецепторами помогают гликопротеины, расположенные на поверхности вирусной частицы ВИЧ.
Есть люди, в организме которых есть ВИЧ, но у которых не развивается СПИД –
то есть они являются носителями вируса, они могут заразить ВИЧ,
но больными они не являются.
Предполагают, что это связано с тем, что ВИЧ не может размножаться в организме этих людей,
поскольку не проникает в их клетки из-за особенностей строения рецепторов ВИЧ у этих людей:
возможно, из-за мутаций генов, кодирующих эти рецепторы.
2. В качестве нуклеиновой кислоты в ВИЧ содержится одноцепочечная РНК.
В клетках РНК вируса становится матрицей для синтеза комплементарной ей цепи ДНК
(комплементарной ДНК, кДНК),
который катализируется ферментом обратная транскриптаза
(потому что синтез ДНК на матрице РНК называется обратной транскрипцией),
который содержался в вирусной частице ВИЧ.
Затем на матрице первой цепи ДНК синтезируется вторая цепь ДНК,
в результате чего образуется двухцепочечная ДНК.
Эта вирусная ДНК способна встраиваться в ДНК человека
с помощью фермента ИНТЕГРАЗЫ, который тоже содержится в вирусной частице ВИЧ.
3. В некоторый момент вирусная ДНК, находящаяся в составе ДНК человека,
может использоваться для синтеза вирусных РНК (то есть для транскрипции – п.80).
4. Вирусные РНК могут использоваться для синтеза вирусных белков (для трансляции) с помощью рибосом самой клетки.
5. Один из ферментов ВИЧ осуществляет расщепление полипептидной цепи, синтезированной на вирусной РНК;
этот фермент называется ПРОТЕАЗОЙ.
В итоге образуются вирусные белки, в том числе уже названные протеаза, интеграза и обратная транскриптаза.
6. Объединение вирусных белков с вирусной РНК приводит
к появлению новых вирусных частиц ВИЧ,
которые могут разрушать клетки, поступать в кровь и проникать в новые клетки.
Диагностика ВИЧ
осуществляется методом ПЦР и с помощью иммуноферментного анализа.
Метод ПЦР быстрее и надёжнее.
СПИД пока считается неизлечимым заболеванием.
Но замедлить развитие СПИДа и продлить жизнь человека, инфицированного ВИЧ, можно за счёт замедления размножения ВИЧ в организме заражённого.
Размножение ВИЧ замедляется за счёт применения лекарств,
снижающих активность ферментов ВИЧ, участвующих в размножении ВИЧ (см. выше),
то есть за счёт применения ИНГИБИТОРОВ ферментов ВИЧ.
Чем раньше начинают лечение инфицированного ВИЧ – тем дольше он проживёт. Поэтому важно вовремя делать анализ на ВИЧ при наличии оснований предполагать, что человек мог заразиться ВИЧ.
При наличии ВИЧ у беременной есть шанс родить здорового ребёнка,
если вовремя сделать анализ на ВИЧ и провести лечение.
Прионами называют белки с особой структурой, которая приводит к так называемым прионовым болезням:
куру, болезни Крейцфельдта-Якобы, почесухе овец, коровьему бешенству.
У прионов есть способность
превращать молекулы нормальных белков в прионы
за счёт того, что под влиянием структуры уже имеющихся молекул прионов
структура молекул ряда других белов может измениться так,
что эти молекулы тоже становятся прионами –
нерастворимыми в воде молекулами,
под влиянием которых могут стать прионами другие молекулы белков –
то есть происходит как бы цепная реакция;
из-за этого число прионов в организме становится всё больше.
Из-за этого свойства прионы считаются инфекционными белками,
а прионовые болезни считаются заразными
(заражение происходит при попадании прионов в организм – обычно в качестве пищи).
В случае с прионами увеличение числа болезнетворных молекул
происходит не путём деления клеток
(в отличие от бактериальных и других инфекций, где бактерии размножаются делением)
и не путём синтеза нуклеиновых кислот (см. выше о размножении вирусов),
а под влиянием уже имеющихся молекул прионов.
Прионы могут оказаться в организме не только в результате заражения
(не только при съедении пищи, содержащей прионы),
но и в результате мутаций генов некоторых белков.
Из-за своей структуры прионы не уничтожаются и не теряют свои вредные свойства при сколь угодно долгой варке, поэтому варка не является способом сделать пищу безопасной и свободной от прионов. Единственный способ исключить возможность заражения прионами при эпидемиях коровьего бешенства – это не употреблять в пищу мясо, тушёнку, консервы, желатин и т.д.А единственный способ уничтожить прионы - только сжечь. Прионы устойчивы и к действию других факторов, которые разрушают структуру обычных белков, то есть прионы устойчивы к денатурации под действием излучений, ультразвука и т.д.
Вызываемое вирусами развитие клинических проявлений болезни и видимые признаки воспаления есть конечный результат множества событий. Большая часть процессов, связанных с репликацией вируса, а следовательно, с начальными воспалительными реакциями на микроструктурном уровне, недоступна для распознавания на клиническом уровне. Эта особенность, характерная для многих вирусных инфекций и впервые описанная С.А. Mims и А.Н. Sharpe], представляет собой важную концепцию для понимания вирусного воспаления. Зачастую привычные признаки: краснота, припухлость, инфильтрация составляют лишь видимую с большим опозданием и весьма незначительную часть того воспаления, которое захватывает многочисленные структуры пораженного организма.
Основные этапы взаимодействия вирусов и клеток чрезвычайно кратко представлены в модифицированной таблице.
В правом столбце таблицы перечислены события, происходящие внутри клетки, именно они определяют ее конечную судьбу: гибель, трансформацию, резервирование персистентного вируса и т.д. В левом столбце приведены события на уровне организма или ткани, ведущие к заражению клеток и развитию защитных иммунных реакций. Эта схема значительно упрощена, многое зависит от специфики вируса и хозяина.
Респираторные вирусы чаще проникают в организм через верхние дыхательные пути. Первичное прикрепление вируса к эпителиальным клеткам зависит от множества факторов: его устойчивости к неспецифическим защитным факторам (протеолитические ферменты, слизь, движение ресничек, соли желчных кислот), к секреторным антителам, а также от наличия специфических комплементарных рецепторов на поверхности клетки, способных к воссоединению с рецепторами вируса.
Как показали исследования последних лет, комплементарность вирусных и клеточных рецепторов зачастую обусловлена генетическими маркерами организма: группой крови и HLA-принадлежностью. Так, самые тяжелые осложненные формы гриппа чаще возникают у носителей антигенов А-Н и HLA-8, склонность к персистированию превалирует у лиц с группой крови ВШ и HLA-A9. Таким образом, развитие тех или иных форм воспаления отчасти предопределено генетически. Не исключено, что каждому из респираторных вирусов соответствует свой излюбленный генотип.
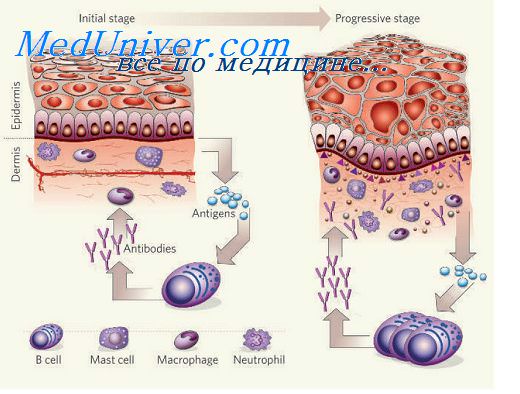
Вторым, не зависящим от специфических рецепторов, механизмом органотропности является протеолитическое расщепление гликопротеиновых оболочек парамиксовирусов (парагрипп, PC-вирус) в присутствии трипсиноподобных ферментов, содержащихся в инфицируемых клетках. Аналогичное свойство присуще вирусу гриппа, у которого ферментативному расщеплению подвергается гемагглютинин. При отсутствии в ткани необходимых ферментов многоцикловая инфекция не воспроизводится.
Прикрепление вируса к клетке является пусковым моментом для развития симптомокомплекса воспаления, который включает в себя альтерацию, экссудацию, трансформацию и пролиферацию в сочетании с нарушениями микроциркуляции и повреждениями функциональной активности. Краткая схема этого сложного и многообразного процесса не может отразить всех возможных вариантов и особенностей, но может облегчить восприятие основных моментов. Большая часть перечисленных в таблице 2 признаков воспаления, прежде всего деструкция и дистрофия, являются общими для всех респираторных вирусов и не могут служить надежными признаками дифференциации. Однако характер вирусных включений: их размеры, локализация, структура в сочетании с определенным типом поражения клеток (образование гигантских структур, пластов, скопления гроздевидных округлых клеток и т.д.) имеют определенное диагностическое значение при рассмотрении воспаления на субклеточном и клеточном уровне. Так, уже в начальной стадии воспаления, через 6-8 часов от момента заражения, при гриппе определяются базофильные включения, содержащие вирусные частицы. Чаще всего они локализуются в апикальной части эпителия; эозинофильные включения реже содержат вирус и чаще рассматриваются в качестве реактивных образований. При парагриппе и PC-инфекции выявляются цитоплазматические фуксинофильные включения, нередко группирующиеся вокруг ядра. При герпетической и цитомегаловирусной инфекции чаще регистрируются четко контурируемые внутриядерные включения, округлые, крупные, эозинофильные и оксифильные, нередко отгороженные от остальной цуклеплазмы светлой зоной. При аденовирусной инфекции также преобладают внутриядерные включения, на ранних стадиях инфекции (через 16 часов) - мелкие, округлые, базофильные или оксифильные; одни из них однородны, другие имеют форму колец с прозрачным центром. В дальнейшем число внутриядерных включений увеличивается, встречаются слоистые и ячеистые формы. На поздних этапах инфекции включения обнаруживаются и в цитоплазме клеток. Наиболее точная дифференцировка специфичности включений осуществляется с помощью прямого и непрямого иммунофлуоресцентного метода с использованием меченых антител.
Формирование вирусных включений интересно не только с точки зрения патогенных возможностей респираторных вирусов, но и с точки зрения анализа самого понятия воспаления. С одной стороны, это несомненный результат непосредственного воздействия вируса на клетку, вероятно это защитная реакция клетки, стремящейся локализовать патогенные частицы, а с другой стороны - это и возможность длительной резервации вируса. Таких двояких по целесообразности моментов в процессе вирусного воспаления достаточно. Так, под действием вирусов может происходить своеобразная активация фагоцитарной способности клеток, что проявляется в усилении цитофагоцитоза, когда инфицированные клетки активно фагоцитируют другие клетки, а также ферменты распавшихся структур. С одной стороны, это полезно с точки зрения утилизации и удаления дегенерировавших элементов, с другой стороны - нередко способствует большему распространению вирусной инфекции по клеточному пласту.
Большинство респираторных вирусов на ранних стадиях инфицирования вызывает ряд изменений клеточных структур, которые можно рассматривать как защитно-компенсаторные: набухание, гиперплазия ряда клеточных элементов, увеличение функционирующих локусов митохондрий, и лишь на более поздних стадиях возникают явления дистрофии, деструкции, либо необратимого перерождения.
Перечисляя изменения клеточных элементов, обусловленных вирусной репродукцией, было бы несправедливым забыть о главной их способности - нарушать синтез клеточных макромолекул. В случае развития вирусного инфицирования в клетке появляется два источника информации: клеточный и вирусный геномы. В подавляющем большинстве случаев ведущим оказывается вирусный геном, под руководством которого осуществляются процессы, направленные на воспроизводство вирусного потомства, в то время как синтез клеточных макромолекул (РНК, ДНК, белка) претерпевает значительное подавление. После длительных экспериментальных поисков было доказано, что в основе ингибирования синтеза клеточных РНК и ДНК в инфицированных клетках лежит феномен блокирования процесса транскрипции, т.е. "считывания" информации, закодированной в клеточном геноме, без нарушения целостности структур ДНК и РНК, а также РНК-полимераз. Ингибитором, ответственным за этот процесс, оказался аргининсодержащий белок - гистон. Большая часть исследователей полагает, что синтез белков-ингибиторов осуществляется самой клеткой в результате реализации информации, закодированной в ее геноме. Вирус лишь стимулирует синтез этих ингибиторов. Помимо подавления синтеза клеточных РНК, гистоны подавляют функцию и синтез ДНК, а также синтез АТФ и перенос аминокислот. Таким образом, появление индуцированных вирусом и воспроизведенных клеткой гистонов является одной из основных причин нарушения синтеза клеточных структур, а, следовательно, и дистрофии, являющейся одним из признаков воспаления.
Лекция 8
Патогенез вирусных инфекций
Модуль 3
Комплексная цель модуля
Комплексная цель модуля состоит в необходимости объединить лекционный материал, касающийся всех возможных проявлений вируса на клеточном и организменном уровне, продемонстрировать единую систему протекания вирусной инфекции при заражении организма-хозяина. В задачу лекционного материала, объединенного в данный модуль входит необходимость обобщить информацию о классификации вирусных инфекций, о вирусном патогенезе на клеточном и организменном уровне, о цитопатическом эффекте вирусного поражения, а также путях проникновения и распространения вирусов в организме хозяина.
Модуль состоит из двух лекций, материал которых позволяет решить поставленную цель.
Под инфекцией понимают комплекс процессов, происходящих при взаимодействии инфекционного агента с организмом хозяина. Однако в связи с тем, что вирусы являются внутриклеточными паразитами, а точнее, генетическими паразитами, в основе их взаимодействия с организмом всегда лежит инфекционный процесс на уровне клетки, который реализуется путем взаимодействия вирусного и клеточного геномов. Поэтому возможно классифицировать инфекции как на клеточном уровне, так и на уровне организма
КЛАССИФИКАЦИЯ ВИРУСНЫХ ИНФЕКЦИЙ НА КЛЕТОЧНОМ УРОВНЕ
Автономные и интеграционные инфекции. Если вирусный геном реплицируется независимо от клеточного генома, такая инфекция называется автономной. Понятие автономии относительно, оно ограничивается лишь отсутствием физической связи между вирусным и клеточным геномами, хотя взаимодействие их постоянно происходит в течение инфекции. Автономная форма вирусной инфекции характерна для большинства вирусов животных.
Если вирусный геном включается в состав клеточного генома, или, как принято называть этот процесс, интегрирует с клеточным геномом и реплицируется вместе с ним, такая инфекция называется интеграционной. Интеграционная инфекция возникает в результате физического объединения генома вируса и клетки. При этой форме инфекции вирусный геном реплицируется и функционирует как составная часть клеточного генома. Интегрировать могут как полный геном, так и часть генома. При гепатите В
возможна интеграция полного генома, при аденовирусных и герпесвирусных инфекциях обычно интегрирует часть генома, при инфекции онковирусами может интегрировать как полный геном, так и часть его. Вирусные последовательности в составе клеточного генома называются провирусом, или провирусной ДНК.
При интеграционных инфекциях нет ни сборки вирусной частицы, ни выхода вируса из клетки. Клетка может сохранить нормальные функции и при ее делении вирусные последовательности могут переходить в геном дочерних клеток. Такая ситуация наблюдается в случае инфекции, вызванной онкогенными вирусами. Интеграция может привести к неопластической трансформации клетки. Трансформированная клетка приобретает способность к неограниченному делению в результате нарушения регуляторных механизмов, контролирующих деление. Интеграционный тип инфекции возможен для нескольких семейств ДНК-содержащих вирусов: аденовирусов, паповавирусов, вирусов герпеса, а также для вируса гепатита В и обязателен для одного семейства РНК-содержащих вирусов — ретровирусов. В соответствии с данными В. М. Жданова, интеграционная форма инфекции может возникнуть при заражении и другими РНК-содержащими вирусами, такими, как вирус клещевого энцефалита (семейство тогавирусов), вирусы кори и SV5 (семейство парамиксовирусов) и др. Обязательным условием в этом случае является присутствие в клетках фермента — обратной транскриптазы, необходимого для процесса интеграции. Возникающая интеграционная инфекция может явиться причиной ряда хронических и аутоиммунных заболеваний.
Механизм интеграции вирусного генома с клеточным геномом. Из многих моделей, объясняющих процесс интеграции, наиболее признанной является модель Кемпбелла. В соответствии с этой моделью для интеграции с клеточным геномом необходима кольцевая форма двунитчатой вирусной ДНК. Эта молекула ДНК прикрепляется к клеточной ДНК, в месте прикрепления обе молекулы разрезаются и образовавшиеся концы сшиваются таким образом, что вирусная ДНК становится частью клеточного генома. Существенную роль в интеграции играют длинные концевые повторы двунитчатой ДНК, которые определяют специфичность интеграции в результате узнавания ими определенных участков клеточного генома. ДНК паповавирусов является циркулярной и двунитчатой и полностью отвечает требованиям модели Кемпбелла.
Продуктивная и абортивная инфекции. Инфекция может быть продуктивной и абортивной. Продуктивная инфекция завершается образованием инфекционного потомства. Абортивной называется инфекция, которая не завершается образованием инфекционных вирусных частиц, или они образуются в гораздо меньшем количестве, чем при продуктивной инфекции. Абортивная инфекция может возникнуть при следующих трех обстоятельствах: 1) заражение чувствительных клеток дефектным вирусом; 2) заражение чувствительных клеток в неразрешающих условиях; 3) заражение нечувствительных клеток стандартным вирусом.
Заражение чувствительных клеток дефектным вирусом. Дефектным называется такой вирус, который не способен проявить все генетические функции, необходимые для образования инфекционного потомства.
Существуют дефектные вирусы и дефектные вирусные частицы. Дефектными называются такие вирусы, которые репродуцируются лишь в присутствии вируса-помощника, например аденоассоциированный вирус (семейство парвовирусов), дающий потомство только в присутствии аденовируса-помощника. Дефектные вирусные частицы накапливаются в популяции многих вирусов, особенно при пассировании их с высокой множественностью инфекции. Дефектные частицы интерферируют при репродукции вируса с инфекционными вирусными частицами и потому называются дефектными интерферирующими частицами (ДИ-частицами). Этот тип вирусных частиц наиболее хорошо изучен на модели вирусов везикулярного стоматита и гриппа. Получение дефектных частиц вируса гриппа при заражении куриных эмбрионов с высокой множественностью инфекции получило название феномена фон Магнуса по имени исследователя, впервые его описавшего. Дефектные вирусные частицы вызывают абортивную инфекцию в связи с тем, что они лишены части генетического материала. Например, дефектные частицы вируса гриппа содержат неполные последовательности Р-генов, кодирующих три высокомолекулярных вирусных белка.
Заражение чувствительных клеток в неразрешающих условиях. Абортивная Инфекция может возникать при изменении условий, в которых происходит инфекционный процесс. Эти условия возникают в организме и могут моделироваться в эксперименте; в организме — повышение температуры, изменение рН в очаге воспаления и концентрации ионов, наличие антиметаболитов, ингибиторов и т. д.; в эксперименте — изменение температуры инкубации, состава питательной среды, внесение антиметаболитов и ингибиторов и т. д. В результате клетка либо погибнет без продукции инфекционного вируса, либо инфекция прерывается на определенном этапе. При устранении неразрешающих условий абортивная инфекция превращается в продуктивную. Смена абортивной инфекции на продуктивную может осуществиться и с помощью вируса-помощника.
Заражение нечувствительных клеток стандартным вирусом приводит к наиболее распространенной форме абортивной инфекции.
Непермиссивность клетки к определенному вирусному агенту может проявиться на любом этапе инфекции. Чувствительность клетки к ряду вирусов определяется наличием на клеточной поверхности специфических рецепторов, обусловливающих адсорбцию и проникновение вируса в клетку. Такой генетически обусловленный механизм клеточной резистентности наиболее четко установлен для пикорнавирусов, а также онковирусов птиц. Для большинства вирусов можно подобрать две клеточные системы, в одной из которых будет развиваться продуктивная, а в другой — абортивная инфекция. Механизм генетически обусловленной резистентности клеток к вирусам широко варьирует, но в основе его лежит либо отсутствие клеточных факторов, необходимых для репродукции вируса, либо наличие факторов, нарушающих процесс репродукции.
У сложно устроенных вирусов клеточная непермиссивность часто проявляется на стадии сборки вирусных частиц; нарушение сборки в некоторых непермиссивных системах для вирусов гриппа и парамиксовирусов обусловлено уменьшением количества молекул матриксного белка вируса.
Острая и хроническая инфекция.Как продуктивная, так и абортивная инфекция может протекать в виде острой или хронической инфекции.
Острой называется такая форма инфекции, при которой после образования вирусного потомства клетка либо погибает, либо выздоравливает и не содержит вирусных компонентов. Хроническая инфекция — это такая форма инфекции, при которой клетка продолжает продуцировать вирусные частицы или вирусные компоненты в течение длительного времени и передает эту способность дочерним клеткам.
Чаще хроническую форму приобретает абортивная инфекция, так как вирусный генетический материал обычно не входит в состав вирусного потомства, а накапливается в клетках и передается в дочерние клетки. Одним из факторов, вызывающих хроническую инфекцию, являются ДИ-частицы. Такие частицы, попадая в клетки вместе с инфекционными вирусными частицами, конкурируют с ними за факторы репродукции и препятствуют образованию инфекционного потомства. В результате гибель клеток предотвращается. При появлении в системе новых чувствительных клеток в них вновь возникает продуктивная инфекция с образованием ДИ-частиц, и такой цикл инфекции возобновляется снова и снова.
Цитолитическая и нецитолитическая инфекции. Острая инфекция на клеточном уровне может быть цитолитической и нецитолитической в зависимости от судьбы зараженной клетки. Инфекция, завершающаяся гибелью (лизисом) клетки называется цитолитической. Инфекция, которая непосредственно не приводит к лизису клетки, и клетка еще может функционировать в течение некоторого периода времени, продуцируя вирусные частицы, называется нецитолитической.
Смешанная инфекция. Вестественных условиях распространен феномен смешанной инфекции, при котором клетка заражается двумя или несколькими разными вирусами. Два и больше инфекционных процесса, происходящих одновременно в одной клетке, могут оказывать различное влияние друг на друга. Возможны несколько вариантов взаимодействия вирусов в процессе смешанной инфекции.
1. Один из вирусов подавляет репродукцию второго вируса, или подавляется репродукция обоих вирусов. Этот феномен называется интерференцией вирусов.
2. Вирус усиливает репродукцию второго вируса в результате комплементации или экзальтации. Комплементация может происходить между двумя родственными или неродственными вирусами, например между аденовирусом и аденоассоциированным вирусом человека или SV40, при этом вирус-помощник предоставляет другому вирусу неструктурный белок. Экзальтация может быть связана с подавлением процесса образования интерферона первым вирусом.
3. Оба вируса не оказывают существенного влияния на процесс репродукции каждого из них, однако может происходить нарушение морфогенеза обоих вирусов.
Смешанная инфекция широко используется вирусологами для изучения генетических функций вирусов и дефектности геномов.
ЦИТОПАТОЛОГИЯ ЗАРАЖЕННОЙ ВИРУСОМ КЛЕТКИ
Цитопатический эффект является следствием нескольких причин: 1) нарушение нормальной жизнедеятельности клетки в результате механического повреждающего действия вирусных компонентов на клеточные структуры; 2) повреждение лизосом, в результате чего освобождаются высокоактивные лизосомальные ферменты, вызывающие аутолиз клетки; 3) интенсивное истощение белковых и энергетических ресурсов клетки за счет переключения клеточных ферментов и белок-синтезирующего аппарата на синтез вирусспецифических макромолекул; 4) специфическое повреждающее действие вирусов на клеточные молекулы. Эти причины повреждения клетки различным образом проявляются и сочетаются при разных вирусных инфекциях.
Симпласты. Некоторые вирусы вызывают характерный цитопатический эффект, проявляющийся в слиянии клеток и образовании многоядерных клеток, называемых симпластами или синтицием. Образование симпластов обусловлено действием на клеточные мембраны прилежащих друг к другу клеток вирусных белков слияния и определяется тем же механизмом, который обеспечивает слияние вирусной и клеточной мембраны и проникновение вирусов в клетку. Слияние может происходить как за счет белков родительского вируса при заражении клеток большими концентрациями вируса (слияние снаружи), так и за счет внутриклеточного накопления вновь синтезированных вирусных белков слияния (слияние изнутри). Образование симпластов вызывают многие вирусы: парамиксовирусы, некоторые ретровирусы, вирусы герпеса. В определенных условиях (при низких значениях рН) слияние вызывают вирусы гриппа, буньявирусы и др.
Особенности вирусной инфекции в клеточной популяции. Основной особенностью вирусной инфекции в клеточной популяции является гетерогенность системы в связи с гетерогенностью вирусных частиц и клеток, входящих в состав популяции. В любом вирусном препарате наряду с инфекционными вирионами находятся ДИ-частицы. Клетки в каждой клеточной популяции широко варьируют по чувствительности к вирусу, и инфекция может протекать не так, как на клеточном уровне. Например, при заражении вирусом, вызывающим в клетках продуктивную инфекцию, чувствительные клетки популяции могут погибнуть, и в популяции за счет некоторого количества нечувствительных клеток может установиться хроническая инфекция.
Читайте также: